
Jiraporn Nikomtat
Biotechnology, Graduate School,Chiang Mai 50200, Thailand
Narumol Thongwai
Department of Biology, Faculty of Science, Chiang Mai University, Chiang Mai 50200, Thailand
Saisamorn Lumyong
Department of Biology, Faculty of Science, Chiang Mai University, Chiang Mai 50200, Thailand
Yingmanee Tragoolpua
Department of Biology, Faculty of Science, Chiang Mai University, Chiang Mai 50200, Thailand
Research Journal of Microbiology
Year: 2008 | Volume: 3 | Issue: 9 | Page No.: 588-594







ABSTRACT
The effect of Cissus repanda Vahl. plant extract on herpes simplex virus type 1 and type 2 was investigated in this study. The cytotoxicity of dichloromethane and methanol extracts of C. repanda on GMK cells was determined by MTT assay. Non toxic concentrations were used in the study. Methanol extracts of C. repanda showed higher anti-HSV efficacy than dichloromethane extracts, by plaque reduction assay, although, HSV particles were directly inhibited by both extracts. Inhibition of HSV attachment, penetration and replication were also observed, after treatment of HSV with C. repanda extracts. The most affected stage was the attachment period. Moreover, C. repanda extracts inhibited replication of HSV-2 more than HSV-1.
PDF Abstract XML References Citation
How to cite this article
Jiraporn Nikomtat, Narumol Thongwai, Saisamorn Lumyong and Yingmanee Tragoolpua, 2008. Anti-Viral Activity of Cissus repanda Vahl. Plant Extract on Herpes Simplex Virus. Research Journal of Microbiology, 3: 588-594.
URL: https://scialert.net/abstract/?doi=jm.2008.588.594
URL: https://scialert.net/abstract/?doi=jm.2008.588.594
INTRODUCTION
Herpes simplex virus (HSV) is a common human pathogen, widespread in the human population, with two types, herpes simplex virus type 1 (HSV-1) and herpes simplex virus type 2 (HSV-2). HSV-1 infects orofacial mucosal surfaces whilst HSV-2 infects genital mucosal surfaces, resulting in clinical symptoms such as herpes labialis, encephalitis and genital herpes. After primary infection, HSV tends to persist in ganglia (Yang et al., 2005). The virus can be reactivated, causing frequent recurrent infections in some patients, while most people experience few recurrences (Tyring, 1998). Moreover, infections in immunocompromised patients are usually more severe than in immunocompetent hosts (Schuhmacher et al., 2003; Logu et al., 2000). Although, acyclovir (ACV), valaciclovir and famciclovir have been used to treat herpes simplex virus infections, the efficacy of these drugs is limited by side effects and increasing resistance of the viruses at present. Antiviral-resistant HSV strains can occur following mutation in either HSV thymidine kinase (TK) or DNA polymerase. Moreover, several patients with frequent attacks may not be able to afford the cost of long-term treatment, particularly immunocompromised patients (Hammer and Inouye, 1997). Therefore, new antiviral agents from natural products are demanded for prevention and treatment of HSV infection.
Anti-HSV activity has been studied on medicinal plants such as Camellia sinensis Ktze. (Fukuchi et al., 1989; Sydiskia et al., 1991), Nicotiana tabacum Linn. (Larsson et al., 1992), Glycyrrhiza glabra Linn. (Ryu et al., 1993), Rhus javanica Linn. (Kurokawa et al., 1995), Barleria lupulina Lindl., Clinacanthus nutans (Burm.f.) Lindau (Yoosook et al., 1999), Mangifera indica Linn., Maclura cochinchinensis Cornor (Yoosook et al., 2000), Scoparia dulcis Linn. (Galvis et al., 2001), Dunbaria bella Prain (Akanitapichat et al., 2006) and Eugenia caryophyllus (Spreng.) Bullock and S.G. Harrison (Tragoolpua and Jatisatienr, 2007). In this study, we aimed to investigate the anti-HSV activity of Cissus repanda Vahl. (Vitaceae). This medicinal plant can be found in northern Thailand. The plant can be used for treating rashes and detoxication (Chattawanith, 1997).
MATERIALS AND METHODS
Plant Material
Cissus repanda Vahl. (Vitaceae) plants were collected from Queen Sirikit Botanic Garden during July-September, 2005 and were identified by Associate Professor Dr. Chusie Trisonthi and Assistant Professor Paritat Trisonthi, Department of Biology, Faculty of Science, Chiangmai University, Chiangmai, Thailand.
Voucher specimens (QSBG 12006, QSBG 12053) were deposited in the herbarium at Queen Sirikit Botanic Garden, Chiang Mai Province, Thailand.
Cell Line and Viruses
Green monkey kidney cells (GMK) were grown in Minimum Essential Medium (MEM) supplemented with 10% heat-inactivated bovine calf serum (Hyclone), 40 μg mL-1 gentamicin (BIO BASIC INC.), in a humidified 5% CO2 incubator at 37°C.
Herpes simplex virus types 1 (HSV-1; F strain) and 2 (HSV-2; G strain) were used throughout the study. Quantitation of the virus was performed in 24-well tissue culture plates, using a plaque titration assay. Plaques were counted and the virus titers were expressed as plaque forming units (PFU) mL-1.
Plant Extracts
Dried plant leaves and climbers (500 g) were milled and soaked with dichloromethane or methanol at room temperature for 3 days. Then, the suspension was filtered and the solvent was evaporated, using a rotary evaporator and dried with high vacuum. The dry material was dissolved in dimethyl sulfoxide (DMSO) before investigation of anti-HSV activity.
Cytotoxicity Test by MTT Assay
Cytotoxicity tests were performed following previously published procedures (Yu et al., 2004). Cell toxicity was quantified using a MTT (3-[4,5-dimethylthiazol-2-yl]-2, 5 diphenyl tetrazolium bromide assay). GMK cells were seeded on 96-well microplates and incubated with the tested plant extracts for 72 h. Media were removed before being incubated with MTT solution for 4 h. Then, the blue formazan product was eluted from cells by adding DMSO. The absorbance at 570 nm was determined using a microplate reader. The concentration of the 50% cytotoxicity dose (CD50) was calculated.
Inactivation Kinetics
HSVs were treated with non-toxic concentrations of the extracts at room temperature. Aliquots of the virus-extract mixture were taken and diluted for residual virus assay at 1 h intervals for up to 4 h, using the plaque titration assay. The residual virus titers were determined from two independent experiments
Plaque Reduction Assay
The GMK cells were grown in 24-well tissue culture plates as a monolayer. Approximately 100 PFU of HSV were added to the cells per well and were incubated at room temperature for 1 h. Then, 200 μL of media, dichloromethane or methanol extract of C. repanda at non toxic concentrations and ACV at ED50 concentration, were applied into duplicate test wells whereas, media were added into control wells. After that, 400 μL of growth media, containing 2% sodium carboxymethyl cellulose, was added to the cells. After 2-3 days incubation in the CO2 incubator, the cells were stained with 0.1% crystal violet in 1% ethanol. The plaques were counted and inhibitory activities of tested extracts were calculated comparing to control. The 50% effective dose (ED50) was also calculated.
Effect of Plant Extract on Viral Attachment
The GMK cell monolayers were pre-chilled at 4°C for 1 h (Cheng et al., 2002). The media were aspirated and the cell monolayers were infected with HSV in the absence or presence of extracts and acyclovir. Then, the infected cells were further incubation at 4°C for 3 h and the cells were washed with PBS twice before overlaid with growth media, containing 2% sodium carboxymethyl cellulose. After 2-3 days incubation, the cells were stained with 0.1% crystal violet in 1% ethanol. The percentage of inhibition for HSV attachment to GMK monolayer was calculated.
Effect of Plant Extract on Viral Penetration
GMK cell monolayers were pre-chilled at 4°C for 1 h (Cheng et al., 2002). The media were aspirated and cell monolayers were infected by HSV. Infected cells were further incubated at 4°C for 3 h and the extract or acyclovir was added. After further incubation at 37°C for 10 min, the infected cells were treated with PBS, pH 3 for 1 min and PBS, pH 11 were added immediately to neutralize acidic PBS. PBS was then removed and cell monolayer was overlaid with growth media, containing 2% sodium carboxymethyl cellulose and incubation for 2-3 days. The cells were stained with 0.1% crystal violet in 1% ethanol and the percentage of inhibition for HSV penetration was calculated.
Effect of Plant Extract on HSV Replication
GMK cells were grown as monolayers in 25 cm2 flasks. The cells were infected with 1x106 PFU mL-1 of HSV. Virus was allowed to adsorb to the cells for 60 min at room temperature. Then, unadsorbed virus was discarded and the cells were washed twice with PBS. The duplicated flasks of infected cells were maintained in media containing crude extract of C. repanda at non-toxic concentration while only media or media containing acyclovir at ED50 concentration were added into control flasks. Infected cells were observed for cytopathic effects (CPE) and were collected at 6, 12, 24 and 30 h after viral infection. The cells were frozen and thawed twice. The supernatants containing viruses were kept at -80°C and virus titers were determined using plaque titration assay.
RESULTS
The CD50 values of C. repanda, extracted with dichloromethane (CD) and methanol (CM) were 737±26 and 1876±0 μg mL-1, respectively. Thus, non-toxic concentrations of C. repanda extract were used to determine anti-HSV activity.
After incubation of viruses with the dichloromethane or methanol extract of C. repanda at room temperature for 1 h intervals up to 4 h, both HSV-1 and HSV-2 were directly inactivated (Table 1). After incubation of HSV-1 and HSV-2 with CD extract for 2 h, the viruses were directly inactivated to negligible amounts. Similar results were observed when HSV-2 was incubated with CM extract, whereas HSV-1 was inactivated after incubation with CM extract for only 1 h.
C. repanda extracts demonstrated anti-HSV activity against both types of HSVs using plaque reduction assay. HSV-1 was inhibited by CM and CD extracts with ED50 values of 310.6±50.0 and 249.2±40.7 μg mL-1 and selective index (SI) values of 6±1.2 and 2.9±0.4, respectively (Table 2). HSV-2 was also inhibited by CM and CD extracts with ED50 values of 295.4±56.4 and 253.4±15.9 μg mL-1 and SI values of 6.3±1.6 and 2.9±0.2, respectively. Therefore, the SI of CM extract on HSV was higher than CD extract. Moreover, ED50 values of ACV on HSV-1 and HSV-2 were 2.31 and 14.92 μg mL-1, respectively.
| Table 1: | Direct inactivation of HSV-1 and HSV-2 by methanol (CM) and dichloromethane (CD) extracts of C. repanda |
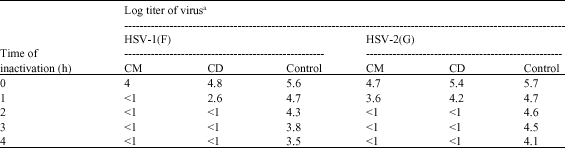 | |
| aValues are means of two independent experiments | |
| Table 2: | Anti-HSV activity of methanol (CM) and dichloromethane (CD) extracts of C. repanda on GMK cells |
 | |
| a,bValues are means±SD of two independent experiments cCD50 ED50-1 | |
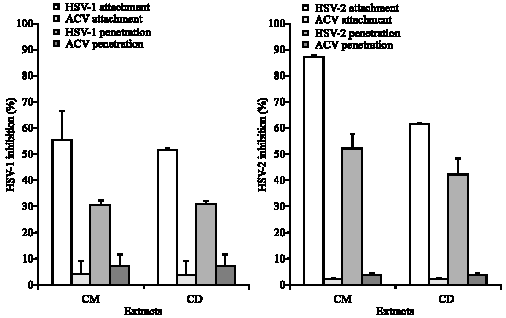 | |
| Fig. 1: | Effects of extracts and acyclovir on HSV attachment and penetration to GMK cells |
The effects of C. repanda extracts on HSV attachment are shown in (Fig. 1). CM and CD extracts blocked HSV-1 attachment with percentage of 55.3±11.1 and 51.5±0.8, respectively. Similar results were observed when CM extract was applied to determine the blocking of HSV-2 attachment. The efficacy of CM extract was better than CD extract with percentages of 87.5±0.7 and 61.5±0.4, respectively.
Furthermore, CM and CD extracts inhibited HSV-1 penetration with percentages of 30.6±1 and 31.0±0.8, respectively, while HSV-2 penetration was blocked by CM and CD extracts with percentages of 51.8±5.9 and 42.1±6.4, respectively (Fig. 1).
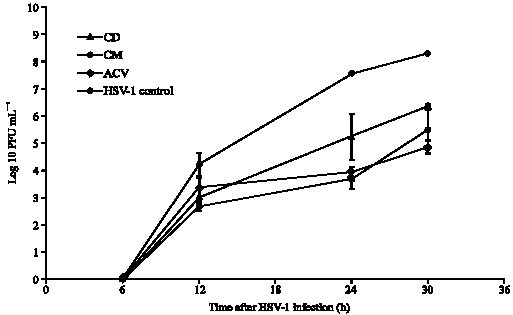 | |
| Fig. 2: | Inhibition of HSV-1 replication by dichloromethane (CD) and methanol (CM) of C. repanda extract compared with ACV and control |
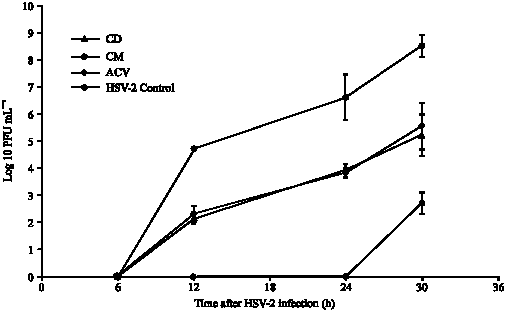 | |
| Fig. 3: | Inhibition of HSV-2 replication by dichloromethane (CD) and methanol (CM) of C. repanda extracts compared with ACV and control |
To determine the effects of C. repanda extracts on HSV yield, infected cells were maintained in the extract and were collected at 6, 12, 24 and 30 h. At 30 h after HSV-1 infection, log10 PFU mL-1 of virus control and ACV treated virus were 8.3±0.01 and 4.8±0.2, respectively. An inhibition of HSV-1 replication was observed, since the log10 PFU mL-1 of virus after treatment with CM and CD extracts were 5.5±0.8 and 6.3±0.1, respectively (Fig. 2). HSV-2 replication was also inhibited after treatment with the extracts. At 30 h after infection the log10 PFU mL-1 of HSV-2 were 2.7±0.4 and 5.2±0.8 after treatment with CM and CD extracts compared with 8.5±0.4 of control virus and 5.6±0.9 of ACV treated virus (Fig. 3). Therefore, the inhibitory activity of CM extract on HSV replication was higher than CD extract.
DISCUSSION
In this study, we investigated anti-HSV activity of C. repanda, which is a medicinal plant species found in northern Thailand, used in traditional remedies (Chattawanith, 1997). The toxicity on GMK cells of dichloromethane extract was higher than methanol extract of C. repanda. Anti-HSV activity was observed after treatment of HSV with non-toxic concentrations of the extracts using the plaque reduction assay. ED50 values of the CD extract were slightly lower than those obtained with the CM extract, when HSV-1 and HSV-2 were tested with the extracts. However, SI values (CD50 ED50-1) of CM extracts were about 2-fold higher than treatment with CD extracts. Thus, CM extracts showed higher anti-HSV efficacy than CD extracts. Moreover, the effects of the extracts on various stages of HSV multiplication cycles were determined to elucidate modes of action of the extracts. Both CM and CD extracts had an ability to inhibit HSV particles. HSV-1 was directly inactivated by CM and CD extracts to negligible amounts within 1 and 2 h, respectively, whereas the inactivation of HSV-2 particles was observed after incubation with CM and CD extracts for 2 h. Therefore, the efficiency of the CM extracts on inactivation of HSV-1 particle was higher than that of the CD extracts. Inhibition of HSV attachment and penetration was also observed. CM and CD extracts inhibited HSV-1 attachment 1.8 and 1.7 fold higher than penetration respectively. Similar results were obtained after treatment of HSV-2 with the extracts, with inhibition of HSV-2 attachment being 1.7 and 1.5 fold higher than viral penetration, after treatment with CM and CD extracts respectively.
Thirty hours after HSV-1 replication, the amounts of the virus were reduced by 33.7 and 24.1%, compared with the control after treatment with CM and CD extracts, respectively. Higher percentage inhibition of viral replication was observed on HSV-2 after treatment with CM and CD extracts with the reduction of viral amounts of 68.2 and 38.8%, respectively.
These results indicate that dichloromethane and methanol extracts of C. repanda inhibit HSV at various stages in the viral multiplication cycle i.e. attachment, penetration and replication. The most affected stage was the attachment period. The highest anti-HSV activity was obtained with methanolic extracts from C. repanda. Moreover, C. repanda extracts inhibited replication of HSV-2 more than HSV-1. This is the first report of anti-HSV activity of C. repanda, besides the activity on HSV-1 by crude extract of endophytic fungi isolated from C. repanda (Wiyakrutta et al., 2004). However, biologically active compounds from C. repanda and their modes of action should be further studied.
ACKNOWLEDGMENTS
This research was granted by the Commission on Higher Education granting Ms. Jiraporn Nikomtat was supported forStrategic Scholarships Fellowships Frontier Research Networks from the Commission on Higher Education. We also thank Uttaradit Rajabhat University, Graduate school Chiang Mai University and The Royal Project Grant number 3015-3396 for supporting this study and Dr. Stephen Elliott for critically reading the manuscript.
REFERENCES
- Akanitapichat, P., A. Wangmaneerat, P. Wilairat and K.F. Bastow, 2006. Anti-herpes virus activity of Dunbaria bella Prain. J. Ethnopharmacol., 105: 64-68.
CrossRefDirect Link - Cheng, H.Y., C.C. Lin and T.C. Lin, 2002. Antiherpes simplex virus type 2 activity of casuarinin from the bark of Terminalia arjuna Linn. Antivir. Res., 55: 447-455.
CrossRefPubMedDirect Link - Fukuchi, K., H. Sakagami, T. Okuda, T. Hatano and S. Tanuma et al., 1989. Inhibition of herpes simplex virus infection by tannins and related compounds. Antivir. Res., 11: 285-297.
PubMedDirect Link - Galvis, L.B., C. Zuluaga, M. Arno, M.A. González and R.J. Zaragozá, 2001. Structure-Activity relationship of in vitro antiviral and cytotoxic activity of semisynthetic analogues of Scopadulane diterpenes. J. Nat. Prod., 64: 1318-1321.
Direct Link - Kurokawa, M., K. Nagasaka, T. Hirabayashi, S. Uyama and H. Sato et al., 1995. Efficacy of traditional herbal medicines in combination with acyclovir against herpes simplex virus type 1 infection in vitro and in vivo. Antiviral Res., 27: 19-37.
PubMedDirect Link - Larsson, P.A., J.M. Hirsch, J.S. Gronovitz and A. Vahlne, 1992. Inhibition of herpes simplex virus replication and protein synthesis by non-smoked tobacco, tobacco alkaloids and nitrosamine. Arch. Oral. Biol., 37: 969-978.
PubMedDirect Link - De Logu, A., G. Loy, M.L. Pellerano, L. Bonsignore and M.L. Schivo, 2000. Inactivation of HSV-1 and HSV-2 and prevention of cell-to-cell virus spread by Santolina insularis essential oil. Antivir. Res., 48: 177-185.
PubMedDirect Link - Ryu, S.Y., C.K. Lee, J.W. Ahu, S.H. Lee and O.P. Zee, 1993. Antiviral activity of triterpenoid derivatives. Arch. Pharml. Res., 16: 339-342.
Direct Link - Schuhmacher, A., J. Reichling and P. Schnitzler, 2003. Virucidal effect of peppermint oil on the enveloped viruses herpes simplex virus type 1 and type 2 in vitro. Phytomedicine, 10: 504-510.
CrossRefDirect Link - Sydiskis, R.J., D.G. Owen, J.L. Lohr, K.H. Rosler and R.N. Blomster, 1991. Inactivation of enveloped viruses by anthraquinones extracted from plants. Antimicrob. Agents Chemother., 35: 2463-2466.
PubMedDirect Link - Tragoolpua, Y. and A. Jatisatienr, 2007. Anti-herpes simplex virus activities of Eugenia caryophyllus (Spreng.) Bullock and S.G.Harrison and essential oil, eugenol. Phytother. Res., 21: 1153-1158.
Direct Link - Tyring, S.K., 1998. Advances in the treatment of herpesvirus infection: The role of famciclovir. Clin. Ther., 20: 661-670.
CrossRefDirect Link - Wiyakrutta, S., N. Sriubolmas, W. Panphut, N. Thongon, K. Danwisetkanjana, N. Ruangrungsi and V. Meevootisom, 2004. Endophytic fungi with anti-microbial, anti-cancer and anti-malarial activities isolated from Thai medicinal plants. World J. Microbiol. Biotechnol., 20: 265-272.
CrossRefDirect Link - Yang, C.M., H.Y. Cheng, T.C. Lin, L.C. Chiang and C.C. Lin, 2005. Acetone, ethanol and methanol extracts of Phyllanthus urinaria inhibit HSV-2 infection in vitro. Antivir. Res., 67: 24-30.
CrossRefDirect Link - Yoosook, C., Y. Panpisutchai, S. Chaichana, T. Santisuk and V. Reutrakul, 1999. Evaluation of anti-HSV-2 activities of Barleria lupulina and Clinacanthus nutans. J. Ethnopharmacol., 67: 179-187.
CrossRefDirect Link - Yoosook, C., N. Bunyapraphatsara, Y. Boonyakiat and C. Kuntasuk, 2000. Anti-herpes simplex virus activities of crude water extracts of Thai medicinal plants. Phytomedicine, 6: 411-419.
PubMedDirect Link - Yu, Z., W. Li and F. Liu, 2004. Inhibition of proliferation and induction of apoptosis by genistein in colon cancer HT-29 cells. Cancer Lett., 215: 159-166.
CrossRefDirect Link