
Addis Teshome
International Centre for Insect Physiology and Ecology, P.O. Box-30772-00100, Nairobi, Kenya
S. K. Raina
International Centre for Insect Physiology and Ecology, P.O. Box-30772-00100, Nairobi, Kenya
F. Vollrath
Department of Zoology, Oxford University, South Parks Road, Oxford OX1 3PS, United Kingdom
J. M. Kabaru
School of Biological Sciences, University of Nairobi, P.O. Box 30197-00100, Kenya
J. Onyari
School of Biological Sciences, University of Nairobi, P.O. Box 30197-00100, Kenya
E. K. Nguku
International Centre for Insect Physiology and Ecology, P.O. Box-30772-00100, Nairobi, Kenya
Journal of Entomology
Year: 2011 | Volume: 8 | Issue: 5 | Page No.: 450-458







ABSTRACT
A study was conducted to determine weight loss and moisture regain properties of silk cocoon shells, shell layers and degummed fibers of the four African wild silkmoths Anaphe panda, Argema mimosae, Epiphora bauhiniae and Gonometa postica and compare with the industry standard mulberry silkmoth, Bombyx mori. No. significant difference was observed between the moisture regain of the wild silk fibers after degumming. However, there were significant differences in weight loss and moisture regain between cocoon shells as well as shell layers. E. bauhiniae had the lowest weight loss and moisture regain of 23.19 and 5.64%, respectively while G. postica had the highest weight loss and moisture regain, 56.84 and 9.05%, respectively. The SEM micrographs also showed the presence of remnant sericin gum on the fibers surface. In both A. panda and E. bauhiniae, the outer shell layers had the lowest moisture regain and highest weight loss while the inner layer of E. bauhiniae and middle layer of A. panda lost the least weight. The physical structure and chemical composition of the cocoon shells and fibers contribute towards the variation in the moisture regain and weight loss and these features may have commercial implications due to their direct effect on the resulting fabric.
PDF Abstract XML References Citation
Received: February 17, 2011;
Accepted: April 12, 2011;
Published: May 30, 2011
How to cite this article
Addis Teshome, S. K. Raina, F. Vollrath, J. M. Kabaru, J. Onyari and E. K. Nguku, 2011. Study on Weight Loss and Moisture Regain of Silk Cocoon Shells and Degummed Fibers from African Wild Silkmoths. Journal of Entomology, 8: 450-458.
DOI: 10.3923/je.2011.450.458
URL: https://scialert.net/abstract/?doi=je.2011.450.458
DOI: 10.3923/je.2011.450.458
URL: https://scialert.net/abstract/?doi=je.2011.450.458
INTRODUCTION
Silks are natural polymers which have been gaining wide use in variety of applications such as degradable biomaterials, biomedical and functional bio-membrane materials and fibers besides their traditional use as a textile raw material (Zhang, 2002; Mondal et al., 2007a; Wilaiwan et al., 2010). The use of such silks, depending on their origin, can be justified both by the combination of mechanical properties, non-toxicity, biocompatibility and biodegradability depending on source and treatment. African wild silk fibers from Saturniidae (Ephiphora and Argema spp.), Lasiocampidae (Gonometa spp.) and Thaumetopoeidae (Anaphe, Epanaphe and Hyposoides spp.) genera produce silk of potential commercial value and with significant economic importance due to their interesting mechanical properties (Raina, 2004; Delport, 2006). However, the performances of all silk biopolymers are affected when exposed to adverse environments.
Specifically, absorbed moisture is likely to determine the mechanical and physical properties of both the sericin gum coating and the integral structural proteins of the silks (Fut et al., 2009). Some silks can indeed be hygroscopic with a moisture regain of 11 at 65% Relative Humidity (RH) and 27°C (Ranganathaian, 2003; Sonthisombat and Speakman, 2004) which is larger than many other commercial fibers and more in line with wool (Salamone, 1999). This interaction of moisture and fiber has technical as well as commercial consequences. For example, in Bombyx mori Linn. silk, the absorption of water causes 1.6% fiber swelling in the longitudinal crosswise (Schoeser, 2007). This apparently small degree of swelling can influence the rate of heat and moisture vapour transfer through the textile fiber. Moreover, the weight change resulting from moisture absorption can affect the commercial weight of the fibers as well as affecting properties such as dying, finishing and apparent comfort of the product (Salamone, 1999).
However, most silk contains not only the fiber matrix protein but in addition have a coating of sericin or similar compounds. This sericin component of the composite silk fibre (typically of two fibre brins held together into the bave by the sericinaceous gum), is usually expressed as weight loss and is an important additional factor affecting the quality of a silk (Vishuprasad, 2004). The sericin content of a fiber is estimated by boiling silk (using a range of chemicals) and calculating the weight loss (Sonthisombat and Speakman, 2004). This weight loss from a raw silk filament depends on the water solubility and higher alkali sensitivity of the sericin as compared to the fibroin (Choundhury, 2006). Quantity and nature of sericin are fundamental characteristics in conferring distinctive traits to the cocoon. Sericin coating has important biological properties such as oxidation and UV resistance as well as antibacterial and moisture control properties (Mondal et al., 2007a). However, the sericin coating also gives a callous and stiff feeling to the fiber and hides the rich luster and whiteness of the silk brins (Arami et al., 2007) in addition to preventing the penetration of dye. These necessitate its removal by process of boiling to obtain an ideal fiber for the textile industry and any other specific applications. Because of these effects, understanding the amount of sericin present in silk bave as wells as their water absorption properties of fibers has considerable technical implications for a silk as a commercial proposition. Hence, this study was conducted with the objective to determine the quantity of sericin gum and moisture regain properties in African wild silk cocoon shells and fibers.
MATERIALS AND METHODS
Weight loss: This study was conducted between June to August, 2010 at the International Centre of Insect Physiology and Ecology, Commercial Insects Program Laboratory. Cocoon shells of four wild silk species were collected from four rather different ecotypes in Kenya namely; the Kakamega (Anaphe panda Biosduval), the Arabuko Sokoke (Argema. Mimosa Biosduval), the West Pokot (E. bauhiniae Guer) and Mwingi district acacia bush lands (G. postica Walker) (Fig. 1). Cocoons of B. mori (Race ICIPE II) were obtained for comparison from the Commercial Insect Program of icipe, International Centre for Insect Physiology and Ecology. Cocoons were cut with a surgical blade and cleaned by removing the pupal debris and other foreign materials such as leaves and branches. Twenty grams of cleaned cocoons were enclosed in wire mesh cages with a volume of 717 cm3 and boiled with 5 g L-1 of Na2CO3 (Magadi soda) solution for 180 min. Boiled cocoons were soaked in star soft solution of 50 mL L-1 of distilled water for 3 min and washed with hot and cold distilled water twice. Excess water was removed by filter paper and ccocoon filaments were allowed to spend 24 h in oven at 110°C to insure uniform drying. The weight loss, expressed in percentage, was obtained using the equation (Nakpathom et al., 2009).
Moisture regain of cocoon shells: Moisture regain of undegummed but clean and weighed cocoon shells was determined by the oven drying procedure. Known weight of cocoon shells were placed in an oven at 110°C for 24 h.
 | |
| Fig. 1: | Degummed fibers of (a) Argema mimosae, (b) Anaphe panda and (c) Gonometa postica showing the remnant gum |
Twenty grams of oven dried shells were placed at laboratory bench for 72 h at room conditions (23±2°C and 71±3% RH). The moisture regain percentage for the cocoon shells was obtained by Nawaz et al. (2002).
To avoid initial weight gain during the weighing process care was taken not to expose the samples to moisture.
Moisture regain of degummed fibers: Cleaned cocoon shells were first dugmmed and then dried in the oven for 24 h at 110°C. Twenty gram of degummd fibers were left on laboratory bench under room conditions (23±2°C and 71±3% RH) for 72 h. The moisture regain percentage for the fibers were obtained according to Nawaz et al. (2002).
Weight loss and moisture regain of cocoon shell layers: For this study, only cocoon shells of A. panda and E. bauhiniae were used. These species had been selected because of the clear distinction in the physical structure of the three layers; a leathery outer envelope, a fluffy intermediate layer and thick walled and rigid inner layer in their cocoon shells. Three distinct layers designated as outer, middle and inner were separated. Weight loss and moisture regain were determined by the same procedure as described above.
Scanning Electron Microscope (SEM) observation: Degummed fibre samples were embedded in black wool and mounted onto copper stubs using double side sticking tape. The mounted specimens were immediately coated with gold (JEOL JFC-1100E ion sputter) and observed with SEM (JEOL-JSM T330A) under an accelerating voltage of 10 kV.
Data analysis: All percentage data were logarithmic transformed to stabilize the variance before the final statistical analysis. Both moisture regain and weight loss of the cocoon shells were analyzed using one-way Analysis of Variance (ANOVA) (p<0.01) while two-way analysis was employed to analyze the difference in cocoon layers of A. panda and E. bauhiniae with PROC ANOVA (SAS Institute, 2010). Least Significance Difference (LSD) test was used to separate means.
RESULTS
Weight loss and moisture regain of cocoon shells: The effect of boiling cocoons shells of four African wild and B. mori silk on weight loss and moisture regain is presented in Table 1. There was a highly significant difference in weight loss between the cocoon shells. G. postica and E. bauhiniae cocoons had the highest and lowest weight loss (57 and 23%, respectively). B. mori and A. mimosae cocoons had comparable weight losses (29 and 31%) which were marginally higher than A. panda (26%). The SEM micrographs revealed that the fibers were covered with different quantities of sericin gum after the degumming treatment (Fig. 1). There were also significant differences in the moisture regain percentage between cocoon shells and degummed fibers. A. mimosae cocoons had the highest regain (9%) while E. bauhiniae cocoons gained the lowest moisture (5.64%). B. mori and A. panda cocoons gained marginally (statistically insignificant) more moisture (7.93 and 8.02%, respectively) than G. postica cocoon shells (7.11%) (Table 1). In contrast to the cocoon shells, the moisture regain of degummed wild silk fibers was not significantly different between species with E. bauhiniae, G. postica and A. panda gaining 10, 9.9 and 9.9%, respectively. However, B. mori had the lowest value (8.53%) which was significantly different from the other wild fibers (Table 1).
Moisture regain and weight loss of cocoon shell layers: Moisture regain of layers of E. bauhiniae and A. panda cocoon shells is shown in Table 2. There was significant difference in moisture regain of cocoon shell layers of E. bauhiniae and A. panda. The inner hard layers of both species have the highest regain (6.8 and 8.6% for E. bauhiniae and A.panda, respectively).
| Table 1: | Mean±SE of weight loss and moisture regain of African wild cocoons and degummed fibers |
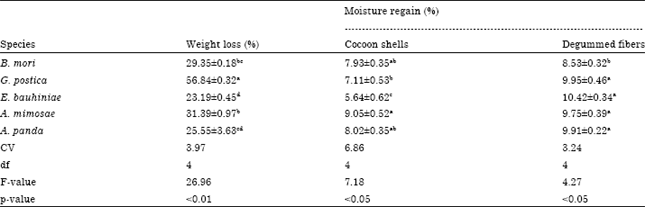 | |
| *Means followed by the same letter in a column are not statistically significant (p<0.05) using the least significant difference test (LSD) | |
| Table 2: | Mean±SE of moisture regain of layers of E. bauhiniae and A. panda cocoon shells |
 | |
| *Means followed by the same letter in a column are not statistically significant (p<0.05) using the least significant difference test (LSD) | |
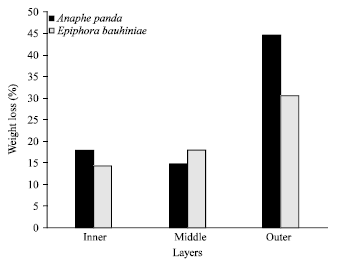 | |
| Fig. 2: | Weight loss of the three layers of A. panda and E. bauhiniae cocoon shells |
However, there was no significant difference between the regain percentages of degummed shell layer fibers of E. bauhiniae while A. panda degummed layer fibers showed significant difference with the inner layer gaining more moisture (9.4%). The moisture regain for A. panda shell layers progressively increased towards the inner layer. There was highly significant difference in weight loss in both species. The outer skin like layers had the highest weight losses of 30.5 and 44.6% for E. bauhiniae and A.panda, respectively and the weight loss significantly decreased to the inner layer in E. bauhiniae (Fig. 2).
DISCUSSION
The result clearly demonstrated that weight loss and moisture regain varies among the African wild silk cocoon shells and degummed fibers. The weight loss and moisture regain for B. mori was well within the limits of other studies, 19 to 30% (Arami et al., 2007; Gulrajani, 1988; Sarovart et al., 2003) and 6.7 to 12% (Maji et al., 2002), respectively. In this study, the weight loss of wild silk fibers ranged from 23 to 56.8% and is inconsistent with previous reports that the sericin content of wild silk fibers is less than the mulberry silk and is as low as 12-16% (Cavaco-paulo and Gubitz, 2003; Prasong et al., 2009). Karumar (1999) reported 5-15% sericin gum coating content for tussah silk fibers. The degumming loss in Antheraea proylei, A. assama, A. pernyi and A. yamamai was in the range of 7-13% except for A. mylitta and Philosamia Cynthia ricicni which showed a 3.7 and 4.5% loss, respectively (Kato, 1991). However, higher sericin (gum) content of 38, 45 and 67.9% have been reported for Thai silk, Rondotia menciana and Bombyx mandarina cocoon shells, respectively (Dhavalikar, 1962; Ito et al., 1992). The food plants and the environment might have contributed to the higher quantity and differential solubility of sericin gum in African wild silk cocoon shells. Despite these higher values, the weight losses reported in this study for the wild silk cocoons might not be the true reflection of the gum content of the cocoons as verified by the intact inner hard parchment like layers for A. panda and E. bauhiniae and traces of sericin gum observed on the SEM images after degumming.
The variations in the amount of gum cementing substance (sericin) and fibroin in the cocoons shells might be due to a number of factors including taxonomic (genetic) difference in the sericin protein as well as external environmental variables (Shamitha and Purushotham, 2006) which together can determine the quality of cocoons for commercial purposes (Singhvi and Bose, 1991 cited in Shamitha and Purushotham, 2006). According to Kato and Hata (1998), the cocoon filaments of polyphagous silkworms with coarse filaments have higher values for the degumming loss than conventional cocoon filaments. This can be an explanation for the high weight loss of G. postica which is a polyphagous silkworm feeding on more than one acacia host plants including Acacia tortilis, A. elatior, A. nilotica, A. mellifera, A. mearnsii, A. hockii and A. brevispica (Fening et al., 2008; Ngoka et al., 2007). Nasreen et al. (1999) also reported difference in quality of B. mori cocoons from larvae fed on mixed leaves. Chemical composition of sericin also affects its solubility in hot water resulting in considerable variation in weight loss (Komatsu, 1975; Robson, 1985). The presence of certain amount of sericin after degumming shows its important role in the processing and application of the fibers.
Physical properties and chemical composition of silk fibers from mulberry and wild silkmoths exhibit variability in different parameters (Mondal et al., 2007b; Srisuwan and Srihanam, 2009). Degummed wild silk fibers have higher regains than those of commercial mulberry silk fibers. Sen and Babu (2004) also observed similar results with the non-mulberry silk fibers having marginally higher moisture regain values of 10.76%. This is in contrast with Snowalker (1993) who reported degummed fibers to have lower moisture regain due to the loss of sericin. The difference in moisture regain might be due to variations in compactness of fibers, change in the quantity, composition and chemistry of the gum and surface structure of the shells. The moisture regain of cocoon shells and fibers could provide information on the extent of areas accessible to water vapour within the fibre. The higher moisture regain for A. mimosae cocoon shells might be due to the presence of holes/perforations in the cocoon surface providing more area for moisture ingress. The presence of higher ratios of hydrophilic versus hydrophobic amino acid residues in the chemical architecture of non mulberry compared to mulberry silks may also affect in the higher moisture regain of the wild silks.
The study showed decreased weight loss and increased moisture regain from outer to inner shell layers. Nawaz et al. (2002) revealed with the change in filament type and size (denier), moisture regain changes significantly due to the difference in structures of fibres as well as changes in liner density and the arrangement of monomers in the polymer (Munro, 1987). The presence of foreign matters such as dust and the hair like brittles on the outer layer of A. panda suggested the influence of physical structure in preventing fibers from absorbing moisture. The variation in weight loss and moisture regain among the different shell layers can be the demonstration of the difference in the amount and composition of sericin present in each layer. Such layer wise variation in cocoon property down the cocoon layers may confound objectionable fabric patterns either from visual or textural perspective (Das and Ghosh, 2010).
In conclusion, with other important qualities determined for some of the African wild silk cocoons and fibers, understanding the moisture regain and sericin content will have a significant importance in establishing quality parameters of the different fibers. The observed variation in the properties of cocoon layers suggests the need for separate boiling of layers in contrast to the usual practise of mass boiling if that were practically possible. Finally, present result suggests the need to selection bespoke degumming techniques for each type of silk in order to optimize the quality and quantity of the fibers that can be drawn from the different cocoon shells.
ACKNOWLEDGMENTS
Thanks are due to ICIPE-African Insect Science for Food and Health for providing logistical and research support. We are indebted to Rolf Gloor, Cabesi project leader, for providing E. bauhiniae cocoons. We also highly appreciate the technical support of other staff of the Commercial Insect Program.
REFERENCES
- Arami, M., S. Rahimi, L. Mivehie, F. Mazaheri and N.M. Mahmoodi, 2007. Degumming of Persian silk with mixed proteolytic enzymes. J. Applied Polymer Sci., 106: 267-275.
CrossRef - Das, S. and A. Ghosh, 2010. An investigation on the progressive change in filament characteristics from outer to inner layers of mulberry and tasar cocoons. J. Applied Polymer Sci., 117: 1319-1324.
CrossRef - Fening, K.O., E.N. Kioko, S.K. Raina and J.M. Mueke, 2008. Monitoring wild silkmoth, Gonometa postica Walker, abundance, host plant diversity and distribution in Imba and Mumoni woodlands in Mwingi, Kenya. Int. J. Biodiver. Sci. Ecosyst. Serv. Manage., 4: 104-111.
CrossRefDirect Link - Fut, C., D. Porter and Z. Shao, 2009. Moisture effects on Antheraea pernyi silk's mechanical property. Macromolecules, 42: 7877-7880.
CrossRef - Ito, T., J. Xu, K. Hamano, T. Inokuchi and M. Yamada, 1992. Amino acid composition of cocoon shell of the mulberry white caterpillar, Rondotia menciana and the wild silk worm, Bombyx mandarina. J. Seric. Sci. Jap., 61: 105-109.
Direct Link - Kato, H. and T. Hata, 1998. Degumming and dyeing properties of cocoon filaments with a special feature differed from conventional silkworm races. Bull. Natl. Inst. Seric. Entomol. Sci., 20: 101-114.
Direct Link - Maji, T.K., D. Basu, C. Datta and A. Banerjee, 2002. Studies of mechanical and moisture regain properties of methyl methacrylate grafted silk fibers. J. Applied Polym. Sci., 84: 969-974.
CrossRef - Mondal, M., K. Trivedy and S.N. Kumar, 2007. The silk proteins, sericin and fibroin in silkworm, Bombyx mori Linn., a review. Caspian J. Enviorn. Sci., 5: 63-76.
Direct Link - Mondal, M., K. Trivedy, S.N. Kumar and V. Kumar, 2007. Scanning electron microscopic study on the cross sections of cocoon filament and degummed fiber of different breeds/hybrids of mulberry silkworm, Bombyx mori Linn. J. Entomol., 4: 362-370.
CrossRefDirect Link - Nakpathom, M., B. Somboon and N. Narumol, 2009. Papain enzymatic degumming of thai Bombyx mori silk fibers. J. Micr. Soc. Thailand, 23: 142-146.
Direct Link - Nasreen, A., G.M. Cheema and M. Ashfaq, 1999. Rearing of silkworm Bombyx mori L. on alternate food plants. Pak. J. Biol. Sci., 2: 843-845.
CrossRefDirect Link - Nawaz, S.M., N.A. Jamil, A. Farooq and T. Mahmood, 2002. Comparative study of shrinkage and moisture regain of silk, viscose rayon and polyester filament yarn. Int. J. Agric. Biol., 4: 493-495.
Direct Link - Ngoka, B.M., E.N. Kioko, S.K. Raina, J.M. Mueke and D.M. Kimbu, 2007. Semi-captive rearing of the African wild silkmoth Gonometa postica (Lepidoptera: Lasiocampidae) on an indigenous and a non-indigenous host plant in Kenya. Int. J. Trop. Insect Sci., 27: 183-190.
CrossRef - Prasong, S., S. Yaowalak and S. Wilaiwan, 2009. Characteristics of silk fiber with and without sericin component: A comparison between Bombyx mori and Philosamia ricini silks. Pak. J. Biol. Sci., 12: 872-876.
CrossRefPubMedDirect Link - Ranganathaian, C., 2003. Positron Lifetime study of thermally induced microstructural changes in nastari silk fibers. Chinese J. Polymer Sci., 21: 325-332.
Direct Link - Sarovart, S., B. Sudatis, P. Meesilpa, B.P. Grady and R. Magaraphan, 2003. The use of sericin as an antioxidant and antimicrobial for polluted air treatment. Rev. Adv. Mater. Sci., 5: 193-198.
Direct Link - Sen, K. and M.K. Babu, 2004. Studies on Indian silk. I. Macrocharacterization and analysis of amino acid composition. J. Applied Polymer Sci., 92: 1080-1097.
CrossRef - Shamitha, G. and R.A. Purushotham, 2006. Studies on the filament of tasar silkworm, Antheraea mylitta D (Andhra local ecorace). Curr. Sci., 90: 1667-1671.
Direct Link - Srisuwan, Y. and P. Srihanam, 2009. Dissolution of Philosamia ricini silk film: Properties and functions in different solutions. J. Applied Sci., 9: 978-982.
CrossRefDirect Link - Vishuprasad, N., 2004. Composite quality index of silk cocoons-application of discriminant analysis. J. Acad. Bus. Econ., Vol. 4, No. 1.
Direct Link - Wilaiwan, S., S. Yaowalak, B. Yodthong and S. Prasong, 2010. Silk fibroin/gelatin hybrid films for medical applications: Study on chlorhexidine diacetate. J. Biol. Sci., 10: 455-459.
CrossRefDirect Link - Zhang, Y.Q., 2002. Application of natural silk protein sericin in biomaterials. Biotechnol. Adv., 20: 91-100.
PubMed