
Adaku A. Lawrence
Department of Life Sciences, University of the West Indies, St. Augustine Trinidad, West Indies
Ayub Khan
Department of Life Sciences, University of the West Indies, St. Augustine Trinidad, West Indies
Journal of Entomology
Year: 2009 | Volume: 6 | Issue: 2 | Page No.: 102-108







ABSTRACT
This study aims to determine the effect of temperature on conidial germination, colony radial growth and virulence of two isolates of Beauvaria bassiana (ARSEF-1186 and IMI-351833) to the cowpea weevil, Callosobruchus maculatus under controlled laboratory conditions. Isolates grown on Potato Dextrose Agar plates indicated that the rate of conidial germination was slower at 20 °C than at 25 and 30 °C and was expressed by the following 50% Germination Time (GT50) values: 17.23 ± 1.02 h for ARSEF-1186 and 16.46 ± 1.02 h for IMI-351833. The lowest GT50 value was estimated as 12.33 ± 1.02 h for IMI-351833 at 25 °C. ARSEF-1186 showed significantly faster colony radial growth rate (Kr) than IMI-351833 for all temperatures investigated. Bioassays using Cajanus cajan seeds dipped in conidial suspensions of both isolates indicated that the highest virulence gainst C. maculatus adults was observed with ARSEF-1186 at 30 °C. This isolate exhibited both the lowest LC50 (2.29x105 spores mL-1) and LT50 (4.17 days) values.
PDF Abstract XML References Citation
How to cite this article
Adaku A. Lawrence and Ayub Khan, 2009. Variation in Germination and Growth Rates of Two Isolates of Beauvaria bassiana (Balsamo) Vuillemin (Deuteromycota: Hyphomycetes at Different Temperatures and their Virulence to Callosobruchus maculatus (Fabricius) (Coleoptera: Bruchidae). Journal of Entomology, 6: 102-108.
DOI: 10.3923/je.2009.102.108
URL: https://scialert.net/abstract/?doi=je.2009.102.108
DOI: 10.3923/je.2009.102.108
URL: https://scialert.net/abstract/?doi=je.2009.102.108
INTRODUCTION
Grain legumes serve as the primary protein source in many parts of the Caribbean, India and Africa, due to their relatively cheap and easy production and rapid proliferation in tropical climates (Labeyrie, 1981; Jeong et al., 2006). These grain legumes suffer severe quantitative and qualitative losses from attack by various species of insects, 75% of which are coleopterans (Labeyrie, 1981; Jeong et al., 2006). The most damaging bruchid pest is the cowpea seed beetle, Callosobruchus maculatus (Fabricius) (Coleoptera:Bruchidae) (Jackai and Daoust, 1986; Ogunwolu and Odunlami, 1996; Jha et al., 2008). Substantial losses of revenue occur in storage, especially in rural areas of developing countries, where grain legumes are kept in mediocre conditions. Caswell (1981) reported a loss of approximately 50% of grain legumes in storage for three to four months due to infestation by C. maculatus.
The control of arthropod pests on stored products has been primarily through the use of fumigants and residual insecticides to augment the more obvious approach of hygiene (Brooker et al., 1992; Adane et al., 1996; McGraw, 1998). Many insects are capable of tolerating virtually all pesticides available for their control as a result of cross- and multiple-resistance (Metcalf, 1980; Khan and Selman, 1989). Coupled with the recognition of the deleterious effects of conventional insecticides is the burgeoning consumer preference for foods grown and stored organically. To avoid the use of pesticides as well as the stricter requirements of pesticide registration, consumers have collectively spurred the war against these traditional pesticides. The use of entomogenous fungi to control storage pests offers the opportunity of using living organisms that are natural enemies of the target pest species to maintain pest populations well below the economic threshold. Mycopesticides have been shown to have considerable potential for the management of insects and they have been used worldwide to control various coleopteran pests (Gottwald and Tedders, 1983; Khan and Selman, 1988; Rodriguez and Pratissolli, 1990; Adane et al., 1996; Padín et al., 1997). One of the principal factors limiting the efficacy of entomopathogenic fungal control is temperature (Carruthers et al., 1985; Benz, 1987; Fernandes et al., 2008). Given that a window of opportunity often exists for the pathogen to infect the host, the influence of temperature on the infection process is important. In the present study, the effect of different temperatures on conidial germination, radial colony growth and virulence to C. maculatus of two isolates of B. bassiana were investigated. Both isolates of B. bassiana were originally collected from Coleopteran pests that usually thrive in conditions similar to those of storage bruchids. ARSEF-1186 obtained from the United States Department of Agriculture (USDA) was isolated from infected Caryedon serratus (Olivier) (Coleoptera: Bruchidae). The second isolate, IMI-351833 was obtained from the CAB International Mycological Institute and was isolated from Sitone discoides (Gyellenhal) (Coleoptera: Curculionidae).
MATERIALS AND METHODS
Insect Cultures
Laboratory cultures of C. maculatus were established from unselected strains of field-collected adults infesting C. cajan in East and Central Trinidad in January 2006. Adults were introduced onto dried C. cajan seeds in clean glass jars (16x16x20 cm) and covered with a fine mesh cloth (250 mesh cm-2). Females were allowed to oviposit freely for 24 h and then removed. The egg covered seeds were left undisturbed in their jars at 27.0 ± 2.0 °C. Emerging adults were removed from the jars and allowed to mature for two weeks in order to allow sufficient development of reproductive organs. These mature adults were subsequently used as test subjects in bioassays.
Fungal Cultures
B. bassiana isolates (ARSEF 1186 and IMI 351833) were passaged through C. maculatus adults by allowing insects to walk onto 2 week old fungal cultures in Petri plates and then removed, placed onto moistened filter paper inside a sterile Petri plate and left until there was complete mortality. The resulting moribund cadavers were removed from the dishes and the surface spores used to inoculate new Petri plates of Potato Dextrose Agar (PDA) which were allowed to germinate at 25 °C for three weeks. Conidia were harvested by gently scraping the surface of the profusely sporulated fungal mats with a sterile scalpel and suspended in 50 mL sterile water with 0.5 ml L-1 Tween 80®. The resulting suspension was placed on a magnetic stirrer for 15 min at room temperature to ensure that conidia were homogenously distributed then filtered through sterile muslin to remove mycelial debris. Conidial concentration was estimated using an improved Neubauer Bright Line® haemocytometer under a light microscope. Conidial suspensions were diluted to 1x109 spores mL-1 and refrigerated at 4 °C overnight.
Effect of Temperature on in vitro Growth of Conidia
Two stock conidial suspensions were diluted to 1x107 spores mL-1 using 0.5 ml L-1 Tween 80®. For each isolate, 108x90 mm Petri plates each containing 10 mL of PDA were inoculated at the centre of each plate with 3 μL of conidial suspension. Thirty six inoculated plates of each fungal strain were incubated at each of three different temperatures (20, 25 and 30 °C). Four plates of each fungal strain at each temperature were removed at 2 h intervals over a 24 h period and spore germination arrested by the introduction of a few drops of cotton blue in lactic acid. For each replicate 200 conidia were assessed microscopically for germination, a conidium being considered to have germinated if it had a germ tube at least as long as the diameter of the average conidium (Yeo et al., 2003).
Effect of Temperature on in vitro Colony Radial Growth
A few drops of the stock conidial suspension of each fungal strain were placed onto PDA plates and incubated at 25 °C for three days to obtain mycelial mats. For each of the two strains, 5 mm diameter plugs were cut from the non-sporulating mycelia using a cork borer and a single plug was inverted in the centre of the new 90 mm Petri plate containing 25 mL of PDA. The circumference of the plates were sealed with Parafilm® and incubated in the dark in separate incubators at 20, 25 and 30 °C. Five replicates were prepared for each strain-temperature combination. Surface radial growth was recorded daily using two cardinal diameters previously drawn on the bottom of the Petri plate. The experiment was conducted for a duration of 30 days or until the fungal colony had covered the entire dish surface.
Effect of Temperature on Virulence of Fungi to Adult Callosobruchus maculatus
Serial dilutions (109, 108, 107, 106, 105 spores mL-1) for each fungal stock suspension were prepared with diluted Tween 80® (0.5 ml L-1) producing five conidial concentrations for each fungal isolate. Ten dried seeds of C. cajan were placed sterile muslin bag and dipped for 15 sec in the selected suspension while on a magnetic stirrer. The seeds were placed on clean filter paper and air dried overnight. These seeds were placed in clean 90 mm Petri plates to which 10 adult C. maculatus were introduced and the plate was covered with a mesh-covered lid and sealed around the circumference with Parafilm® and incubated at each of three temperatures (20, 25 and 30 °C). There were five replicates for each conidial suspension-fungal strain-temperature combination. Controls were treated with diluted Tween 80® in the same manner as those inoculated with conidia. Mortality was recorded daily commencing 24 h after inoculation and continued for 14 days. Insect cadavers resulting from the bioassays were placed on moist filter paper in clean 90 mm Petri plates and observed for the appearance of fungal growth over a 5 days period.
Data Analysis
For each replicate at each temperature-isolate combination, the percentage conidial germination over time was analyzed using PROBIT procedure (EPA ver. 1.4) to calculate the time taken in hours for 50% of the conidia to germinate (GT50). Comparisons of different fungal treatment-temperature combinations were made by examination of the respective probit regression lines of the GT50 values. A difference between two intervals was considered to be significant (p<0.05) if the 95% confidence intervals did not overlap (Mason and Johnson, 1987). A linear relationship between colony radius and time was assumed based on earlier studies demonstrated by other authors (Pirt, 1967; Yeo et al., 2003). For each replicate of each isolate- temperature combination, the colony radial growth rate, Kr (μm h-1) was estimated from the slope of the linear regression of the colony radius on time. Parallel model regression analysis of the Kr estimates against temperature were used to compare the two isolates. The Lethal Concentration causing 50% mortality (LC50) and Lethal Time for 50% mortality (LT50) were analyzed using PROBIT (EPA ver. 1.4). Comparisons of different fungal treatment-temperature combinations were made by examination of the respective probit regression lines of the LC50 and LT50 values, similar to those made for the GT50.
RESULTS AND DISCUSSION
B. bassiana is a cosmopolitan facultative entomopathogenic Hypomycete capable of growth within all temperature regimes investigated; however, there was a wide variation in response between isolates. There was generally a good correlation between temperature effects on germination and vegetative colony growth in both isolates (Fig. 1, 2). Conidial germination rate as measured by GT50 for both isolates were significantly faster (p<0.05) at 25 and 30 °C than at 20 °C. ARSEF-1186 exhibited no significant difference (p>0.05) in growth rate at 25 and 30 °C, while IMI-351833 grew significantly faster (p<0.05) at 25 °C than either 20 °C or 30 °C (Table 1). The optimal temperature for conidial germination of IMI-351833 was 25 °C whereas that for ARSEF-1186 was 30 °C although this was not significantly different from 25 °C (Table 1). These findings are in accordance with those obtained in other studies of entomopathogenic Hypomycetes (Fargues et al., 1997; Ekesi et al., 1999; Aiuchi et al., 2008; Fernandes et al., 2008).
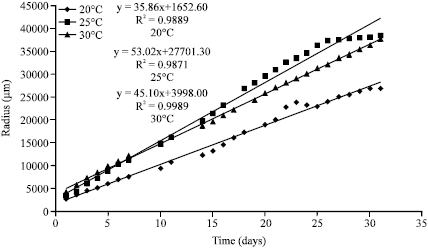 | |
| Fig. 1: | Radial colony growth of Beauvaria bassiana (ARSEF-1186) at three temperatures |
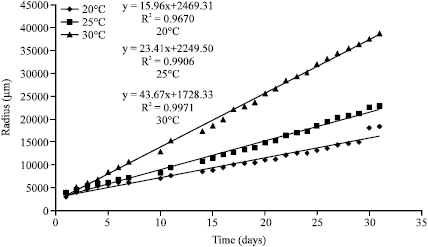 | |
| Fig. 2: | Radial colony growth of Beauvaria bassiana (IMI-351833) at three temperatures |
| Table 1: | GT50 values for Beauvaria bassiana strains at three temperatures |
 | |
| *Means followed by the same letter(s) in a column are not significantly different from each other (p>0.05) | |
The relationship between Kr values and temperature was expressed as a linear model since the Kr values for both isolates showed a general increase in growth rate with increasing temperature (Fig. 3).
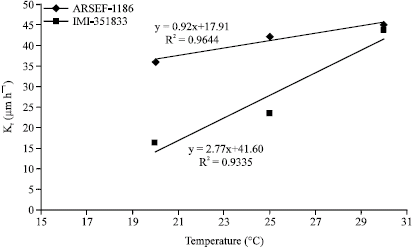 | |
| Fig. 3: | Comparison of Kr values for isolates of Beauvaria bassiana at three temperatures |
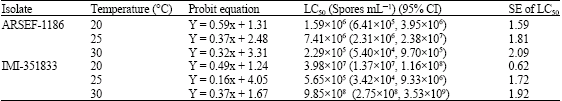
| Table 2: | LC50 values for adult Callosobruchus maculatus exposed to Cajanus cajan seeds treated with different strains of Beauvaria bassiana at three temperatures |
 | |
| Table 3: | LT50 values for adult Callosobruchus maculatus exposed to Cajanus cajan seeds treated with different strains of Beauvaria bassiana at three temperatures |
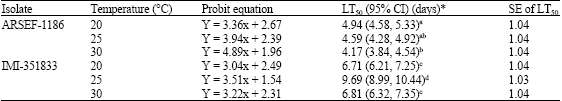 | |
| *Means followed by the same letter(s) in a column are not significantly different from each other (p>0.05) | |
According to Fargues et al. (1997) there is significant difference in variability of growth at different temperatures among isolates of B. bassiana. Selection according to thermal requirements may therefore be strongly warranted when choosing an isolate for use as a microbial control agent against C. maculatus. As with other stored product insects, development and survival as well as oviposition of C. maculatus is influenced to a large extent by environmental conditions including temperature and humidity (Iloba and Osuji, 1996; Lale and Vidal, 2001). Both factors have been shown to set the lower and upper limits for oviposition and progeny development in pure populations of C. maculatus. It was observed that seeds treated with IMI-351833 conidia bore fewer eggs than those inoculated with ARSEF-1186. Additionally, there were no second generation adult emergences for all treatments with IMI-351833, whereas adult emergence was not affected by treatments with ARSEF-1186. However, sufficient data was not collected in order to conduct any acceptable statistical analysis on this aspect of the study.
ARSEF-1186 showed higher virulence to C. maculatus when compared to IMI-351833 as evident by the lowest LC50 (2.29x105 spores mL-1) and LT50 (4.17 ± 1.04 days) values both occurring at 30 °C (Table 2, 3) and this was corroborated by a faster germination time and rate of colony growth. ARSEF-1186 thus provides a combination of low lethal concentration and rapid mortality.
Due to the widespread distribution of C. maculatus in the tropics and subtropics, it is assumed that the pest can tolerate temperature fluctuations in the warmer regions. With this underlying premise, microbial control agents that thrive within a relatively wide range of thermal conditions should be chosen. Unlike field constraints to microbial control however, the stored product setting is almost devoid of detrimental, unpredictable environmental factors including extreme temperature fluctuations, thus affording a more stable milieu for biological control using entomopathogenic fungi (Burkholder and Faustini, 1991; Lawrence and Khan, 2002).
REFERENCES
- Adane, K., D. Moore and S.A. Archer, 1996. Preliminary studies on the use of Beauvaria bassiana to control Sitophilus zeamais (Coleoptera: Curculionidae) in the laboratory. J. Stored Prod. Res., 32: 105-113.
Direct Link - Aiuchi, D., Y. Baba, K. Inami, R. Shinya, M. Tani and M. Koike, 2008. Variation in growth at different temperatures and production and size of conidia in hybrid strains of Verticillium lecanii (Lecanicillium sp.) (Deuteromycotina: Hypomycetes). Applied Entomol. Zool., 43: 427-436.
CrossRefDirect Link - Carruthers, R.I., Z. Feng, D.S. Robson and D.W. Roberts, 1985. In vitro temperature dependent development of Beauvaria bassiana (Deuteromycotina: Hypomycetes) mycosis of the European corn borer, Ostrina nubilalis (Lepidoptera: Pyralidae). J. Invertebr. Pathol., 46: 305-311.
CrossRefDirect Link - Caswell, G.H., 1981. Damage to stored cowpeas in Northern Nigeria. Samaru J. Agric., 1: 11-19.
Direct Link - Ekesi, S., N.K. Maniania and K. Ampong-Nyarko, 1999. Effect of temperature on germination, radial growth and virulence of Metarhizium anisopliae and Beauvaria bassiana on Megalurothrips sjostedti. Biocontrol. Sci. Technol., 9: 177-185.
CrossRefDirect Link - Fargues, J., A. Ouedraogo, M.S. Goettel and C.J. Lomer, 1997. Effects of temperature, humidity and inoculation method on susceptibility of Schistocera gregaria to Metarhizium flavoviridae. Biocontrol Sci. Technol., 7: 345-356.
CrossRefDirect Link - Fernandes, E.K.K., D.E.N. Rangel, A.M.L. Moraes, V.R.E.P. Bittencourt and D.W. Roberts, 2008. Cold activity of Beauvaria and Metarhizium and thermotolerance of Beauvaria. J. Invertebr. Pathol., 98: 69-78.
CrossRefDirect Link - Gottwald, T.R. and W.L. Tedders, 1983. Suppression of pecan weevil (Coleoptera: Curculionidae) populations with entomopathogenic fungi. Environ. Entomol., 12: 471-474.
Direct Link - Iloba, B.N. and F.N.C. Osuji, 1996. The thermal death point of Callosobruchus Maculatus (Fabricius). Nigerian J. Entomol., 7: 18-23.
Direct Link - Jackai, L.E.N. and R.A. Daoust, 1986. Insect pests of cowpeas. Annu. Rev. Entomol., 31: 95-119.
CrossRefDirect Link - Jeong, H.J., J.E. Oh, S.W. Hong, M.I. Ryoo, K.D. Kim and H. Lee, 2006. Molecular marker-assisted detection of rice weevil in stored rice. Int. J. Pest Manage., 52: 325-331.
CrossRefDirect Link - Jha, A.N., C. Srivastava and S.K. Misra, 2008. Weight loss in cowpea cultivars to Callosobruchus maculatus in storage conditions. Ind. J. Entomol., 70: 181-183.
Direct Link - Khan, A.R. and B.J. Selman, 1988. Effect of insecticide, microsporidian and insecticide-microsporidian doses on the growth of Tribolium castaneum larvae. J. Invertebr. Pathol., 44: 230-232.
CrossRefPubMedDirect Link - Khan, A.R. and B.J. Selman, 1989. Nosema spp. (Microspora: Nosematidae) of stored product Coleoptera and their potential as microbial control agents. Agric. Zool. Rev., 30: 193-223.
Direct Link - Lale, N.E.S. and S. Vidal, 2001. Mortality of the different developmental stages of Callosobruchus maculatus (F.) and C. subinnotatus Pic. (Coleoptera: Bruchidae) in bambara groundnut, Vigna subterraneanm (L.) Verdc. seeds exposed to simulated solar heat. J. Plant Dis. Protect., 107: 553-559.
Direct Link - Lawrence, A.A. and A. Khan, 2002. Comparison of the pathogenicity of the entomopathogenic fungi, Beauvaria bassiana, Metarhizium anisopliae and Paecilomyces fumoso-roseus to Callosobruchus maculatus. Int. Pest Contr., 44: 125-127.
Direct Link - Mason, G.A. and M.W. Johnson, 1987. Assessment of insecticide susceptibility of Liriomyza sativae (Diptera: Agromyzidae) to topical and residual bioassays. J. Econ. Entomol., 80: 1083-1086.
Direct Link - Metcalf, R.L., 1980. Changing role of insecticides in crop protection. Ann. Rev. Entomol., 25: 219-256.
CrossRefDirect Link - Ogunwolu, E.O. and A.T. Odunlami, 1996. Suppression of seed bruchid (Callosobruchus maculatus) development and damage on cowpea (Vigna unguiculata) with Zanthoxylum zanthoxyloides (Lam.) Waterm. (Rutaceae) root bark when compared to neem seed powder and pirimiphos-methyl. Crop Prot., 15: 603-607.
CrossRefDirect Link - Padín, S.B., G.M. Dal Bello and A.L. Vasicek, 1997. Pathogenicity for adults of Tribolium castaneumm (Coleoptera: Tenebrionidae) in stored grains. Entomophaga, 42: 569-574.
CrossRefDirect Link - Pirt, S.J., 1967. A kinetic study of the mode of growth of surface colonies of bacteria and fungi. J. Gen. Microbiol., 47: 181-197.
PubMedDirect Link - Rodriguez, C. and D. Pratissoli, 1990. Pathogenicity of Beauvaria brongniartii(Sacc.) Petch. and Metarhizium anisopliae Mols. Sorokin and its effect on corn weevil and bean beetles. Ann. Entomol. Soc. Brazil, 19: 302-306.
Direct Link - Yeo, H., J.K. Pell, P.G. Alderson, S.J. Clark and B.J. Pye, 2003. Laboratory evaluation of temperature effects on the germination and growth of entomopathogenic fungi and their pathogenicity to two aphis species. Pest Manage. Sci., 59: 156-165.
CrossRefPubMedDirect Link
Jeff Lord Reply
Beauveria is spelled incorrectly in this paper.