
Jaya Vejayan
Faculty of Industrial Science and Technology, Universiti Malaysia Pahang, Lebuhraya Tun Razak, 26300, Gambang, Kuantan, Pahang Darul Makmur, Malaysia
LiveDNA: 60.2180
Amira Alia Zulkifli
Faculty of Industrial Science and Technology, Universiti Malaysia Pahang, Lebuhraya Tun Razak, 26300, Gambang, Kuantan, Pahang Darul Makmur, Malaysia
Rupbansraaj Bathmanathan
Faculty of Industrial Science and Technology, Universiti Malaysia Pahang, Lebuhraya Tun Razak, 26300, Gambang, Kuantan, Pahang Darul Makmur, Malaysia
Halijah Ibrahim
Institute of Biological Sciences, University of Malaya, 50603 Kuala Lumpur, Malaysia
Journal of Biological Sciences
Year: 2020 | Volume: 20 | Issue: 4 | Page No.: 138-146







ABSTRACT
Background and Objectives: The dairy industry is in constant search for newer milk clotting coagulants to substitute rennet due to its growing demands and controversies such as; its origin from young slaughtered animals. Kistomin (EC:3.4.24) is a protease long identified on its role to cleave platelet and fibrinogen. In this study, kistomin discovered to function as a milk coagulant and investigated for its milk clotting activities. Materials and Methods: Kistomin investigated by measuring its Milk Clotting Activity (MCA) and Proteolytic Activity (PA), optimum conditions of pH, temperature, concentrations of enzyme and calcium chloride, exposures to various chelating agents and cofactor ions. Results: The MCA of kistomin was 810.44 (SU mL–1) and the PA was 1.39 (U mL–1), resulted in a ratio of MCA/PA value of 583. The coagulating activity of kistomin on milk was the highest at 0.76 mg mL–1 enzyme concentration, 8% (w/v) of CaCl2 concentration, the temperature of 48°C and stable over a wide range of pH 5-7 with activity peaking at pH6.5. The protease completely inhibited by EDTA and 1,10 phenanthroline verifying to be a metalloprotease. The addition of Ba2+, Mn2+ and Ca2+ significantly increased the enzyme activity but inhibited by Hg2+, Pb2+ and Fe2+ ions. Kistomin promoted extensive hydrolysis of κ-casein and low level of β-casein cleavage. Conclusion: This concluded that kistomin can be used as a milk clotting candidate for the dairy industry.
PDF Abstract XML References Citation
Copyright: © 2020. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Jaya Vejayan, Amira Alia Zulkifli, Rupbansraaj Bathmanathan and Halijah Ibrahim, 2020. Enzymic Milk Clotting Activities of Metalloproteinase Kistomin. Journal of Biological Sciences, 20: 138-146.
DOI: 10.3923/jbs.2020.138.146
URL: https://scialert.net/abstract/?doi=jbs.2020.138.146
DOI: 10.3923/jbs.2020.138.146
URL: https://scialert.net/abstract/?doi=jbs.2020.138.146
INTRODUCTION
Milk is made up of 3-4% total protein with approximately 80% of milk protein is casein and will coagulate or clot at isoelectric pH of casein at pH4.6 or driven by protease hydrolysis to form curd (gel-like) and whey (liquid)1. Since, the milk coagulation properties significantly affect the yield and quality of dairy products such as cheese or yogurt, they are getting more and more attention in recent years2. Enzyme milk clotting can be generally divided into three steps: Activation of milk clotting particles (casein micelles), aggregation of enzyme-modified casein micelles and lastly is the modification for the properties and structure of the coagulum3. During the aggregation of casein micelles, the concentration of calcium ions reduces the electrostatic resistance of micelles by neutralizing the surface charge on casein micelles, hence promoting the casein micelles aggregation4.
The rennet obtained from calf contains more than 90% of chymosin which is responsible for the milk coagulation of milk to curd in the dairy products industry5. Chymosin is a protease secreted in the stomach of young ruminants like calves and kids raising ethical awareness in the slaughtering of such young animals. The advanced technology of recombinant DNA permits the cloning of chymosin using microorganisms3. Additionally, genetic engineering can be employed to enhance protein stability and modulate the activity6. Genetically modified organism or living modified organism experiencing their challenges suspected to cause allergic reactions prompting Genetically Modified (GM) labeling of such products. In the last decade, there has been an increased demand for cheese consumption and production due to the population explosion7. With the increased demands of rennet enzymes, restricted use of animal rennet for certain religions (e.g., porcine rennet for Islam), diets (vegetarianism), safety (bovine spongiform encephalopathy) and activist against genetically engineered foods, other possible alternative sources of rennet are studied and exploited8. Kistomin being a homogenous protease allows the opportunity to be studied on its ability to coagulate milk and as an alternative source for rennet.
The final stage of blood coagulation is the conversion of the soluble fibrinogen to the insoluble fibrin-clot9. This conversion occurs by the hydrolysis of the protease thrombin of fibrinogen molecule10. Meanwhile, for milk clotting, it was stated that ĸ-casein (casein fraction which acts as the substrate of chymosin) is the main protagonist in the primary phase of milk coagulation. According to Rutherfurd and Gill11, there are similarities of sequence between fibrinogen γ-chain and ĸ-casein. The clotting of the blood and milk process possesses a common feature on the action of thrombin on fibrinogen and the action of chymosin on ĸ-casein. It was suggested that they are having a similar mechanism for these enzymatic cleavages. Both processes are said to involve limited proteolysis where both use protease enzymes to hydrolyze specific linkages11. Kistomin cleaves alpha- and gamma-chains of fibrinogen leaving the beta-chain unaffected12. It is one of the metalloproteinases extracted from Calloselasma rhodostoma venom. A metalloproteinase is a protease enzyme where the water molecule that is used for hydrolysis is complexed to a metal ion in the catalytic center of the enzyme and the metal ions itself hold in the position by several amino acid residues13. Kistomin is a 47446 Da molecular weight monomer of 417 amino acids with zinc bound at positions of 333, 337 and 34312,14. Kistomin being a protease needs to be explored as well for its potential in milk clotting. In this study, Kistomin is known to cleave specific blood protein hence rationalized able to also cleave protein specifically casein in milk. Kistomin was investigated for its important features to coagulate milk to become curd and whey.
MATERIALS AND METHODS
Study area: This study conducted between Sept, 2017 and Aug, 2019.
Isolation of kistomin using stepwise chromatography techniques: The purification of kistomin to homogeneity done as previously described in Vejayan et al.15. Briefly, freeze-dried C. rhodostoma venom (purchased from Snake and Reptile Farm, Sungai Batu Pahat, Perlis, Malaysia) fractionated to its active peak using a HiTrap SP FF (GE Healthcare Life Sciences) pre-packed with Sulfopropyl Sepharose Fast Flow, a strong cation connected to the ÄKTAexplorer (Amersham Biosciences, Sweden). A 50 mM ammonium acetate at pH 6.0 and 1.0 M NaCl in 50 mM ammonium acetate at pH 6.0 were the start buffer and elution buffer, respectively. The active fraction from the ion exchange chromatography collected, freeze-dried and consequently desalted by using HiTrap Desalting (GE Healthcare Life Sciences, USA), a prepacked Sephadex G-25 Superfine column. Next, the active fraction eluted from the ion exchange chromatography column was applied onto HiPrep 26/60 Sephacryl S 200 HR pre-packed column (GE Healthcare Life Sciences, USA). The column was equilibrated with 50 mM of ammonium acetate buffer of pH 7 and the elution was carried out using the same buffer with 2 mL sample injection at a flow rate of 1 mL min–1 to yield kistomin known peak (denoted as peak 5b)15. The protein content of the kistomin estimated by using the dye-binding technique of Bradford16 with Bovine Serum Albumin (BSA) at 2.0 mg mL–1 concentration, purchased from Thermo Fisher Scientific, USA.
Milk Clotting Activity (MCA): The milk clotting activity of the kistomin determined by using the method of Arima et al.17 and Majumder et al.18 with few modifications. A 10% (w/v) of BD Difco skim milk (Thermo Fisher Scientific, USA) containing 10 mM of CaCl2 as a substrate. A total of 1 mL of milk substrate was added to the 0.5 mL enzyme solution at 35°C. The milk clotting time was measured according to 3 visible parameters, i.e., viscosity, color changes and the development of white spots19,20. Milk clotting activity was expressed in Soxhlet Unit (SU mL–1). One SU was defined as the enzyme amount which clots 1 mL of the substrate within 40 min at 35°C. The MCA for coagulant was calculated by using the equation:
Where:
| M | = | Volume of substrate (mL) |
| t | = | Clotting time (s) |
| E | = | Volume of enzyme (mL) |
| T | = | Reaction temperature (°C) |
Proteolytic Activity (PA): The proteolytic activity was measured by using 2.5 mL of 1% (w/v) casein (Nacalai Tesque, Japan) in 50 mM potassium phosphate buffer (pH 7.5). The casein solution was equilibrated in a water bath at 37°C for 10 min. A total of 0.1 mL kistomin and a blank (without any coagulant) added to the casein preparations and incubated at 35°C for 30 min. After incubation, 2.5 mL of 440 mM trichloroacetic acid (Sigma Aldrich, USA) was added to stop the reaction. Then the appropriate volume of enzyme solution was added, also in the blank so that the final volume of enzyme solution in each is 1 mL. The solution was incubated again at 35°C for 30 min and was centrifuged at 12000 g for 10 min. A total of 5 mL of 500 mM sodium carbonate (Sigma Aldrich, USA) was added to 2 mL supernatant and 1 mL of Folin and Ciocalteu’s phenol reagent (Sigma Aldrich, USA) was added immediately afterward. The absorbance of the solution was measured at 660 nm.
The standard calibration curve of L-tyrosine (Sigma Aldrich, USA) was prepared between range 0.0-0.60 μMoles. In each 2 mL concentration of tyrosine, a total of 5 mL of 500 mM sodium carbonate and 1 mL of Folin reagent added immediately. The absorbance of the solution was measured at 660 nm by using a UV-V is spectrophotometer.
The calibration curve of L-tyrosine concentration (μMoles) versus Absorbance (660 nm) was constructed and the equation was generated using MS Excel software. The enzyme activity for kistomin calculated by using this equation19:
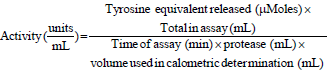 |
Determining optimum parameters of kistomin coagulant: A total of 1 mL of 10% (w/v) skim milk (BD Difco, Thermo Fisher Scientific, USA) containing 10 mM CaCl2 (Sigma Aldrich, USA) was used as the milk substrate (except during the determination of optimum CaCl2 concentration). The temperature and pH maintained to be constant at 35°C and pH 6.5 (except during the determination of optimum temperature and pH).
Enzyme concentration: Kistomin was serially diluted in the range of 0.01-1.28 mg mL–1 protein content. A total of 1 mL of milk substrate was then introduced to 0.5 mL of different concentrations of the coagulant at 35°C, pH 6.5. The time taken for the milk to coagulate was recorded based on three visible parameters, i.e., viscosity, color changes and the development of white spots19,20. The protein concentration was increased until the coagulation rate become constant and the graph of MCA versus concentration was plotted.
pH: The pH stability of kistomin investigated at pH 5-9. Acetate buffer (pH 5), phosphate buffer (pH 6-pH 8) and glycine-NaOH buffer (pH 9) used. Each 0.5 mL of sample solution consisting of coagulant and an appropriate buffer with varying pH of 5- pH 9 was mixed with 1 mL of milk substrate and swirled for 10 sec at 35°C. The time taken for the milk to coagulate was recorded. The graph of MCA versus pH was plotted.
Temperature: The milk substrate (pH 6.5) with kistomin was incubated in a water bath with varying temperature settings i.e., 20-80°C. The time taken for the milk to coagulate was recorded. The graph of MCA versus temperature (°C) was plotted.
Calcium ion concentration: Various concentrations of CaCl2 ranging from 2-14% (w/v) dissolved in 1 mL of 10% (w/v) skim milk at 35°C and pH 6.5. A volume of 0.5 mL of the coagulant was mixed with the skim milk and swirled for 10 sec. The time taken for the milk to coagulate was recorded. The graph of MCA vs. CaCl2 concentration (w/v) was plotted.
Substrate specificity of kistomin: Substrate specificity of coagulant was determined by using a substrate range of 0.05-5.0 mg mL–1 for κ-casein and β-casein while 0.05-8.0 mg mL–1 of α-casein (all purchased from Sigma Aldrich, USA). The substrates were dissolved in 50 mM potassium phosphate buffer (pH 6.5) and were serially diluted. The reaction was initiated by adding 100 μL (0.015 mg) of kistomin and was incubated for 10 min at 48°C.
The reaction stopped by adding 900 μL of trichloroacetic acid (TCA) and the solution was incubated for 30 min. After incubation, the solution was centrifuged at 12000 g for 10 min. One milliliter of supernatant was mixed with 900 μL of 500 mM sodium carbonate and 100 μL of Folin reagent. The absorbance of the solution was measured at 660 nm. The substrate specificity of kistomin toward substrates was compared by using the Km value calculated by the Michaelis Menten equation with nonlinear regression by using GraphPad Prism 7 software.
Effects of protease inhibitor and metal ions on kistomin activity: All materials for this experiment sourced from Sigma Aldrich, USA. The effects of various metal ions and protease inhibitors were determined by using 2% (w/v) of casein dissolved in 50 mM potassium phosphate buffer (pH 6.5) as a substrate. A total of 100 μL of 20 mM of various metal ions (CuSO4, MgCl2, MgSO4, FeSO4, Pb (NO3)2, HgCl2, BaCl2, MnSO4 and CaCl2) and protease inhibitor: 10 mM PMSF, 1 mM Pepstatin A, 10 mM EDTA, 5 mM Iodoacetamide and 10 mM 1,10 phenanthroline was pre-incubated with 100 μL (0.015 mg) protease for 10 min in 48°C. One milliliter of the substrate was then added to each tube containing the mixture of the enzyme and metal ions/inhibitor.
The solution was incubated in the water bath for 30 min at 48°C and the reaction was stopped by adding 900 μL of trichloroacetic acid (TCA). The solution was incubated for 10 min. After incubation, the solution was centrifuged at 12000 g for 10 min. One milliliter of supernatant was mixed with 900 μL of 500 mM sodium carbonate and 100 μL of Folin reagent. The absorbance of the solution was measured at 660 nm. The residual activities were determined and compared to the control which prepared by preincubating the enzyme with buffer and corresponds to 100% activity.
RESULTS AND DISCUSSION
The values determined for MCA, PA and ratio of MCA/PA of kistomin were 810.44±42.45 SU mL–1, 1.39±0.01 U mL–1 and 583.05, respectively. In general term Milk Coagulating Activity (MCA) is the ability of the rennet to aggregate milk by cleaving the amino acid of Phe105-Met106 of κ-casein21. The high value of MCA indicating that the enzyme has good potential as a coagulant in the dairy industry. The second property is Proteolytic Activity (PA) which is the ability of the coagulant to cleave any bond of casein22. A high value of Milk Clotting Activity/Proteolytic Activity (MCA/PA) reflects an excellent end product of cheese with desirable firmness with no bitter flavors. The high ratio of MCA/PA is considered a good coagulant so the higher the value, the better the coagulant and its generally indicates the restricted degradation of casein substrate23-25. The major drawback of the protease extracted from most plant sources has the excessive proteolytic nature and low MCA/PA ratios which limited their use in cheese manufacturing such as ginger, curcumin or hieronyma in protease26,27. The measured value of 583 for kistomin found higher than 367 for papain. Papain has been suggested not useful as a coagulant due to the bitterness of the curd obtained28. Although the MCA/PA ratio calculated for kistomin better than papain nevertheless not possible to ascertain whether this MCA/PA ratio obtained is suitable as a dairy industry protease as there is no sufficient data available on the lower cut-off value for the ratio.
As shown in Fig. 1 the several factors affecting the enzymatic reaction of kistomin in milk clotting. The profiles of the graphs were as expected of an enzyme reacting on a substrate. Initially, to determine the optimum enzyme concentration the temperature and pH maintained constant to 35°C and pH7, respectively while CaCl2 given more than 10 mM. The clotting activity was expressed in Soxhlet Unit (SU). One Soxhlet Unit (SU) of milk clotting activity was defined as29 the amount of enzyme required to coagulate 1 mL of the substrate within 40 min at 35°C. The clotting activity increase when enzyme concentration increased because of a higher level of proteolysis of kappa casein19. The optimum concentration of 0.76 mg mL–1 determined based on Fig. 1a as the highest milk clotting activity achieved by kistomin. Consequently, all other optimum conditions for kistomin with the highest milk clotting activity determined to be 48°C for temperature (Fig. 1b), pH 6.5 (Fig. 1c) and CaCl2 concentration of 8% w/v (Fig. 1d).
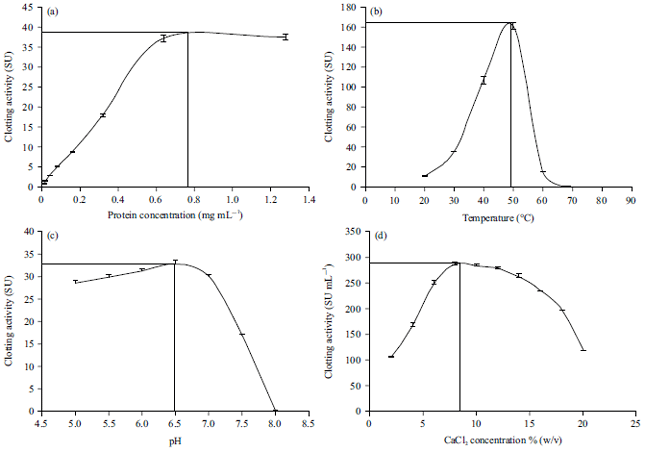
Figure 2a, b showed the milk clotting time of kistomin under optimum conditions for temperature, pH as well as concentrations of enzyme and CaCl2.
 | |
| Fig. 1: | Graph of (a) Milk clotting activities versus enzyme concentration, (b) Temperature, (c) pH, (d) CaCl2 concentration |
| Values are the Mean±Standard Deviation, n = 4 | |
 | |
| Fig. 2(a-b): | Coagulating activity of kistomin on milk at optimum parameters (a) Before Coagulation (b) After coagulation |
The coagulation time determined (n = 4) at optimum conditions of kistomin: Enzyme concentration of 0.76 mg mL–1, 48°C for temperature, pH 6.5 and CaCl2 concentration of 8% (w/v). Observation under light microscope at low magnification (100X) of; (a) Control, bovine skim milk and (b) Coagulated by kistomin | |
The images viewed under microscopic magnification clearly showed the formation of gaps represented by whey after milk coagulated. The observation before coagulation showed milk without any aggregation of casein micelles however subsequently obvious colloidal structure of aggregation occurred.
The optimum temperature30 for metalloproteinase enzyme stated between 37-55°C. The optimum temperature for kistomin reduced the risk of contamination by mesophilic microorganisms that able to grow at a moderate temperature between 20-45°C and with optimum growth temperature between 30-39°C. The elevation of temperature additionally favors substrate and product solubility, increases reaction rates for reducing viscosity and for increasing the diffusion coefficients of substrates31. Higher than room temperature is favored for the milk coagulant in the dairy industry. The commercial calf rennet shows the optimum temperature between 39-45.5°C15,19,32. Genetically engineered calf chymosin for cheese production found with an optimum temperature of 45°C33. This shows that kistomin has potential as a coagulant in the cheese industry as it has a broad range of thermal stability and has a high optimum temperature. Additionally, if required in the future, a recombinant kistomin variant can be generated to lower the optimum temperature in reducing the cost of cheese production.
The kistomin is shown to be favorable in the acidic environment as it stable at pH 5 to pH 6.9 with an optimum pH at 6.5. The amino acid sequence of kistomin was computed by using ProtParam tool-ExPASy online software and the results showed that the total number of negatively charged residues (Aspartic acid and Glutamic acid) is higher than the total number of positively charged residues (Arginine and Lysine)34. This protein also moderately stable at pH 7 which maintained 71% of its activity. In comparison to the chymosin (industrial rennet) found to cleave κ-casein of milk at the optimum pH between 5.1-5.535. In industry additional cost required to be considered to adjust pH to be highly acidic or basic.
The addition of calcium chloride was reported to affect the milk coagulation as it reduces the milk coagulation time36. Calcium chloride speeds up the milk coagulation as calcium ions reduced the electrostatic resistance of micelles by neutralizing the surface charge on casein micelles4. The calcium ions also form bridges between micelles of para casein indicating that the more there are calcium ions in milk, the more there will be linkage19. This promotes the aggregation of casein micelles and leads to faster milk coagulation. However, the addition of calcium chloride beyond what is required leads to the rate of milk clotting activity to gradually decrease. This is due to the high concentration of calcium chloride, Ca2+ ions present inhibits the milk clotting network19. It is therefore necessary in providing the optimum or even slightly lower concentration of CaCl2 rather than resorting to excess for milk clotting activity.
| Table 1: | Kistomin specificity toward various casein substrates |
 | |
| Values are the Mean±Standard deviation, n = 4 | |
| Table 2: | Effect of different protease inhibitors on the activity of kistomin |
 | |
| Values are the Mean±Standard Deviation, n = 4 | |
Table 1 compared the Vmax and the Km of α-casein, β-casein and κ-casein by kistomin. The results showed the Vmax and Km value of kappa-casein is the lowest, 0.0275±0.0019 μmoL min–1 and 27.15±5.277 mM, respectively. This indicated kistomin has a high affinity towards kappa casein and potentially preferred cleaving it to curd and whey fastest. The major enzyme of calf rennet, chymosin has been extensively used in the production of cheese to produce a stable curd with good flavor due to its high specificity for κ-casein37. The specificity of kistomin for casein was similar to chymosin which extensively cleaved κ-casein and promoted very slight hydrolysis of β-casein. The specificity of kistomin on κ-casein followed by α-casein. This can avoid the bitterness development of cheese which is considered as a defect in cheese. It has been reported that the hydrolysis of casein especially β-casein generates hydrophobic peptides which when accumulating is responsible for the bitterness of dairy products. The higher the affinity towards β-casein leads to the product of hydrophobic peptides hence becoming bitter cheese38. In this instance, it was found kistomin having a lower affinity towards β-casein based on higher Km.
Table 2 showed the effect of protease inhibitors on kistomin. Kistomin was incubated with five different protease inhibitor and the residual activities of the protease were determined and compared with the control which corresponds to 100% of activity. Kistomin was completely inhibited in the presence of EDTA and 1,10-phenanthroline and the maintenance of activity in the presence of iodoacetamide (cysteine protease inhibitor), pepstatin A (aspartic protease inhibitor) and PMSF (serine inhibitor) inhibitors. Kistomin is a metalloprotease and as expected both EDTA and 1,10-phenanthroline inhibited it similar to previous observations14,39. EDTA and 1, 10-phenanthroline inhibit metalloprotease activity by lowering the concentration of metal ion and removed it from the active site of the enzyme40.
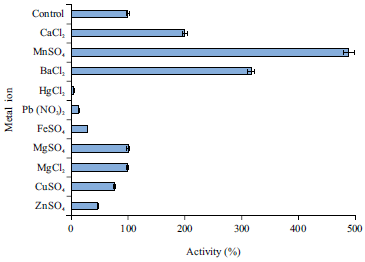 | |
| Fig. 3: | Effects of various metal ions on milk clotting activity of kistomin |
| Values are the Mean±Standard Deviation, n = 4 | |
Figure 3 showed the effects of metal ions on milk clotting activity of the kistomin enzyme. The addition of Mn2+, Ba2+ and Ca2+ (in descending orders) significantly increased the enzyme activity. However, kistomin was inhibited by Hg2+, Pb2+ and Fe2+ ions. The effect of ions over protease stability because of interaction between them, with the formation of complexes such as ions used as cofactors, substrates and co-substrates31. Kistomin is a metalloprotease enzyme and this class of proteases is characterized by its requirement of divalent metal ions for its catalytic activity37. Metalloprotease usually associated with Zn2+, but other metals have been found at the active site of metalloproteases41-43, such as; Mn2+, Ca2+, Mg2+, Cu2+ and Co2+. Pb2+ and Hg2+ were found to be toxic when present in high amount. A similar result was found where the metalloprotease isolated from Oudemansiella radicata was inhibited44 by Hg2+, Pb2+ and Fe2+. Also, Hg2+ also has been reported to inhibit keratinolytic metalloprotease from Microbacterium sp.45.
CONCLUSION
Kistomin is shown as a capable milk coagulant with MCA/PA ratio better than papain and having optimum conditions of 0.76 mg mL–1 enzyme concentration, 48°C temperature, pH6.5 and 8% CaCl2 concentration. Its affinity at least towards β-casein likely producing cheese with less bitter taste. Kistomin completely inhibited by EDTA and 1,10 phenanthroline verifying it as a metalloproteinase and required most the metal ion of Mn2+ as a cofactor.
ACKNOWLEDGMENT
The study was funded by Universiti Malaysia Pahang internal grant (RDU180375) and postgraduate grant (PGRS180354).
SIGNIFICANCE STATEMENT
This study discovered the kistomin function of coagulating milk to curd and whey. The study determined and revealed the protease MCA/PA ratio, rapid coagulation time, specificity towards kappa casein rather than beta-casein, its preferred cofactor and optimum conditions. Overall this work introduces a potential protease in its natural and homogenous state capable to coagulate milk to curd with important features highlighted. Its potential capabilities can be improved further by other researchers by genetic engineering to become an alternative coagulant to chymosin.
REFERENCES
- Troch, T., E. Lefebure, V. Baeten, F. Colinet, N. Gengler and M. Sindic, 2017. Cow milk coagulation: Process description, variation factors and evaluation methodologies. A review. Biotechnol. Agron. Soc. Environ., Vol. 21.
CrossRefDirect Link - Tabayehnejad, N., M. Castillo and F.A. Payne, 2012. Comparison of total milk-clotting activity measurement precision using the Berridge clotting time method and a proposed optical method. J. Food Eng., 108: 549-556.
CrossRefDirect Link - Hyslop, D.B., 2003. Enzymatic Coagulation of Milk. In: Advanced Dairy Chemistry-1 Proteins, Fox P.F. and McSweeney P.L.H. (Eds.)., Springer, US, ISBN: 978-1-4419-8602-3, pp: 839-878.
CrossRefDirect Link - Landfeld, A., P. Novotna and M. Houska, 2002. Influence of the amount of rennet, calcium chloride addition, temperature and high-pressure treatment on the course of milk coagulation. Czech J. Food Sci., 20: 237-244.
CrossRefDirect Link - Kumar, A., S. Grover, J. Sharma and V.K. Batish, 2010. Chymosin and other milk coagulants: Sources and biotechnological interventions. Crit. Rev. Biotechnol., 30: 243-258.
CrossRefDirect Link - Kamionka, M., 2011. Engineering of therapeutic proteins production in Escherichia coli. Curr. Pharm. Biotechnol., 12: 268-274.
CrossRefDirect Link - Tajalsir, A.E., A.S. Ebraheem, A.M. Abdallah, F.J. Khider, M.O. Elsamani and I.M. Ahmed, 2014. Partial purification of milk-clotting enzyme from the seeds of Moringa oleifera. J. Microbiol. Biotechnol. Food Sci., 4: 58-62.
CrossRefDirect Link - Badgujar, S.B and R.T. Mahajan, 2014. Nivulian-II a new milk clotting cysteine protease of Euphorbia nivulia latex. Int. J. Biol. Macromol., 70: 391-398.
CrossRefDirect Link - Jollès, P. and A. Henschen, 1982. Comparison between the clotting of blood and milk. Trends Biochem. Sci., 7: 325-328.
CrossRefDirect Link - Kaye, N.M.C. and P. Jollès, 1978. Characterization of the amino acids of bovine fibrinogen involved in the fibrinogen-thrombin interaction of the blood clotting process. Comparison with the milk clotting process. Mol. Cell. Biochem., 20: 173-182.
CrossRefDirect Link - Rutherfurd, K.J. and H.S. Gill, 2000. Peptides affecting coagulation. Br. J. Nutr., 84: 99-102.
CrossRefDirect Link - Hsu, C.C., W.B. Wu and T.F. Huang, 2008. A snake venom metalloproteinase, kistomin, cleaves platelet glycoprotein VI and impairs platelet functions. J. Thrombosis Haemostasis, 6: 1578-1585.
CrossRefDirect Link - Takeda, S., H. Takeya and S. Iwanaga, 2012. Snake venom metalloproteinases: Structure, function and relevance to the mammalian ADAM/ADAMTS family proteins. Biochim. Biophys. Acta (BBA), Proteins Proteomics, 1824: 164-176.
CrossRefDirect Link - Huang, T.F., M.C. Chang, H.C. Peng and C.M. Teng, 1992. A novel α-type fibrinogenase from Agkistrodon rhodostoma snake venom. Biochim. Biophys. Acta (BBA), Protein Struct. Mol. Enzymol., 1160: 262-268.
CrossRefDirect Link - Vejayan, J., A.A. Zulkifli, S.M. Saufi, N. Munir, H. Ibrahim and S. Ambu, 2017. Uncovering a protease in snake venom capable to coagulate milk to curd. Int. J. Adv. Biotechnol. Res., 8: 409-423.
Direct Link - Bradford, M.M., 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem., 72: 248-254.
CrossRefPubMedDirect Link - Arima, K., J. Yu and S. Iwasaki, 1970. Milk-Clotting Enzyme from Mucor pusillus var. Lindt. In: Methods in Enzymology, Galluzzi, L. and N.P. Rudqvist (Eds.)., Elsevier, London, pp: 446-459.
CrossRefDirect Link - Majumder, R., S.P. Banik and S. Khowala, 2015. Purification and characterisation of κ-casein specific milk-clotting metalloprotease from Termitomyces clypeatus MTCC 5091. Food Chem., 173: 441-448.
CrossRefDirect Link - Najera, A.I., M. De Renobales and L.J.R. Barron, 2003. Effects of pH, temperature, CaCl2 and enzyme concentrations on the rennet-clotting properties of milk: A multifactorial study. Food Chem., 80: 345-352.
CrossRef - Vejayan, J., N. Munir, L.L. Yee, R. Bathmanathan, H. Ibrahim and S. Chakravarthi, 2019. Tropical fruits of mango and noni having dual effects of coagulating milk and enriching the curds with micro-constituents of medicinal potential. Malaysian J. Sci., 38: 34-47.
CrossRefDirect Link - Egito, A.S., J.M. Girardet, L.E. Laguna, C. Poirson, D. Molle, L. Miclo, G. Humbert and J.L. Gaillard, 2007. Milk-clotting activity of enzyme extracts from sunflower and albizia seeds and specific hydrolysis of bovine k-casein. Int. Dairy J., 17: 816-825.
CrossRefDirect Link - Kappeler, S.R., H.J.M. van der Brink, H. Rahbek-Nielsen, Z. Farah, Z. Puhan, E.B. Hansen and E. Johansen, 2006. Characterization of recombinant camel chymosin reveals superior properties for the coagulation of bovine and camel milk. Biochem. Biophys. Res. Commun., 342: 647-654.
CrossRef - Nasr, A.I.A.M., I.A.M. Ahmed and O.I. Hamid, 2016. Characterization of partially purified milk‐clotting enzyme from sunflower (Helianthus annuus) seeds. Food Sci. Nutr., 4: 733-741.
CrossRefDirect Link - Langholm Jensen, J., A. Mølgaard, J.C. Navarro Poulsen, M.K. Harboe and J.B. Simonsen et al., 2013. Camel and bovine chymosin: The relationship between their structures and cheese-making properties. Acta Crystallogr., Sect. D: Biol. Crystallogr., 69: 901-913.
CrossRefDirect Link - Sato, S., H. Tokuda, T. Koizumi and K. Nakanishi, 2004. Purification and characterization of an extracellular proteinase having milk-clotting activity from Enterococcus faecalis TUA2495L. Food Sci. Technol. Res., 10: 44-50.
CrossRefDirect Link - Amira, A.B., S. Besbes, H. Attia and C. Blecker, 2017. Milk-clotting properties of plant rennets and their enzymatic, rheological and sensory role in cheese making: A review. Int. J. Food Proper., 20: S76-S93.
CrossRefDirect Link - Shah, M.A., S.A. Mir and M.A. Paray, 2014. Plant proteases as milk-clotting enzymes in cheesemaking: A review. Dairy Sci. Technol., 94: 5-16.
CrossRefDirect Link - Lemieux, L. and R.E. Simard, 1991. Bitter flavour in dairy products. I. A review of the factors likely to influence its development, mainly in cheese manufacture. Le Lait, 71: 599-636.
Direct Link - Abd El-Salam, B.A.E.Y., O. Abd El-Hamid Ibrahim and H.A. El-Razek El-Sayed, 2017. Purification and characterization of milk clotting enzyme from artichoke (Cynara cardunculus L.) flowers as coagulant on white soft cheese. Int. J. Dairy Sci., 12: 254-265.
CrossRefDirect Link - Mazzi, M.V., S. Marcussi, G.B. Carlos, R.G. Stábeli and J.J. Franco et al., 2004. A new hemorrhagic metalloprotease from Bothrops jararacussu snake venom: Isolation and biochemical characterization. Toxicon, 44: 215-223.
CrossRefDirect Link - Merheb-Dini, C., H. Cabral, R.S.R. Leite, L.M. Zanphorlin and D.N. Okamoto et al., 2009. Biochemical and functional characterization of a metalloprotease from the thermophilic fungus Thermoascus aurantiacus. J. Agric. Food Chem., 57: 9210-9217.
CrossRefDirect Link - Mamo, A. and N. Balasubramanian, 2018. Calf rennet production and its performance optimization. J. Applied Natural Sci., 10: 247-252.
CrossRefDirect Link - Rogelj, I., B. Perko, A. Francky, V. Penca and J. Pungerar, 2001. Recombinant lamb chymosin as an alternative coagulating enzyme in cheese production. J. Dairy Sci., 84: 1020-1026.
CrossRefDirect Link - Gasteiger, E., C. Hoogland, A. Gattiker, S. Duvaud, M.R. Wilkins, R.D. Appel and A. Bairoch, 2005. Protein Identification and Analysis Tools on the ExPASy Server. In: The Proteomics Protocols Handbook, Walker, J.M. (Ed.). 1st Edn., Humana Press, New Jersey, USA., ISBN-13: 978-1588295934, pp: 571-607.
- Shalabi, S.I. and P.F. Fox, 1982. Influence of pH on the rennet coagulation of milk. J. Dairy Res., 49: 153-157.
CrossRefDirect Link - Sandra, S., M. Ho, M. Alexander and M. Corredig, 2012. Effect of soluble calcium on the renneting properties of casein micelles as measured by rheology and diffusing wave spectroscopy. J. Dairy Sci., 95: 75-82.
CrossRefDirect Link - Rao, M.B., A.M. Tanksale, M.S. Ghatge and V.V. Deshpande, 1998. Molecular and biotechnological aspects of microbial proteases. Microbiol. Mol. Biol. Rev., 62: 597-635.
Direct Link - Savijoki, K., H. Ingmer and P. Varmanen, 2006. Proteolytic systems of lactic acid bacteria. Applied Microbiol. Biotechnol., 71: 394-406.
CrossRefDirect Link - Hsu, C.C., W.B. Wu, Y.H. Chang, H.L. Kuo and T.F. Huang, 2007. Antithrombotic effect of a protein-type I class snake venom metalloproteinase, kistomin, is mediated by affecting glycoprotein Ib-von Willebrand factor interaction. Mol. Pharmacol., 72: 984-992.
CrossRefDirect Link - Hazra, S., R. Guha, G. Jongkey, H. Palui and A. Mishra et al., 2012. Modulation of matrix metalloproteinase activity by EDTA prevents posterior capsular opacification. Mol. Vision, 18: 1701-1711.
Direct Link - Asoodeh, A. and H.M. Musaabadi, 2012. Purification and characterization of a thermostable neutrophilic metalloprotease from Pseudomonas sp. DR89. Iran. J. Biotechnol., 10: 120-128.
Direct Link - Graham, S.C., C.S. Bond, H.C. Freeman and J.M. Guss, 2005. Structural and functional implications of metal ion selection in aminopeptidase P, a metalloprotease with a dinuclear metal center. Biochemistry, 44: 13820-13836.
CrossRefDirect Link - Hase, C.C. and R.A. Finkelstein, 1993. Bacterial extracellular zinc-containing metalloproteases. Microbiol. Rev., 57: 823-837.
Direct Link - Geng, X., R. Te, G. Tian, Y. Zhao, L. Zhao, H. Wang and T.B. Ng, 2017. Purification and characterization of a novel metalloprotease from fruiting bodies of Oudemansiella radicata. Acta Biochim. Pol., 64: 477-483.
CrossRefDirect Link - Thys, R.C.S. and A. Brandelli, 2006. Purification and properties of a keratinolytic metalloprotease from Microbacterium sp. J. Appl. Microbiol., 101: 1259-1268.
CrossRefDirect Link