
Humayra Akter
Institute of Food and Radiation Biology, Atomic Energy Research Establishment, Bangladesh Atomic Energy Commission, Dhaka, Bangladesh
Shakil Ahmed Khan
Institute of Food and Radiation Biology, Atomic Energy Research Establishment, Bangladesh Atomic Energy Commission, Dhaka, Bangladesh
Journal of Entomology
Year: 2014 | Volume: 11 | Issue: 2 | Page No.: 56-67







ABSTRACT
Aedes aegypti is a cosmopolitan urban mosquito that causes dengue every year in Bangladesh. The present study was carried out to observe the sensitivity level of immature stages of A. aegypti mosquito to gamma irradiation. Different developmental stages of A. aegypti were exposed to a series of irradiation doses in Co60 gamma source to observe sensitivity regarding egg hatching, pupation, adult emergence, mortality and body size. Irradiation doses of 1-10 Gray were applied to eggs, 10-100 Gy to larvae and 10-250 Gy were applied to pupae. Egg hatching, pupae formation and adult emergence decreased with increasing dose (p ’ 0.000). Pupation decreased significantly (p ’ 0.000) with increasing radiation dose in 1st, 2nd, 3rd and 4th instar larvae. Regression analysis showed increase of percent mortality with increasing dose significantly in both 19-23 h old (early) and 42-46 h old (late) pupae (p ’ 0.000). A linear positive relationship was found between doses and mortality in both larvae and pupae. Irradiation of early and late pupae had no significant effect on adult emergence up to 40 Gy, however, while higher doses applied (100-250 Gy) emergence rate decreased significantly (p ’ 0.000). Lethal dose, LD50 and LD90 for 4th instar larvae were lower than pupae but higher than eggs. Even, LD50 and LD90 for early pupae were lower than late pupae. There was no significant effect of radiation on wing length compared to control (unirradiated) which were exposed to radiation at pupal stage (p ’ 0.153). Overall radiation had effect on egg hatching, pupation, adult emergence, mortality but had no effect on adult’s body size of Aedes aegypti.
PDF Abstract XML References Citation
Received: July 07, 2013;
Accepted: September 21, 2013;
Published: March 08, 2014
How to cite this article
Humayra Akter and Shakil Ahmed Khan, 2014. Sensitivity of Immature Stages of Dengue Causing Mosquito, Aedes aegypti
(L.) to Gamma Radiation. Journal of Entomology, 11: 56-67.
DOI: 10.3923/je.2014.56.67
URL: https://scialert.net/abstract/?doi=je.2014.56.67
DOI: 10.3923/je.2014.56.67
URL: https://scialert.net/abstract/?doi=je.2014.56.67
INTRODUCTION
The dengue causing mosquito, Aedes aegypti has been reported from many countries including Bangladesh. In recent years people are suffering from dengue in Bangladesh at alarming rate. Two well known vectors of dengue namely A. aegypti and A. albopictus have been reported from Bangladesh (Aziz et al., 1967; Ameen and Moizuddin, 1973). Dengue is a serious problem in Bangladesh, every year people are attacked by this disease. Sometimes it is difficult to control by indoor insecticide spraying for the human health safety; outdoor spraying is continuing to suppress this vector. However, insecticide is also hazardous for environment. Although, vector control programme in several countries were based on chemical strategies, unrestricted and prolonged use of insecticides led to resistance development to this vector and even to the dengue causing virus for many conventional insecticides in many countries (Ansari et al., 2005; Ali et al., 1999; Karunaratne and Hemingway, 2001; Hemingway and Ranson, 2000; Selvi et al., 2005; Mulla, 1995; Tikar et al., 2008). Therefore, alternative vector control method like botanical insecticides, Sterile Insect Technique (SIT) and integrated pest management techniques have been considering to combat this problem (Singh et al., 2006; Almehmadi, 2011). Although, SIT has been applied successfully in new world screwworm, Cochliomyia hominivorax and the Mediterranian fruit fly, Ceratitis capitata (Alphey, 2002), for mosquito control SIT is new technique and trials have been going on in different countries (Benedict and Robinson, 2003).
Before going to control any pest or vector, effect of control measure on its different developmental stages should be know at first. In the development of mosquito its environmental factors like temperature, density and availability of food plays an important role (Gomes et al., 1995; Hawley, 1988; Mori, 1979). Feeding types and larval density may also influence mortality rate and adult body size (Mori, 1979). Dengue causing virus is carried by A. Aegypti mosquito the control of which has become an important concerning issue for the government of Bangladesh as every year enormous number of dengue patients are admitted in hospitals. Still chemical spray has been using along with the use of mosquito net during day time in dengue season to prevent A. aegypti biting. However, it is not permanent solution; long term persistent control of this vector is crucial. For this reason, radiosensitivity research is going on in Bangladesh. Without the prior knowledge on sensitivity to irradiation SIT development is impossible. This present study was undertaken to observe the response of different immature stages of A. aegypti to different radiation doses. The objective was to test whether radiation has any effect on adult emergence, mortality and adult body size once they had been irradiated at immature stages.
MATERIALS AND METHODS
This research work was carried out in Insect Biotechnology Division, Institute of Food and Radiation Biology (IFRB), Atomic Energy Research Establishment (AERE), Savar, Dhaka from November 2011 to March 2013. In this experiment irradiation dose of 1-10 Gray were applied to eggs and 10-100 Gy to larvae; 10-250 Gy were applied to pupae in Co60 gamma source. Mosquitoes were obtained from the laboratory colony of Aedes aegypti reared in a large square cage (56x56x56 cm) made with aluminum frame and the mosquito nets in the insectary of IFRB, AERE, Savar, Dhaka, Bangladesh. The laboratory colony of Aedes aegypti was maintained at 26±1°C, 60-70% Relative Humidity (RH) and 12 h light; 12 h dark condition.
Collection and irradiation of eggs: To collect eggs for irradiation an artificial modified membrane blood feeding technique was used to feed female mosquito (Mourya et al., 2000; Mishra et al., 2005). Chicken blood was collected from local market in blood bag containing anticoagulant substrate (Latrate Phosphid dextrose). Mosquitoes were fed blood three consecutive days. After blood feeding the female mosquitoes laid eggs on a moistened paper strip that was attached inside the beaker containing water in such a way that about half part of the paper strip should remain beneath the water level. Egg strips were dried and stored in covered petridish and a small vial with wet tissue was put in to avoid desiccation of eggs. About 15-30 days aged eggs were irradiated in petridish along with moistened tissue to avoid desiccation. For each dose about 1/4th inches of egg strip was used. Experiment was replicated 3 times.
Irradiation of larvae and pupae: Dosimetry confirmed that all doses delivered lay within a 5% error range. Both larvae and pupae of A. aegypti were irradiated in small plastic lid vial containing water. For each dose 20 insects were used. After irradiation insects of each dose were transferred to 100 mL beaker containing 50 mL water and were kept in a bigger beaker (250 mL) and were covered with mosquito net to avoid outgoing of adults and/or incoming of other insects. Food was provided daily for proper development. The development of larva to its next stage and to pupae and adult was observed and the number of insects in each stage was recorded to calculate the percent mortality at treatment stage and percent adult emergence at different doses. In this way, 7 replicates were done for each larval instars and pupae.
All larval instars and two groups of pupae of A. aegypti were irradiated; one of within 19-23 h old aged pupae (early) and another of within 42-46 h old aged pupae (late). Pupae were collected the day before irradiation at 3 p.m. from trays that had been cleared of all pupae before 11 a.m. At 10 a.m. the next day, the pupae, aged 19-23 h and at 9 a.m. on the following day 42-46 h old aged pupae were irradiated.
Lethal dose selection: Lethal dose for egg, larva and pupa was selected by using standard protocol. Insects those could not go to next stage after certain dose application within 24 h were counted and subjected to probit analysis (Finney, 1947). The dose at which 50 and 90% insect population was died selected as LD50 and LD90.
Egg hatching, pupation and adult emergence observation: After irradiation eggs were dipped in water in plastic cup to observe hatching. Following hatching larvae and eggs were counted; hatching percentage was calculated and kept for pupation and adult emergence rate observation. After irradiation larvae and pupae were kept in beaker with water covered by mosquito net. Every day it was checked whether insects become pupae or adults. From the day of pupation in 3 consecutive days pupae were counted. After pupae formation, usually within 53-59 h adults were emerged. Both pupal recovery and adult emergence were recorded as percentage.
Mortality observation: After irradiation during the developmental period mortality of insects was recorded at two stages; firstly when irradiated insects developed to just next stage (immediate mortality) and secondly when they developed to adult (gradual mortality). The number of dead insects were counted and calculated for percent mortality.
Body size: In case of mosquito body size is measured by the measurement of wing length. Wings of adult were measured under stereo microscope. In total, 10 adults were selected from 2 batches for each dose to measure wing length.
Statistical analysis: One way ANOVA was performed for significance level studies using statistical software MINITAB, USA (Version 13.1), with DF 5 (degrees of freedom) and Tukey,s family error rate 0.05. Graph was created in Microsoft Excel 2007 and Sigma Plot software. Probit analysis was performed in EPA PROBIT ANALYSIS PROGRAM software (Version 1.5).
RESULTS
Mosquito survival: Lethal Dose (LD) for A. aegypti varied within eggs, larvae and pupae; from the probit analysis it is observed that LD50 and LD90 for eggs were13.107 and 23.760; for 4th instar larva were 68.49 and 307.76 Gy (Table 1).
| Table 1: | Susceptibility of different stages of Aedes aegypti (mortality within 24 h) to gamma irradiation (Gray) |
 | |
| Eggs are most susceptible to radiation among eggs, larvae and pupae as least radiation dose was required for 50 and 90% population death (LD50 LD90), for larvae comparatively less radiation dose was required for 50 and 90% population death than pupae | |
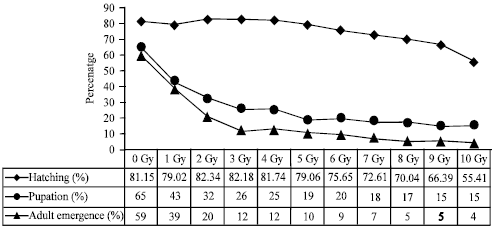 | |
| Fig. 1: | Percent hatching, pupation and adult emergence of Aedes aegypti mosquito in response to different irradiation doses (1-10 Gy) compared to control (0 Gy) which were applied on eggs |
Lethal dose varied with age of pupae; LD50 and LD90 for early pupae are 179.16 and 367.49 Gy whereas for late pupae the respective lethal doses were 265.18 and 506.53 Gy (Table 1).
Egg hatching and pupal recovery: Both egg hatching and pupation rate decreased with increasing radiation dose (p = 0.000) in irradiated eggs. In insects developed from irradiated eggs, pupae formation rate fell sharply with increased radiation dose; from unirradiated eggs 65% pupae was recovered that was reduced to 2% at 2 Gy (Fig. 1). Although from insects developed from irradiated larvae pupation did not fell like eggs but it decreased significantly (p = 0.000) with increasing radiation dose in 1st, 2nd, 3rd and 4th instar larvae (Fig. 2). In control about 83, 78, 84 and 91% larvae was developed to pupae which reduced to about 10, 26, 32 and 52% in 1st, 2nd, 3rd and 4th instars, respectively at 50 Gy (Fig. 2).
Adult emergence: Irradiation of eggs and larvae has significant effect on adult emergence. The emergence decreased with the increase of irradiation dose significantly (p = 0.000) in insects developed from irradiated eggs; adult emergence was 59% in control which was reduced to 20% at 2 Gy and 4% at 10 Gy (Fig. 1). In all larval instars, irradiation dose of 10 Gy had no significant effect on adult emergence. Adult emergence rate was below 40% in 1st and 2nd instars at 20 Gy (Fig. 2a, b); while at 40 Gy in 3rd and 4th instars (Fig. 2c, d). These results suggest that 3rd and 4th instar larvae are more radio tolerant than 1st and 2nd instar larvae. Adult emergence rate sharply fell in irradiated 1st instar larvae than other instars.
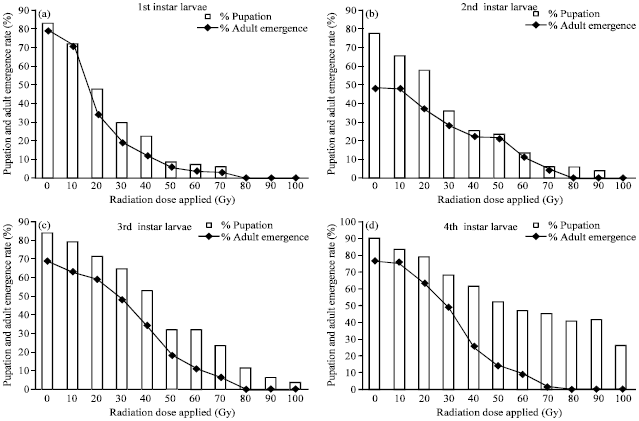 | |
| Fig. 2(a-d): | Pupae formation and adult emergence rate of Aedes aegypti mosquito in response to different irradiation doses which were applied on different larval instars. Both pupation and adult emergence decreased with increasing dose in all instars, (a) Both pupation and adult emergence decreased sharply with increasing doses and stopped at 80 Gy in 1st instar larvae; within 50 Gy it became 6% while it was 79% in control, (b) In 2nd instar larvae emergence reduced to about half (21%) of control (49%) at 50 Gy, (c) In 3rd instars emergence became about one fourth (18%) at 50 Gy compared to control (69%) and (d) In 4th instars emergence became one fifth (14%) of control (77%) at 50 Gy |
Radiation had no remarkable effect on adult emergence at 10 Gy and adult emergence was dropped to under 40% at 20 Gy (Fig. 2a). However, within 50 Gy it became 6% while it was 79% in control (Fig. 2a). In 2nd instass larvae emergence reduced to about half (21%) of control (49%) at 50 Gy (Fig. 2b). In 3rd instass emergence became about one fourth (18%) at 50 Gy compared to control (69%) (Fig. 2c). In 4th instar emergence become one fifth (14%) of control (77%) at 50 Gy (Fig. 2d). This result suggests that 1st instar is more susceptible to radiation than other instars up to 50 Gy. Till 40 Gy there was no significant reduction in adult emergence in 2nd instar (p = 0.30) and 3rd instar (p = 0.317) larvae but significant reduction was found in different 1st instar (p = 0.00) and 4th instar (p = 0.014).
Overall, emergence was high (Fig. 3) in irradiated pupae. Radiation had no significant effect on adult emergence till 40 Gy in both early and late pupae (p = 0.894 and p = 0.179 for late and early pupae), on average 90% or more emergence was observed in early and late pupae where in control it was 95%. Similar results were reported for male pupal irradiation of Aedes polynesiensis vector of Wuchereria bancrofti with 40 Gy resulted in no effect on emergence (Brelsfoard et al., 2009).
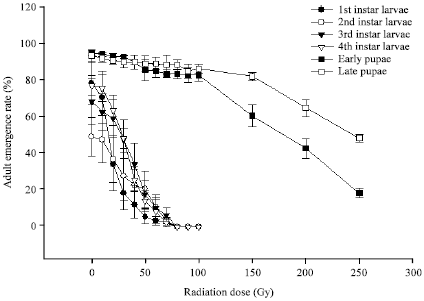 | |
| Fig. 3: | Comparison of adult emergence rate developed from larvae and pupae of Aedes aegypti irradiated at different doses. Vertical bars represent standard error. (Early pupae refers to 19-23 h; late pupae refers to 42-46 h aged) |
However, up to 40 Gy emergence significantly differed between early and late pupae (p = 0.045). Significant differences were observed between adult emergence for the early and late pupae up to 100 Gy (p = 0.001). In 100 Gy emergence was 83 and 86% in early and late pupae, respectively. No significant difference between unirradiated and iirradiated in late pupae (p = 0.886) but emergence differed significantly between irradiated and unirradiated early pupae (p = 0.004) up to 100 Gy. Emergence started to decrease significantly when irradiation dose increased up to 250 Gy in both early and late pupae (p = 0.000) (Fig. 3). Even, emergence varied significantly between early and late pupae (p = 0.000).
Mortality: Both immediate and gradual mortality in larvae and pupae of A. aegypti increased significantly with the increasing irradiation dose (p = 0.000). There was a linear positive relationship between doses and mortality in both larvae and pupae of A. aegypti (R2 = 0.96, 0.98, 0.88, 0.94 for immediate mortality in 1st, 2nd, 3rd and 4th instar larvae, respectively; R2 = 0.74, 0.95, 0.93, 0.89 for gradual mortality in 1st, 2nd, 3rd and 4th instars, respectively and R2 = 0.93 and 0.87 in early and late pupae, respectively) (Fig. 4a-d and 5). In all larval instars 100% mortality was achieved at 80-100 Gy (Fig. 4a-d). No significant differences were found in mortality between irradiated 1st and 2nd instar larvae up to 50 Gy. From 60 Gy with the increasing dose the difference was noticed. Immediate mortality in 1st instar larvae was higher than 2nd instar larvae whereas gradual mortality was same in 1st and 2nd instar larvae (100%). One way ANOVA showed this increased percent mortality with increasing dose in pupae is significant in both early and late pupae (p = 0.000). From this experiment it is also revealed that early pupae are significantly more susceptible to irradiation than late pupae (p = 0.000) (Fig. 5).
Body size: Over all, there was no significant difference in wing length between irradiated and unirradiated adult males those were exposed to radiation at pupal stage (p = 0.153).
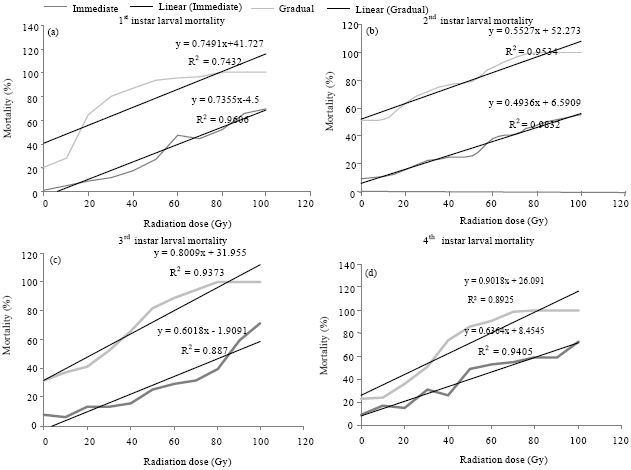 | |
| Fig. 4(a-d): | Sensitivity of Aedes aegypti mosquito to gamma radiation; immediate (immediately after irradiation) and gradual (total mortality during developmental period up to adult emergence) mortality of different larval instars in response to different radiation doses. In all instars immediate mortality is lower than gradual mortality |
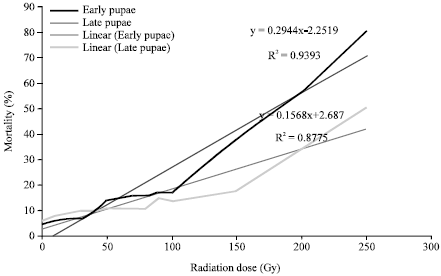 | |
| Fig. 5: | Sensitivity of Aedes aegypti mosquito to gama radiation; effect of radiation on pupae (Early pupae refers to 19-23 h; late pupae refers to 42-46 h aged) |
 | |
| Fig. 6: | Effect of irradiation on the body size of Aedes aegypti. Vertical bars represent standard errors (SE) |
In control wing length was 2.29±0.05 mm and in irradiated it varied from 2.16±0.06 to 2.32±0.05 mm (Fig. 6). Therefore, this result suggests that radiation has no effect on body size of A. aegypti.
DISCUSSION
Radiation has immediate effect on larvae and pupal survival. Lethal dose LD50 and LD90 for 4th instar larvae were higher than eggs and lower than that of early and late pupae. Egg hatch decreased significantly with increasing dose compared to control (p = 0.000). There is example of such hatching trend of eggs produced from females those were irradiated at pupal stage (Brelsfoard et al., 2009).
Present study revealed that radiation has effect on larval and pupal mortality. The mortality of the immatutre stages during mosquito growth is a very well-known fact that depends mainly on density, food supply and temperature; even 100% mortality may occur in all stages of A. albopictus at 46.1°C (Chan et al., 1971; Hawley, 1988; Smith et al., 1988). In present study radiation caused significant mortality in larvae compared to control and from 80-100 Gy 100% gradual mortality (%mortality upto adult emergence) was observed in all larval instars (Fig. 4a-d). Therefore, it could be stated inspite of having sufficient food supply and controlled temperature the immatutre stages of A. aegypti may die only because of irradiation. However, such results were not observed in pupae; radiation did not have any noticeable effect on pupae up to 100 Gy. Higher doses had effect on mortality (Fig. 5). In present study late larval instars (3rd and 4th instars) showed almost similar or in some doses more susceptible to radiation than 1st and 2nd instar larvae (Fig. 4a-d). This result, though unusual, has resemblance with the study of Gomes et al. (1995) in Brazil. They also found higher mortality in 3rd and 4th instar larvae than in the 2nd instar while no mortalities in 1st instar in the bamboo. Unlike larvae, late pupae were more radio tolerant than early pupae in terms of adult emergence and mortality (Fig. 3, 5).
Adult emergence rates were used as a measure of survivorship after irradiation as eggs, larvae and pupae (Brelsfoard et al., 2009). Low dose gamma radiation (1-10 Gy) had significant effect on pupation and adult emergence when they were irradiated as eggs (p = 0.0001) (Fig. 1). A series of irradiation doses of up to 100 Gy had significant effect on pupation and adult emergence when they were irradiated as larvae (p = 0.0001) (Fig. 2). Overall, emergence was high (Fig. 3) in irradiated pupae. Up to 100 Gy non-significant effect was observed when they were irradiated as early and late pupae which had high adult emergence rate (>85%) (Fig. 3). Radiation had no significant effect on adult emergence till 40 Gy in both early and late pupae (p = 0.894 and p = 0.179 for late and early pupae, respectively). On average 90% or more emergence was observed in early and late pupae where in control emergence was 95%. Similar results were reported for male pupal irradiation of Aedes polynesiensis vector of Wuchereria bancrofti with 40 Gy resulted in no effect on emergence (Brelsfoard et al., 2009). Present results also coincide with the previous studies of Abdel-Malek et al. (1967) and Helinski et al. (2006) who reported no significant effect of irradiation and high (>90) adult emergence rates when mosquitoes were irradiated as 16 h old or older pupae. Most experimental work on mosquito irradiation has focused on pupae specially on older (>15 h) pupae as young pupae irradiation results in a reduced emergence (Curtis, 1976; Abdel-Malek et al., 1967, 1975,; Sharma et al., 1978; Ali and Rozenboom, 1972; Tantawy et al., 1967). However, when irradiation doses increased up to 250 Gy for early and late pupae, it had significant effects on adult emergence rates (p = 0.000) (Fig. 3) compared to control mosquito.Adult emergence rate differed between eggs, larvae and pupae when compared. Within 10 Gy adult emergence reduced to only 4% in eggs while it varied from 48-76% in 1st to 4th instar larvae. At 80 Gy adult was stopped to emerge from irradiated larvae (Fig. 1, 2). On the other hand up to 100 Gy high rate of (>85%) adult emergence was observed from irradiated pupae (Fig. 3). This result suggests that larvae are more susceptible to irradiation than pupae when they are treated with up to 100 Gy radiation. Results of adult emergence infer more radio tolerance capacity of pupae (early or late) compared to eggs and larvae. Almost similar result was found by Helinski et al. (2006). Not only that, from the present investigation it is also revealed that early pupae are more susceptible to radiation compared to late pupae in terms of adult emergence (p = 0.000) (Fig. 2) which coincide with the statement of Dame et al. (2009). This might be because of the fewer somatic cells and a more developed germ line presence in older pupae, resulting in lessened effects of irradiation (Asman and Rai, 1972; Curtis, 1971). Present results stated that irradiation of pupae has no significant effect on adult emergence compared to control till 100 Gy. From previous studies it was also reported that irradiation of pupae had no effect on the emergence up to 100 Gy in Anopheles arabiensis and Anopheles pharoensis (Helinski et al., 2006; Abdel-Malek et al., 1967).
Male body size is important as usually they are released in field in SIT programme. If body size reduces due to irradiation it would be unfavorable for SIT as sterile males have to compete with unirradiated wild males for mating. A. Aegypti females emerging from field-collected pupae had an average wing length of 2.47±0.20 mm (Nasci, 1986) although adult body size usually varied among species of mosquitoes with habitats (Chadee, 1993; Gomes et al., 1995; Nasci, 1986). In present study wing length was 2.29±0.05 mm in control and in irradiated it varied from 2.16±0.06-2.32±0.05 mm. As reported in many countries due to irradiation somatic cell damage may occur along with germ cell (Asman and Rai, 1972; Curtis, 1971; Proverbs, 1969) for which longevity could decrease. However, this kind of somatic cell damage might not occur in all stages of irradiated insects. This could be the reason of decreasing longevity significantly when early pupae were irradiated but absolutely no such effect of irradiation on late pupae (Brelsfoard et al., 2009). Similarly in present study it did not show any radiation effect up to 100 Gy on wing length that means these radiation doses might not damage somatic cell. In SIT programme sterile male should have similar strength as wild male to compete for mating with wild female and adult’s body size may affect the mosquitoes’ vectorial capacity (Landry et al., 1988; Nasci, 1986). Thus present finding is promising as radiation will not hamper the competitiveness of male with the wild male due to body size for mating with female. Also there will not possibility of being infected more by virus as stated before that larger A. aegypti females were significantly more susceptible to infection by Ross River Virus than smaller ones (Nasci and Mitchell, 1994).
To sum up, it is evident that radiation has effect in each developmental stage of the life cycle of Aedes aegypti mosquito. From egg hatching to adult emergence radiation showed its effect by reducing the percentage (p = 0.000). In susceptibility test eggs were affected more with radiation dose compared to larvae and pupae, thus, LD50 and LD90 for eggs were lower than larvae and pupae. In addition, only at 2 Gy pupae formation and adult emergence rate became about half and one third of control, respectively. A linear positive relationship was found between doses and mortality in both larvae and pupae. There remained no significant effect on adult emergence up to 40 Gy in early and late pupae, however, at higher doses (100-250 Gy) emergence rate decreased significantly (p = 0.000). Over all, there was no significant effect of radiation on wing length compared to control (unirradiated) which were exposed to radiation at pupal stage (p = 0.153). The results could be useful in assessing irradiation doses applied to Aedes aegypti prior to go for Sterile Insect Technique (SIT). Moreover it could be useful in indicating which immature stage will be perfect to exploit for SIT considering body size, adult emergence and most importantly mortality.
REFERENCES
- Abdel-Malek, A.A., A.O. Tantawy and A.M. Wakid, 1967. Studies on the eradication of Anopheles pharoensis by the sterile-male technique using cobalt-60. 3. Determination of the sterile dose and its biological effects on different characters related to "fitness" components. J. Econ. Entomol., 60: 20-23.
PubMedDirect Link - Abdel-Malek, A.A., A.M. Wakid, A.O. Tantawy and L.M. El-Gazzar, 1975. Studies on factors influencing the induction of sterility in Anopheles pharoensis Theobald by gamma radiation. Proceedings of the Symposium on the use of Isotopes and Pesticides in Pest Control, March 1974, Beirut-Lebanon, pp: 161-174.
- Ali, A., M.A. Chowdhury, M.I. Hossain, Mahmud-ul-Ameen, D.B. Habiba and A.F.M. Aslam, 1999. Laboratory evaluation of selected larvicides and insect growth regulators against field-collected Culex quinquefasciatus larvae from urban Dhaka, Bangladesh. J. Am. Mosq. Control Assoc., 15: 43-47.
PubMed - Almehmadi, R.M., 2011. Larvicidal, histopathological and ultra-structure studies of Matricharia chamomella extracts against the rift valley fever mosquito Culex quinquefasciatus (Culicidae: Diptera). J. Entomol., 8: 63-72.
CrossRefDirect Link - Ameen, M. and M. Moizuddin, 1973. Bionomics of the common mosquitoes of Dacca. J. Natl. History, 7: 1-21.
CrossRef - Ansari, M.A., R.K. Razdan and U. Sreehari, 2005. Laboratory and field evaluation of Hilmilin against mosquitoes. J. Am. Mosq. Control Assoc., 21: 432-436.
PubMed - Alphey, L., 2002. Re-engineering the sterile insect technique. Insect Biochem. Mol. Biol., 32: 1243-1247.
CrossRefDirect Link - Asman, M. and K.S. Rai, 1972. Developmental effects of ionizing radiation in Aedes aegypti. J. Med. Entomol., 9: 468-478.
PubMedDirect Link - Benedict, M.Q. and A.S. Robinson, 2003. The first releases of transgenic mosquitoes: An argument for the sterile insect technique. Trends Parasitol., 19: 349-355.
CrossRef - Brelsfoard, C.L., W.S. Clair and S.L. Dobson, 2009. Integration of irradiation with cytoplasmic incompatibility to facilitate a lymphatic filariasis vector elimination approach. Parasites Vectors, Vol. 2.
CrossRef - Chadee, D.D., 1993. Size of emerging and host-seeking Aedes aegypti and the relationship to containers and blood-feeding success in Trinidad, West Indies. Bull. Soc. Vector Ecol., 18: 105-108.
Direct Link - Chan, K.L., B.C. Ho and Y.C. Chan, 1971. Aedes aegypti (L.) and Aedes albopictus (Skuse) in Singapore City. Bull. World Health Organ., 44: 629-633.
Direct Link - Dame, D.A., C.F. Curtis, M.Q. Benedict, A.S. Robinson and B.G.J. Knols, 2009. Historical applications of induced sterilisation in field populations of mosquitoes. Mal. J., Vol. 8.
CrossRef - Gomes, A.D.C., S.L.D. Gotlieb, C.C. de Azevedo Marques, M.B. de Paula and G.R.A.M. Marques, 1995. Duration of larval and pupal development stages of Aedes albopictus in natural and artificial containers. Rev. Saude Publica, Vol. 29.
CrossRef - Helinski, M.E.H., A.G. Parker and B.G.J. Knols, 2006. Radiation-induced sterility for pupal and adult stages of the malaria mosquito Anopheles arabiensis. Malaria J., Vol. 5.
CrossRef - Hemingway, J. and H. Ranson, 2000. Insecticide resistance in insect vectors of human disease. Ann. Rev. Entomol., 45: 371-391.
CrossRef - Hawley, W.A., 1988. The biology of Ae.albopictus. Am. Mosquito Control Assoc., 4: 1-39.
PubMedDirect Link - Karunaratne, S.H. and J. Hemingway, 2001. Malathion resistance and prevalence of the malathion carboxylesterase mechanism in populations of mosquito vectors of disease in Sri Lanka. Bull. World Health Organ., 79: 1060-1064.
Direct Link - Landry, S.V., G.R. DeFoliart and D.B. Hogg, 1988. Adult body size and survivorship in a field population of Aedes triseriatus. J. Am. Mosq. Contr. Assoc., 4: 121-128.
Direct Link - Mishra, K., R.D. Kumar, R.K. Hazra and A.P. Das, 2005. A simple, artificial-membrane feeding method for the radio-isotope labelling of Aedes aegypti polypeptides in vivo. Ann. Trop. Med. Parasitol., 99: 803-806.
CrossRef - Mulla, M.S., 1995. The future of insect growth regulators in vector control. J. Am. Mosq. Control Assoc., 11: 269-273.
PubMed - Nasci, R.S., 1986. The size of emerging and host-seeking Aedes aegypti and the relation of size to blood-feeding success in the field. J. Am. Mosq. Control Assoc., 2: 61-62.
PubMedDirect Link - Nasci, R.S. and C.J. Mitchell, 1994. Larval diet, adult size and susceptibility of Aedes aegypti (Diptera: Culicidae) to infection with Ross River virus. J. Med. Entomol., 31: 123-126.
PubMedDirect Link - Proverbs, M.D., 1969. Induced Sterilization and control of insects. Ann. Rev. Entomol., 14: 81-102.
PubMed - Selvi, S., M.A. Endah, W.A. Nazni, H.L. Lee and A.H. Azahari, 2005. Resistance development and insecticide susceptibility in Culex quinquefasciatus against selection pressure of malathion and permethrin and its relationship to cross-resistance towards propoxur. Trop. Biomed. 22: 103-113.
PubMedDirect Link - Sharma, V.P., R.K. Razdan and M.A. Ansari, 1978. Anopheles stephensi: Effect of gamma-radiation and chemosterilants on the fertility and fitness of males for sterile male releases. J. Econ. Entomol., 71: 449-452.
Direct Link - Singh, R.K., R.C. Dhiman and P.K. Mittal, 2006. Mosquito larvicidal properties of Momordica charantia Linn (Family: Cucurbitaceae). J. Vect. Borne. Dis., 43: 88-91.
Direct Link - Tantawy, A.O., A.A. Abdel-Malek and A.M. Wakid, 1967. Studies on the eradication of Anopheles pharoensis by the sterile-male technique using cobalt-60. V. Mating competitiveness in radiosterilized males. J. Econ. Entomol., 60: 696-699.
Direct Link - Tikar, S.N., M.J. Mendki, K. Chandel, B.D. Parashar and Shri Prakash, 2008. Susceptibility of immature stages of Aedes (Stegomyia) aegypti; vector of dengue and chikungunya to insecticides from India. Parasitol. Res., 102: 907-913.
CrossRefDirect Link