
Sahar El-Shatoury
Department of Botany, Faculty of Sciences, Suez Canal University, Ismailia, Egypt
Hesham Abdulla
Department of Botany, Faculty of Sciences, Suez Canal University, Ismailia, Egypt
Ahmed Dewedar
Department of Botany, Faculty of Sciences, Suez Canal University, Ismailia, Egypt
Research Journal of Microbiology
Year: 2007 | Volume: 2 | Issue: 1 | Page No.: 74-80







ABSTRACT
A Box-Wilson central composite design was applied to study the effect of Micromonospora chalcea inoculum-size, municipal sludge and rice straw ratios on decomposition of the straw in sandy soil. The experiment included 20 runs with five levels for each of the three factors. Results indicated highly significant effect of sludge (p = 0.005) on the straw decomposition and organic carbon release. Although inoculum size of M. chalcea was not significantly effective, it showed a significant interaction with sludge (p = 0.021). Probably, indicating a critical importance of sludge as source of essential nutrients for supporting M. chalcea growth. Extracellular enzyme profiling of M. chalcea revealed a general wide activity including: polysaccharide hydrolases, proteases, lipases, phosphatases and amino-peptidases. Response surface methodology was employed for the optimization of rice straw decomposition by M. chalcea. Conditions that supported highest carbon release were incorporating 100 g straw with 40 g sludge into 100 g sandy soil, leading to 10 folds reduction in time of straw decomposition compared to that before optimization. On the other hand, sludge ratios above 81% have retarded the decomposition process even under straw ratio as low as 10%.
PDF Abstract XML References
How to cite this article
Sahar El-Shatoury, Hesham Abdulla and Ahmed Dewedar, 2007. Factorial Design for Optimization of Rice Straw Incorporation into Soil Using Micromonospora chalcea. Research Journal of Microbiology, 2: 74-80.
URL: https://scialert.net/abstract/?doi=jm.2007.74.80
URL: https://scialert.net/abstract/?doi=jm.2007.74.80
INTRODUCTION
The microbial decomposition of lignocellulosic agricultural wastes is recently recognized as a major renewable resource, with potential for conversion to a variety of products such as soil biofertilizers, industrial enzymes, animal feeds and biofuels (Howard et al., 2003). However, cellulose crystallinity and lignin - which offers cellulose physical protection against cellulolytic enzymes-limit the decomposition of polysaccharides present in lignocellulose residues for biotechnological application (Lynd et al., 2002). Rice straw is one example of the lignocellulosic wastes that is often disposed by biomass burning; a practice which is not restricted to developing countries alone, but is considered a global phenomenon (Levine, 1996). Rice straw composed mainly of cellulose and hemicellulose encrusted by lignin, in addition to a small amount of protein which makes it high in C:N ratio and persistent to microbial decomposition compared to straw from other protein-rich grains such as wheat and barely (Parr et al., 1992).
Many actinomycetes can degrade cellulose and solubilize the lignin structure as their primary metabolic activity at high nitrogen levels compared to white rot fungi (Eriksson et al., 1990). Previous studies shown that actinomycete strains, particularly Streptomyces, Micromonospora and Nocardioides are capable of attacking the lignocellulosic components of rice straw, causing significant release of carbon and to degrade synthetic organic dyes that are structurally related to lignin (El-Shatoury et al., 2005a, b; Abdulla and El-Shatoury, 2006). The addition of sludge, as a rich source of nitrogen and labile carbon, enhanced the decomposition of straw by the above mentioned actinomycete strains and was suggested to accelerate its incorporation into soil.
Traditional methods of optimizing straw decomposition have involved changing one variable, while fixing the others at a certain level. This single dimensional approach is time consuming and fails to locate optimal conditions because it doesn't consider the effect of possible interactions between variables. Factorial design and response surface analysis, first described by Box and Wilson (1951), is an experimental strategy for seeking the optimum conditions for a multivariable system. This technique has been widely used in applied sciences such as optimizing enzyme production and assessment of drug interactions (Kalil et al., 2000); however, it has not been well exploited to optimize decomposition of agriculture wastes by microorganisms. Although few previous studies have focused on response surface optimization of lignocellulosic enzymes in fermentation media (Cacchio et al., 2001; Trupkin et al., 2003), this is the first report-to the best of our knowledge-regarding optimization of agriculture wastes incorporation into soil.
The goal of this study is to study the effect of various ratios of sludge, rice straw and M. chalcea inoculum on accelerating the straw decomposition in soil and to investigate the extracellular enzyme profile of M. chalcea that may contribute to accelerating the decomposition process. Response surface optimization technique was applied to determine the optimal conditions for accelerating straw incorporation into soil.
MATERIALS AND METHODS
Microorganism and Inoculum Preparation
Micromonospora chalcea strain isolated by Abdulla and El-Shatoury (2006) was used in the experiment. The inoculum was produced from spore suspension preserved at -20°C, inoculated into yeast extract dextrose broth and incubated at 28°C, 100 rpm for 5 days. Mycelia were harvested by centrifugation, homogenized, washed twice in phosphate buffer and used for straw incorporation and enzyme profiling experiments.
Straw Incorporation Factorial Design
A Box-Wilson Central Composite Design (CCD) with five levels for each of three factors (straw, sludge and inoculum size) was studied which required 20 experimental runs, including 6 central points (Table 1). Central points provide additional degree of freedom for error estimation, which increase power when testing the significance of effects (Carvalho et al., 1997). All runs were performed in duplicates using 17x12x7 cm plastic pots containing 100 g sandy soil.
| Table 1: | Values for the 20 runs in the CCD |
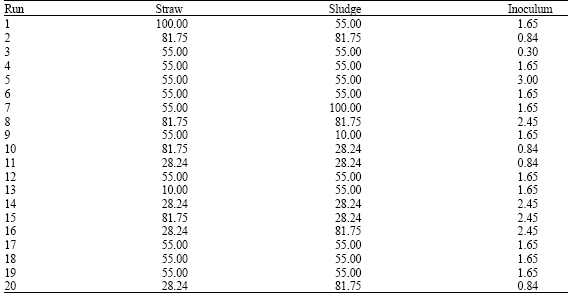 | |
| Table 2: | Activity of extracellular enzymes from Micromonpspora chalcea measured on solid media |
 | |
| Table 3: | Activity of extracellular enzymes from Micromonpspora chalcea measured using API-ZYM® system |
 | |
| a Zero corresponds to a negative reaction, 5 to a reaction of maximum color intensity. For the purpose of this study, values were reported as low activity (1), moderate activity (2-3) and high activity (4-5) | |
Thermally treated municipal sludge (to eliminate pathogens), coarsely chopped rice straw and M. chalcea harvested mycelia were mixed thoroughly with soil in the pots according to the corresponding levels for each run and incubated under laboratory conditions. Moisture content was adjusted gravimetrically at 40-50% by water spraying and mixing. Organic carbon was detected by wet oxidation using modified Walkely-Black method (Alf and Nannipieri, 1995). Samples were analyzed at intervals and statistical analysis was performed using multiple regressions and ANOVA using Minitab v12 statistical computing package.
Enzyme Profiling
Extracellular enzyme profiling was determined on solid media and using the API-ZYM® system (bioMérieux). Extracellular enzymes detection on solid media (Table 2) was performed according to the methods of (Wollum II, 1982; Williams et al., 1983). Enzyme screening plates were inoculated with 5 ΜL of M. chalcea mycelia suspension and scored for enzymatic activities after incubation at 28°C for 7-14 days.
The API-ZYM® system (bioMérieux) is a semi-quantitative method allowing examination of 19 hydrolysates (Table 2) in a strip with a series of microcupules containing dehydrated chromogenic substrates of 19 different enzymes against a control (substrate free) microcupule. A 65 μL of M. chalcea mycelia suspension were dispensed into each of the 20 microcupules of the API-ZYM®, incubated and color reactions were performed according to the producer instructions. Readings were scored using the API-ZYM® color chart ranging from 0 (negative) to 5 (maximum) (Table 3).
RESULTS AND DISCUSSION
Results for change in carbon as a function of time in the CCD are shown in Fig. 1. The statistical analysis were performed with data obtained at 10 days of incorporation as there was no significant increase in the available organic carbon for most of the runs (p>0.05) after this time. Also, organic carbon has significantly decreased after 25 days (p = 0.009), indicating the start of mineralization of carbon, particularly in runs number 2, 8 and 20.
The effect estimate for each variable (straw, sludge and inoculum) and their interactions at 10 days of experiment were determined and reported in Table 4. The decomposition process performance was measured by the release of organic carbon response. Both t-test and p-value confirmed the highly significant effect of sludge on the straw decomposition and organic carbon release. Similar conclusion was reported by Strauss et al. (2003); since sludge is relatively rich in nitrogen and phosphorus, its co-incorporation with straw is advantageous and complements the high carbon content in rice straw. Although inoculum size was not effective in this factorial design (p = 0.066), it exhibited a significant interaction with sludge (p = 0.021). This result indicates a critical importance of sludge as source of essential nutrients for supporting M. chalcea growth. In addition, M. chalcea is probably well adapted to metabolize organic matter in sludge and maintain good mycelia growth; therefore variation in inoculum size was not significantly effective. A previous pilot study (Hayashida et al., 1988) descried complete composting of one ton of poultry feces by Streptomyces, Thermoactinomyces and Thermomonospora strains after 10 days incubation over a wide temperature range (15-60°C) in 30 cm depth containers. That pilot study showed satisfactory growth and efficient penetration of mycelia into feces in 30 h under optimal conditions. In order to assess the metabolic ability of M. chalcea, its extracellular enzymatic profiling was investigated (Table 2 and 3). The strain showed wide extracellular enzymatic abilities, including polysaccharide hydrolases, esterases, amino-peptidases, proteases, lipases, phosphatases, pectinase and chitinase. Tiquia (2002) have reported a particular abundance of polysaccharide hydrolases, esterases and amino-peptidases during manure composting.
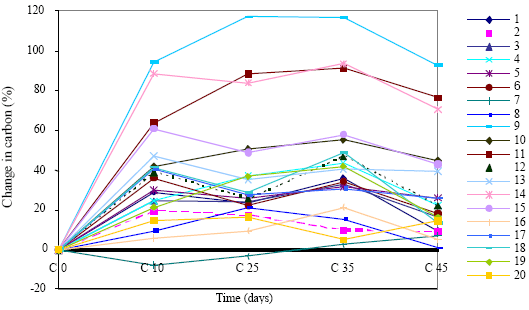 | |
| Fig. 1: | Percentage of change in organic carbon as a function of time for the 20 runs in the central composite experimental design |
| Table 4: | Main effects and interactions analysis from the CCD for carbon after 10 days of incorporation |
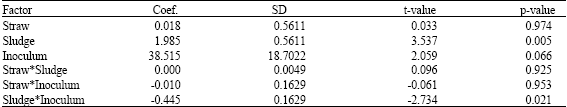 | |
| Values in bold indicates significance at p<0.05 | |
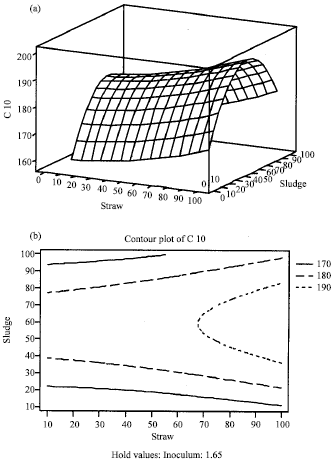 | |
| Fig. 2: | Response surface (a) and contour plot (b) for the release of carbon as a function of straw and sludge percentage, with 1.65% Micromonospora inoculum, after 10 days of incorporation into sand soil |
The enzymatic ability of M. chalcea is, thus, well suited for successful growth in straw-sludge environment, attacking the lignocellulosic material of straw and may explain the significant organic carbon release after 10 days of inoculation. The resulted partially decomposed lignocellulose represents a good source of labile carbon for other indigenous microflora in the experiment to sustain the decomposition process.
Estimation of the effect for the variables (straw, sludge and inoculum) and their interactions at 10 days of experiment using nitrogen content response was also determined. Regression analysis indicated no significant effect of any of the three variables on nitrogen content (p>0.05). Moreover, no significant interactions between variable was obtained (data not shown). Therefore, nitrogen content was not considered an adequate response for measuring the straw decomposition process in this study.
| Table 5: | Analysis of variance for the CCD at 10 days of incorporation |
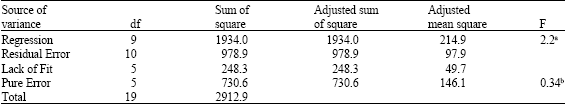 | |
| Regression coefficient: R = 0.96. a calculated F-value (regression/residual), b critical F-value (lack of fit/pure error) | |
A second order model equation was established, based on ANOVA (Table 5), to describe the increase of carbon as function of straw and sludge ratio in soil, for 10 days of incorporation experiment. The p-value for lack-of-fit was 0.911 indicating the adequacy of this full quadratic model to fit the data. Based on the F-test, the model is predictive since its calculated F-value is greater than critical F-value and the regression coefficient (0.96) is close to unity, indicating that 96% of the variations in the response are explained by this model, as it is well established that values above 0.9 are considered very good (Haaland, 1989).
The full quadratic model was used to generate response surfaces for the analysis of the variable effects on organic carbon release. As seen in Fig. 2, an increase in both straw and sludge ratio can lead to accelerated straw decomposition with subsequent increase in organic carbon release. However, sludge ratios above 81 g/100 g soil have retarded the release of organic carbon, even under straw ratio as low as 10% in soil. This result may indicate an inhibitory effect of high nitrogen concentrations in sludge on the decomposition process. Previous studies have shown that nitrogen may actually inhibit the decomposition of the lignin fraction of litter either by inhibiting synthesis of lignocellulolytic enzymes or by reacting with breakdown products to form other compounds that resist decay (Hobbie and Vitousek, 2000; Fog, 1988).
In conclusion, the extracellular enzyme profiling of M. chalcea revealed presence of various enzymatic groups that are involved in lignocellulose decomposition. On the basis of response surface analysis, the optimum conditions to accelerate rice straw decomposition in soil were incorporating 100% straw with 40% sludge and 1.65% inoculum of M. chalcea. At these conditions, a value of 190 C mg g-1 was obtained after 10 days. This represents a reduction of 10 folds in the time required for straw decomposition, as obtained by Shiga et al. (1985). Therefore, with the effective decomposition of rice straw and simultaneous reduction in time, its incorporation into soil using M. chalcea can be regarded as possible and economically attractive technique for safe disposal of rice straw.
ACKNOWLEDGMENTS
We are grateful to Fatma Hagag, Suez Canal University Center for Environmental Studies and Consultation, for her technical assistance.
REFERENCES
- Abdulla, H.M. and S. El-Shatoury, 2007. Actinomycetes in rice straw decomposition. Waste Manage., 27: 850-853.
Direct Link - Box, G.E.P. and K.B. Wilson, 1951. On the experimental attainment of optimum conditions. J. Roy Stat. Soc., 13: 1-45.
Direct Link - Carvalho, C.M.L., M.L.M. Serralheiro, J.M.S. Cabral and M.R. Aires-Barros, 1997. Application of factorial design to the study of transesterification reactions using cutinase in AOT-reversed micelles. Enzyme. Microbiol. Technol., 21: 117-123.
Direct Link - Hobbie, S.E. and P.M. Vitousek, 2000. Nutrient limitation of decomposition in Hawaiian forests. Ecology, 81: 1867-1877.
Direct Link - Howard, R.L., E. Abotsi, E.L.J. van Rensburg and S. Howard, 2003. Lignocellulose biotechnology: Issues of bioconversion and enzyme production. Afr. J. Biotechnol., 2: 602-619.
Direct Link - Lynd, L.R., P.J. Weimer, W.H. van Zyl and I.S. Pretorius, 2002. Microbial cellulose utilization: Fundamentals and biotechnology. Microbiol. Mol. Biol. Rev., 66: 506-577.
CrossRefPubMedDirect Link - Parr, J.F., R.I. Papendick, S.B. Hornick and R.E. Meyer, 1992. Soil quality: Attributes and relationship to alternative and sustainable agriculture. Am. J. Alter. Agric., 7: 5-11.
CrossRef - Strauss, M., S. Drescher, C. Zurbrugg, A. Montangero, O. Cofie and P. Drechsel, 2003. Co-composting of fecal sludge and municipal organic waste: A literature and state of knowledge-review. International Water Management Institute (IWMI) Accra, Ghana, Swiss Federal Institute of Environmental Science and Technology (EAWAG), Department of Water and Sanitation in Developing Countries (SANDEC) CH-Duebendorf, Switzerland.
- Tiquia, S.M., 2002. Evolution of extracellular enzyme activities during manure composting. J. Appl. Microbiol., 92: 764-775.
CrossRefDirect Link - Trupkin, S., L. Levin, A. Forchiassin and A. Viale, 2003. Optimization of a culture medium for ligninolytic enzyme production and synthetic dye decolorization using response surface methodology. J. Ind. Microbiol. Biotechnol., 30: 682-690.
CrossRefDirect Link - Williams, S.T., M. Goodfellow, G. Alderson, E.M.H. Wellington, P.H.A. Sneath and M.J. Sackin, 1983. Numerical classification of Streptomyces and related genera. J. Gen. Microbiol., 129: 1743-1813.
CrossRefPubMedDirect Link