
Ann S. O`Neil
Department of Biology, University of Kentucky, Lexington, 40506-0225 Kentucky, United States of America
Changsoo Kim
School of Biological Sciences and Technology, Chonnam National University, 61186 Gwangju, Republic of Korea
Robin L. Cooper
Department of Biology, University of Kentucky, Lexington, 40506-0225 Kentucky, United States of America
LiveDNA: 1.523
Journal of Entomology
Year: 2020 | Volume: 17 | Issue: 2 | Page No.: 36-47







ABSTRACT
Background and Objective: A chemical stimulus might cause a mild repulsive reaction which can be used to train an animal to avoid a cue. Repeated training may be able to induce associative learning to an unconditioned stimulus (US). The retention (memory) of the US would likely be stronger for more sensitive animals. Materials and Methods: A genetically engineered strain of Drosophila which contains inserted genes to code for the TRPV receptor (i.e., the capsaicin or heat receptor) were used to examine this topic. Results: The background control (UAS-TRPV1 strain) did show a preference to light or dark. However, associative learning to fructose (FRU) or quinine hemisulfate (QUI) was only demonstrated in 2nd instars. The transgenic larvae are extremely sensitive to capsaicin while background strain show no aversion to the capsaicin. There is preference for the dark in both background and TRPV1 expressing larvae. The background and TRPV1 receptor expressing larvae did not show associative learning to a single exposure of capsaicin. Conclusion: Even though there is a strong response to a noxious stimulus, this learned behavior is not retained when associated with non-noxious stimulus.
PDF Abstract XML References Citation
Copyright: © 2020. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Ann S. O`Neil, Changsoo Kim and Robin L. Cooper, 2020. Learning and Memory Retention in Larval Drosophila. Journal of Entomology, 17: 36-47.
DOI: 10.3923/je.2020.36.47
URL: https://scialert.net/abstract/?doi=je.2020.36.47
DOI: 10.3923/je.2020.36.47
URL: https://scialert.net/abstract/?doi=je.2020.36.47
INTRODUCTION
Associative learning became known from experiments in 1902 by the physiologist Ivan Pavlov. Many studies have now been conducted in associative learning in various types of animals. A genetically amenable model animal used to understand the basic physiological mechanism in neurobiology is the fruit fly, Drosophila melanogaster. Flies and mammals share many similar genes1, suggesting that underlying cellular mechanisms are similar. There are various studies on adult Drosophila with associative learning2; however, less focus has been targeted to developing instar stages3-6. Larvae are useful to focus on due to the simplicity of the animal with having 21 versus 1300 receptors for olfactory and 80 versus 650 for taste as compared to mammals7,8. The Drosophila larvae show the ability to learn through olfactory conditioning9. There are multiple ways to condition larvae for associative learning (i.e. olfactory, gustatory). In the studies herein, a light stimulus with different food sources as positive and negative reinforcements was used. Classical conditioning with positive (fructose) and negative (quinine hemisulfate or capsaicin) reinforcements were paired with either light or dark.
One trail learning paradigms are known to occur in humans and in some vertebrate animals10,11. Recently one trial learning in Drosophila has been addressed12. One would assume that very intense pain inducing stimuli, such as intense heat to the skin for mammals, would be an evolutionary engrained response for many animals, even possibly for insects. The density and responsiveness of the sensory neurons determines how well the stimuli are received. Some pathological conditions occur in humans where an individual is super sensitive to a mild stimulus13. The rationale in this study was to examine if such conditions are present in invertebrates with enhanced sensory perception.
The questions examined in this study are: (1) Will the larval stages (1st, 2nd and 3rd) show equal abilities to perform associative learning within a given experimental paradigm? (2) Will enhanced sensory perception to negative reinforcement allow induced 1 trail associative learning? (3) What is the retention time of learning in Drosophila larvae that have an enhanced sensory perception to negative reinforcement?
The hypothesis being tested is that 1st and 2nd instars will have a greater ability to demonstrate associative learning than the early 3rd instars. The rationale for this hypothesis is that 3rd instars have a transitional period where early 3rd instars switch preferences from dark to light environment in becoming late 3rd instars. A second hypothesis is that enhanced negative stimulation to larvae that are more sensitive to a stimulus will learn quicker and, with a strong stimulation, will demonstrate a 1 trial learned response. For the last point above, another hypothesis being tested is that a strong negative reinforcement in the larvae expressing an over production of TRPV1 (vanilloid receptor or capsaicin receptor) will have a longer memory retention than background strains throughout larval development.
MATERIALS AND METHODS
Positive and negative reinforcements
Fructose and quinine sulfate: Learning in larvae was examined in the manner of light/dark association to fructose (FRU) or quinine hemisulfate (QUI). The association of FRU and QUI was reversed with dark or light to insure learning was associative. Evidence for learning was then assessed by preference to various areas within the Petri dish.
Associative learning was conducted within Petri dish assay plates (90mm in diameter). Control plates consisting of the 1% agarose solution and were used for the observations of pre- and post-conditioning. For the learning regiment assay a 1% agarose with 1 M fructose for positive reinforcement and 1% agarose and 0.2% quinine hemisulfate for negative reinforcement. The concentrations were determined from the study by Hendel et al.14. In order to have a baseline of preference between light and dark, the larvae were first observed in pre-conditioning to eliminate initial desire toward light or dark areas on the dish.
Experimentation was conducted at the temperature the animals were raised to eliminate temperature changes. All light in the room was turned off except for the light box upon which each of the dishes were placed on. Each animal was alternated in two minute intervals back and forth between the FRU in the light environment and QUI in the dark environment agar plates. These alternating environments were conducted for 5 trials in each for 2 min. The protocol for recording post conditioning data is the same as with preconditioning. New larvae were used to test with FRU in dark and QUI in light (Fig. 1a-b).
The novel design of strips instead of pie shaped quadrants, as used in past studies15, was used in order to make the light vs. dark preference more readily available to the larvae.
 | |
| Fig. 1(a-b): | (a) Plain agar plate used for pre and post conditioning and (b) Conditioning plate placed in light and the black felt lined box covers the plate during the conditioning in the dark |
The width of the strips is 10 mm for 3rd instars (twice the body length of 3rd instar larvae) and the widths are twice the lengths for the 2nd instars and 1st instars (Fig. 2). This method allows for the larvae to move readily into different environments.
Pure capsaicin: To test for one trial associative learning, different concentrations of capsaicin were used (1 nM and 1 μM). Capsaicin was mixed in the agar prior to making the plates. In this method the same set up as the FRU and QUI learning was used, but the pure capsaicin took the place of the QUI in the agar plate. To test which concentration would be best to use for future studies, associative learning in the 3rd instars in both background strain (UAS-TRPV1) strain and over expression TRPV1 larvae were examined. It was determined a high concentration (1 μM) was to be used as the negative reinforcement for the retention studies.
Drosophila: To address if inducing one trial established associative learning and retention the strain with over expression of the TRPV1 receptor was used. The expression was targeted specifically for multi dendritic sensory neurons (md-Gal4>UAS TRPV1). The background UAS-TRPV1 was used as a control.
Data analysis
Preference values: For each experiment, larvae were tested individually. On the pre- and post-conditioning agar plates the larvae were scored on their preference of light or dark every 30 sec for 2 min. The method described by Gerber et al.16 to calculate the preference value was used. Each individual larva was calculated as follows:
PREF values are between 1 and -1, where a positive value indicates preference for and a negative value for avoidance of light. For each group of larvae trained on Light (+)/dark (-), a second group was reciprocated with Light (-)/dark (+). To test for significance, the data is paired with the reciprocated experimental data and a Wilcoxon signed rank test was used. The comparison of reciprocally trained animals and hence the conclusion regarding associative learning is unaffected by baseline preferences for dark or light16.
Learning index: To measure learning performance between experimental conditions the approach by Gerber et al.16 was followed. The learning index (LI) for paired individual larvae was as follows:
where, LI values are between 1 and -1. A positive value indicates conditioned learning towards light and a negative value indicates conditioned avoidance of light. For statistical testing of comparing the LI against zero, a one-sample sign test is used; for two group comparisons of LIs the Mann-Whitney U test was used. This significance testing was used when comparing the background strain to the overexpression TRPV1 strain (p<0.05 is considered significant).
 | |
| Fig. 2(a-c): | Banded patterns are printed on plastic transparencies that are used on the light box for the pre and post conditioning, (a) Banded pattern for 3rd, (b) 2nd and (c) 1st instars |
| Double headed arrow is 1 cm scale | |
Testing retention: To test retention, the larvae in the over expression TRPV1 strain were used. Capsaicin was assumed to be the negative reinforcement. Three experiments were conducted to test retention. The first test was to address retention from 1st to 2nd instars after a one time exposure. The second experiment was to test retention from 1st to 3rd instars after a one time exposure. The last experiment was to test retention from 2nd to 3rd instars after a one time exposure.
This was performed by conditioning the larvae during either the 1st or 2nd instars. Then each individual larva was placed in a Petri dish of cornmeal until it became the appropriate instar stage. For example in the 1st to 3rd instar retention experiment, the larvae were conditioned during the 1st instar and tested right after conditioning. They were then placed into cornmeal for two days until they became 3rd instars. They were then retested for associative learning in the 3rd instar.
The study was carried out in the Department of Biology, University of Kentucky from 2009-2019 and the genetic lines were produced in the School of Biological Sciences and Technology, Chonnam National University, Gwangju, 61186, Republic of Korea in 2008-2009.
RESULTS
Conditioning in the larval stages: Background UAS-TRPV1 Drosophila larvae were tested individually in the developing larval stages. Figure 3 summarizes the conditioning method used in this experiment. Larvae are exposed to the banded stripes for 2 min and transferred to the fructose agar dish with light exposure for 2 min. The larvae were then transferred to the dish with quinine hemisulfate for 2 min in a dark condition. The exchange back and forth to the fructose and quinine hemisulfate agar dishes occurred 5 times with the associated lighting conditions. Afterwards, the larvae were exposed to a banded dish of light and dark stripes. As shown in Fig. 4a, the 1st instar larvae show a higher preference for light when receiving Light+/dark - conditioning than the Light-/dark + conditioning. This difference can be seen by the median learning index (LI) being above 0 indicating associative learning. However, the LI is not statistically significant. The 2nd instar larvae show preferences for dark in both types of conditioning (Fig. 4b). The light+/dark - conditioning is statistically significant (p<0.05). The difference of the two conditioning can be seen in the LI, which is below 0 indicating no associative learning.
Figure 4c illustrates that the early 3rd instar larvae show a preference for dark in both conditioning. Light-/Dark+ conditioning is statistically significant (p<0.01) to show preference for the dark. The median LI is above 0 indicating associative learning. However, the LI is not statistically significant.
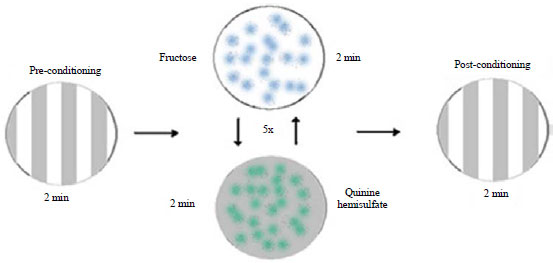 | |
| Fig. 3: | Conditioning procedure used for the background larval learning |
Retention induced with using capsaicin: Before examining the abilities to potential show associative learning in the larval stages and retention of a negative stimulus, a measure of which concentration induced negative reinforcement in the TRPV1 expressing larvae sensitive larvae was examined. A 1 nM and 1 μM capsaicin concentration was used for the conditioning method depicted in Fig. 5. The same paradigm was used as for the fructose and quinine hemisulfate agar dishes except capsaicin was exchanged for quinine hemisulfate. Separate experiments were conducted for the 1 nM or 1 μM. For the lower concentration (1 nM) there was a preference for the dark (p<0.01) in both background and TRPV1 expressing larvae. The median LI was 0, thus the larvae did not show associative learning (Fig. 6a). For the higher concentration (1 μM) the background larvae show a preference for dark (p<0.01) and with the Light-/Dark+ conditioning for the TRPV1 expressing larvae there was a stronger preference value for dark than the reciprocal conditioning (Fig. 6b). The median LI for background equals 0, thus there was no display associative learning. The median LI for the TRPV1 expressing larvae is above 0, indicating associative learning. So the 1 μM capsaicin was used for the negative reinforcement for the larval learning and retention future studies.
One time exposure to capsaicin during the larval stages: The conditioning paradigm used is depicted in Fig. 5 but using only the capsaicin at 1 μM. The results for the preference and LI for the 1st instar background and TRPV1 expressing larvae are shown in Fig. 7a. The background larva had a stronger preference for light (light-/dark+) than the light+/dark-conditioning. The TRPV1 expressing larvae did not show preference for either light or dark. The median LI equals 0 for both background and TRPV1 expressing larvae indicating no associative learning.
The preference in 2nd instar and the LI score are displayed in Fig. 7b. The background and TRPV1 expressing larvae have a preference for dark in both reciprocal training regimes. The light-/dark + conditioning for the instars in the background is statistically significant (p<0.05). The median LI value for background larvae is above 0 indicating associative learning; the median LI equals 0 for the TRPV1 expressing larvae indicating no associative learning. The preference and LI for 3rd instars is shown in Fig. 7c. The background and TRPV1 expressing larvae have a preference for dark in both reciprocal training regimes. The background strain is statistically significant (p<0.01) in both reciprocal training regimes. The TRPV1 expressing larvae are statistically significant for both the Light+/dark- and light -/dark+ (p<0.01, p<0.05, respectively). The median LI values for background larvae equals 0 indicating no associative learning (p<0.02), the median LI value for TRPV1 expressing larvae is below 0 indicating no associative learning.
Memory retention for the one-time exposure: To test memory retention, the TRPV1 expressing larvae were examined with capsaicin as the negative reinforcement. Three experiments were conducted to test retention. First is to test retention from 1st to 2nd instars after a one time exposure. The second experiment is to test retention from 1st to 3rd instars after a one time exposure. The last experiment is to test retention from 2nd to 3rd instars after a one time exposure.
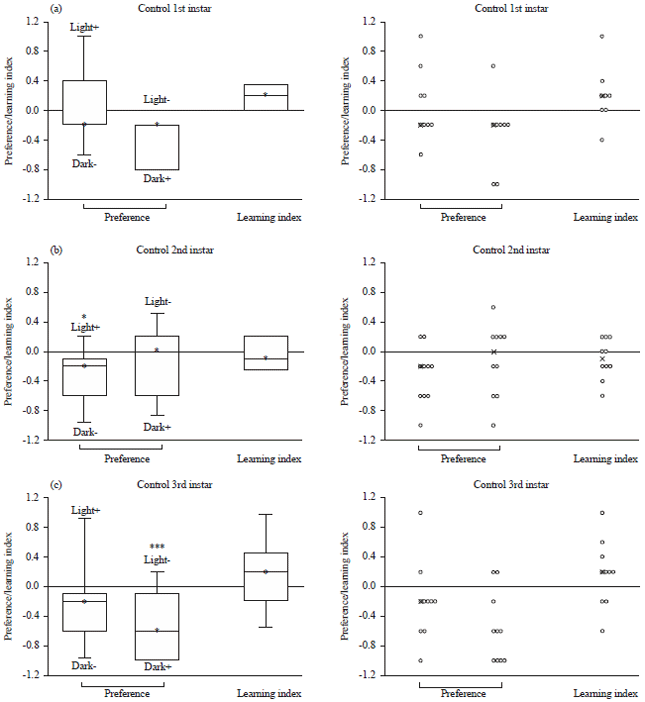 | |
| Fig. 4(a-c): | Preference and learning index for the (a) 1st, (b) 2nd and (c) 3rd instar background strain (UAS-TRPV1) strain |
| Scatter plot is located on the right, *p<0.05 for the light+/dark-, ***p<0.01 for the light-/dark+ | |
The preference from 1st to 2nd instars is shown in Fig. 8a. 1st instar background larvae show preference for light in the light-/dark+ condition. The 1st instar TRPV1 expressing larvae did not show preference for the light. In the 2nd instar stage, the background larvae showed a preference for light in the light+/dark- condition and the TRPV1 expressing larvae show a preference for light in the light-/dark+ condition (p<0.05). In the learning index, the 1st instars for both background and TRPV1 expressing larvae did not show learning due to the median of LI at 0 (Fig. 8b). The 2nd instar background larvae shows associative learning (p<0.01), but the 2nd instar TRPV1 expressing larvae did not. Thus, the learned behavior was not retained for the TRPV1 expressing larvae.
The preference from 1st to 3rd instars is represented in Fig. 9a. First instar background larvae displays a preference for dark (p<0.01) and the TRPV1 expressing larvae shows a preference for dark (p<0.05 in light-/dark+). For the 3rd instars both background and TRPV1 expressing larvae show a preference for dark (p<0.05 for background 3rd instars) and the learning index is represented by Fig. 9b.
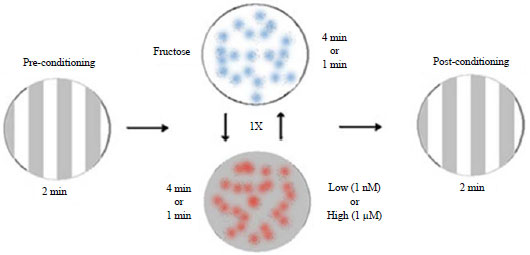 | |
| Fig. 5: | Outline for the conditioning of the 3rd instar larvae (Background and TRPV1 expressing larvae) to test the two concentrations of capsaicin |
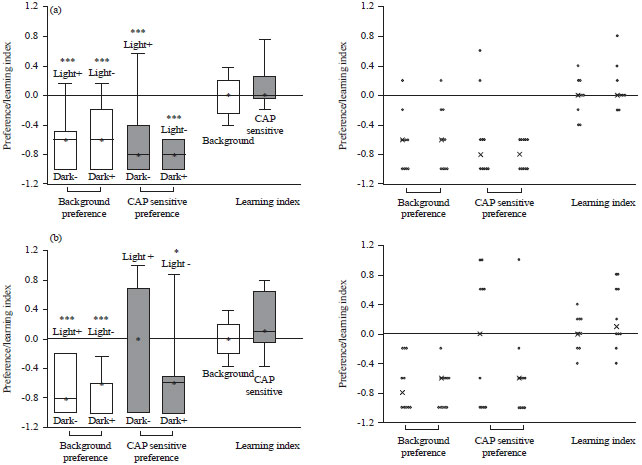 | |
| Fig. 6(a-b): | Preference and learning index for the 3rd instars background (UAS-TRPV1) and TRPV1 expressing larvae in (a) Low concentration of capsaicin (1 nM) and (b) High concentration of capsaicin (1 μM) |
| Scatter plot of the data is located on the right, ***p<0.01, CAP sensitive are the TRPV1 expressing larvae, ***p<0.01, *p<0.05 | |
First instar background and TRPV1 expressing larvae did not show associative learning. The 3rd instar background larvae did show associative learning, but TRPV1 expressing larvae did not. Thus, retention was not maintained in the TRPV1 expressing larvae from 1st to 3rd instars.
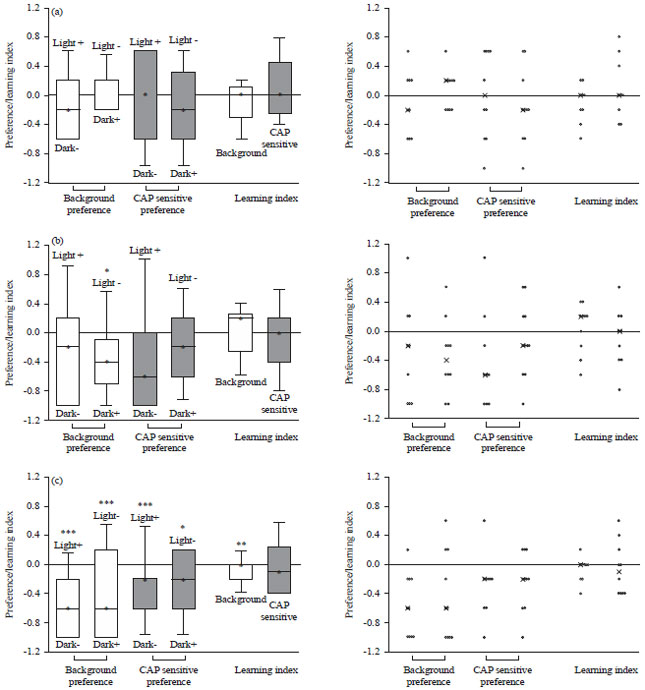 | |
| Fig. 7(a-c): | Preference and learning index of (a) 1st, (b) 2nd and (c) 3rd instar background and TRPV1 expressing larvae |
| Scatter plot is located on the right, CAP sensitive are the TRPV1 expressing larvae, *p<0.05 CAP sensitive are the TRPV1 expressing larvae, ***p<0.05, **p<0.02, *p<0.01 | |
The preference values for larvae 2nd to 3rd instars is shown in Fig. 10a. The 2nd instar background and TRPV1 expressing larvae did not show preference for light (p<0.05 for background larvae light-/dark+). Also, the 3rd instar background and TRPV1 expressing larvae did not show preference for light (background larvae light+/dark- p<0.01, background larvae and TRPV expressing larvae light-/dark+ p<0.05). Learning index is shown in Fig. 10b which illustrates the 2nd instar background larvae show associative learning. However, 2nd instar TRPV1 expressing larvae as well as the 3rd instar background and TRPV1 expressing larvae did not show associative learning. Thus, TRPV1 expressing larvae did not retain the learned behavior.
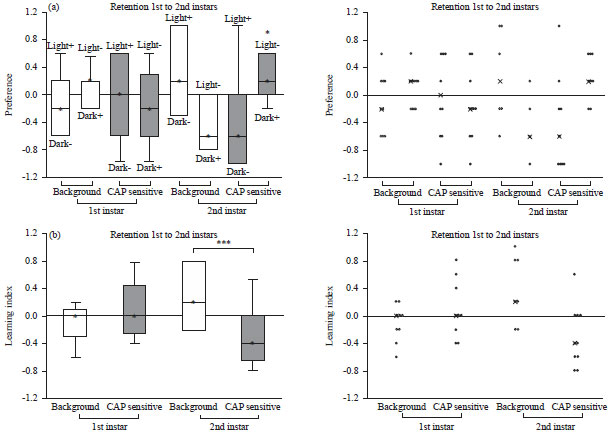 | |
| Fig. 8(a-b): | (a) Preference and (b) Learning index of 1st and 2nd instar background and TRPV1 expressing larvae |
| Scatter plot of the data is located on the right, *p<0.05, ***p<0.01, CAP sensitive are the TRPV1 expressing larvae | |
 | |
| Fig. 9(a-b): | (a) Preference and (b) Learning index of 1st and 3rd instar background and CAP sensitive larvae |
| *p<0.05, **p<0.02, ***p<0.01, CAP sensitive are the TRPV1 expressing larvae, scatter plot of the data is located on the right | |
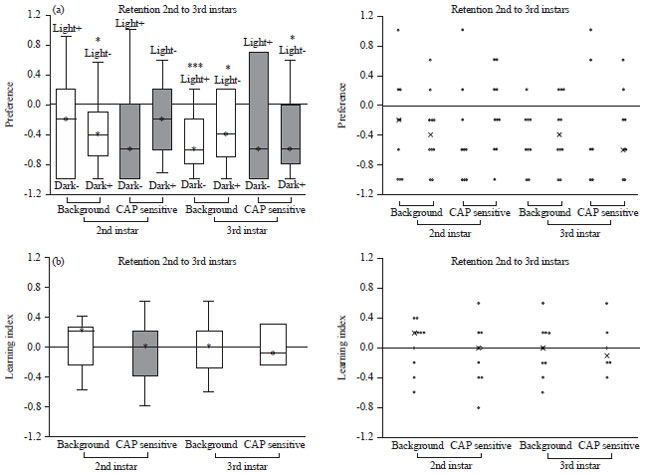 | |
| Fig. 10(a-b): | (a) Preference and (b) Learning index of 2nd and 3rd instar background and CAP sensitive larvae |
| Scatter plot of the data is located on the right, *p<0.05, ***p<0.01, CAP sensitive are the TRPV1 expressing larvae | |
DISCUSSION
This study demonstrates that in the 1st and 3rd larval stages of the background strain (UAS-TRPV1) strain show abilities to have a preference when associating visual and gustatory reinforcements. The transgenic Drosophila strain that possesses an enhanced sensory perception (capsaicin sensitive Drosophila) shows abilities to also have a preference when the negative reinforcement is at a concentration at 1 μM during the 3rd larval stage. When tested with a one-time exposure to the reinforcements during the developing stages the larvae did not retain a preference; thus, they did not retain an association. The TRPV1 expressing larval stages did not demonstrate an ability to learn with the one-time exposure to the reinforcements. The TRPV1 expressing larvae did not show any retention of the associated preference.
Since the reciprocal conditioning was also tested (light+/dark- and light-/dark+), associative learning is shown between visual stimuli and gustatory reinforcements. As stated by Gerber et al.16, fructose is an effective reinforcement; however, quinine sulfate is not as effective. Using the negative reinforcement of capsaicin for the TRPV1 expressing larvae did serve as a useful negative reinforcement because of the high sensitivity. Future studies could address varying the concentration of capsaicin to potential address if a stronger stimulus might induce a memory retention. Capsaicin cannot be used as a negative reinforcement for the background (UAS-TRPV1) since the larvae did not show a response when placed on capsaicin agar plates.
Based on the results in this study, the larval stages in background larvae showed variance in learning. The background larval strains for the TRPV1 expressing larvae learned as 2nd instars. The brain sizes of 1st, 2nd and 3rd instars of the wild type strain are remarkably different in size17,18. Thus, the variance of learning ability for TRPV expressing larvae throughout development could be due to remodeling of neuronal structures. In addition, it is not known if the manipulations in genomic sequence for the background and TRPV1 expressing larvae have pleiotropic actions on neural development or cellular function19. It would not be surprising if activation of the sensory neurons expressing TRPV1 resulted in a heightened stress for the larvae and alteration in release of hormones or biogenic amines possibly altered mental status occurs and the larvae cannot focus on the associative learning20. Testing other mutants with learning deficits, such as dunce and amnesic strains, have shown that adults do possess abilities to learn; however, memory is affected21. It would be of interest if the larval stages for dunce and amnesic strains show some ability to learn and if they have the ability to retain a learned behavior.
Since there are multiple ways to examine learning in adults, such as gustatory, visual, or olfactory, a future study could be to condition the TRPV1 expressing larvae with visual conditions and heat. Since the TRPV1 receptors are receptors for capsaicin and well as heat, heat could be examined as a negative reinforcement in this same line. In addition, it would be of interest to know if a learning ability could be retained through metamorphosis from the larvae to the adult using an over expressing strain for TRPV1 in sensory neurons. Such studies could provide additional information in how and which neural circuits retain learning and memory during this developmental transition.
CONCLUSION
This study demonstrated that preference to a mild favorable or mild noxious stimulus is present throughout larval development in genetically engineered strain of Drosophila. This preference is still present when larvae are paired with light preference in larvae not expressing the TRPV1 as well as ones expressing the TRPV1 but with exposure to capsaicin the larvae do not retain the associative learning with one trail or multiple trails of association.
ACKNOWLEDGMENTS
Funding was provided by Kentucky Young Researchers Program at the University of Kentucky (A.S.O.) and NSF-IBN-0131459.
SIGNIFICANCE STATEMENT
This is significant as associate learning experiments to noxious stimuli are shown that a preference to a stimulus maybe observed but the retention to a strongly aversive stimulus may not be retained.
REFERENCES
- Rubin, G.M., M.D. Yandell, J.R. Wortman, G.L. Gabor and Miklos et al., 2000. Comparative genomics of the eukaryotes. Science, 287: 2204-2215.
CrossRefDirect Link - Tully, T. and W.G. Quinn, 1985. Classical conditioning and retention in normal and mutant Drosophila melanogaster. J. Comp. Physiol. A, 157: 263-277.
CrossRefDirect Link - Aceves-Pina, E.O. and W.G. Quinn, 1979. Learning in normal and mutant Drosophila larvae. Science, 206: 93-96.
CrossRefDirect Link - Gerber, B. and R.F. Stocker, 2007. The Drosophila larva as a model for studying chemosensation and chemosensory learning: A review. Chem. Senses, 32: 65-89.
CrossRefPubMedDirect Link - Diegelmann, S., B. Klagges, B. Michels, M. Schleyer and B. Gerber, 2013. Maggot learning and Synapsin function. J. Exp. Biol., 216: 939-951.
CrossRefDirect Link - Thum, A.S. and B. Gerber, 2019. Connectomics and function of a memory network: The mushroom body of larval Drosophila. Curr. Opin. Neurobiol., 54: 146-154.
CrossRefDirect Link - Stocker, R.F., 1994. The organization of the chemosensory system in Drosophila melanogaster: A review. Cell Tissue Res., 275: 3-26.
CrossRefDirect Link - Stocker, R.F., 2001. Drosophila as a focus in olfactory research: Mapping of olfactory sensilla by fine structure, odor specificity, odorant receptor expression and central connectivity. Microsc. Res. Tech., 55: 284-296.
CrossRefDirect Link - Scherer, S., R.F. Stocker and B. Gerber, 2003. Olfactory learning in individually assayed Drosophila larvae. Learn. Memory, 10: 217-225.
CrossRefDirect Link - Rock, I. and W. Heimer, 1959. Further evidence of one-trial associative learning. Am. J. Psychol., 72: 1-16.
CrossRefDirect Link - Roediger, III H.L. and K.M. Arnold, 2012. The one-trial learning controversy and its aftermath: Remembering. Am. J. Psychol., 125: 127-143.
PubMedDirect Link - Weiglein, A., F. Gerstner, N. Mancini, M. Schleyer and B. Gerber, 2019. One-trial learning in larval Drosophila. Learn. Memory, 26: 109-120.
CrossRefDirect Link - Schurks, M. and H.C. Diener, 2008. Migraine, allodynia and implications for treatment. Eur. J. Neurol., 15: 1279-1285.
CrossRefDirect Link - Hendel, T., B. Michels, K. Neuser, A. Schipanski and K. Kaun et al., 2005. The carrot, not the stick: Appetitive rather than aversive gustatory stimuli support associative olfactory learning in individually assayed Drosophila larvae. J. Comp. Physiol. A, 191: 265-279.
CrossRefDirect Link - Badre, N.H. and R.L. Cooper, 2008. Reduced calcium channel function in drosophila disrupts associative learning in larva and behavior in adults. Int. J. Zool. Res., 4: 152-164.
CrossRefDirect Link - Gerber, B., S. Scherer, K. Neuser, B. Michels, T. Hendel, R.F. Stocker and M. Heisenberg, 2004. Visual learning in individually assayed Drosophila larvae. J. Exp. Biol., 207: 179-188.
CrossRefPubMedDirect Link - Hartenstein, V., S. Spindler, W. Pereanu and S. Fung, 2008. The development of the Drosophila larval brain. Adv. Exp. Med. Biol., 628: 1-31.
CrossRefDirect Link - Pereanu, W. and V. Hartenstein, 2006. Neural lineages of the Drosophila brain: A three-dimensional digital atlas of the pattern of lineage location and projection at the late larval stage. J. Neurosci., 26: 5534-5553.
CrossRefDirect Link - Rollmann, S.M., L. Zwarts, A.C. Edwards, A. Yamamoto and P. Callaerts et al., 2008. Pleiotropic effects of Drosophila neuralized on complex behaviors and brain structure. Genetics, 179: 1327-1336.
CrossRefDirect Link - Dasari, S. and R.L. Cooper, 2004. Modulation of sensory-CNS-motor circuits by serotonin, octopamine and dopamine in semi-intact Drosophila larva. Neurosci. Res., 48: 221-227.
CrossRefPubMedDirect Link - Gong, Z.F., S.Z. Xia, L. Liu, C.H. Feng and A.K. Guo, 1998. Operant visual learning and memory in Drosophila mutants dunce, amnesiac and radish. J. Insect Physiol., 44: 1149-1158.
CrossRefDirect Link