
Dwi Sutiningsih
Department of Epidemiology and Tropical Disease, Faculty of Public Health, University of Diponegoro, Semarang, Indonesia
LiveDNA: 62.16151
Nurjazuli
Department of Environmental Health, Faculty of Public Health, University of Diponegoro, Semarang, Indonesia
Journal of Biological Sciences
Year: 2017 | Volume: 17 | Issue: 3 | Page No.: 127-135







ABSTRACT
Background and Objective: Brusatol is one of quassinoid, derived from Makassar fruit (Brucea javanica (L.) Merr.) and belong to the family of Simaroubaceae. Brusatol has been proven to have insecticidal and anti-feeding activity on larvae of Spodoptera exigua. This study aimed to investigate the larvicidal activity and behavioral response to the larvae of A. aegypti after administration of brusatol and to demonstrate cytotoxicity activity in vero cells. Materials and Methods: Larvicidal activity was monitored using bioassay test. The behavioral response of larval A. aegypti was observed after treatment with various brusatol concentrations and compared directly with control every 10, 30 min and 1, 5 and 24 h. The examination of brusatol cytotoxicity on vero cells performed by MTT (3-[4,5-dimethylthiazol-2-yl]-2,5 diphenyltetrazolium bromide) method. Results: The results of this study showed that the behavioral response of A. aegypti larvae after treatment with brusatol has similarity with the behavioral response after treatment with temephos 1 ppm. The behavioral response of A. aegypti larvae showed symptoms of a nerve poison that is excitation, convulsions, tremors and paralysis. The lethal concentration of 50 and 90 (LC50, LC90) to A. aegypti larvae were 0.669±0.106 and 8.331±0.060 ppm, respectively. Brusatol cytotoxic activity on vero cells is low, with IC50 values worth 1402.029±0.164 mg mL–1. Conclusion: The results of this study proved the potential of brusatol as biolarvicide to control the mosquito vector with low levels of cytotoxicity.
PDF Abstract XML References Citation
Received: February 02, 2017;
Accepted: March 08, 2017;
Published: March 15, 2017
Copyright: © 2017. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Dwi Sutiningsih and Nurjazuli, 2017. Effect of Brusatol Biolarvicide Administration on Behavioral Response of Aedes aegypti and its Toxicity on Vero Cells. Journal of Biological Sciences, 17: 127-135.
DOI: 10.3923/jbs.2017.127.135
URL: https://scialert.net/abstract/?doi=jbs.2017.127.135
DOI: 10.3923/jbs.2017.127.135
URL: https://scialert.net/abstract/?doi=jbs.2017.127.135
INTRODUCTION
Dengue Hemorrhagic Fever (DHF) until today is one of the diseases which becomes a serious public health problem in Indonesia. This disease is transmitted by the Aedes aegypti mosquito1. One method to limit the spread of dengue is by using larvicides. In 1980, temephos 1% (abate) established as part of a mass eradication program to eradicate Aedes aegypti in Indonesia. But using temephos repeatedly may lead to the emergence of resistance of various species of mosquitoes which are also known as disease’s vectors2. Development of new insecticides which do not pose a risk of resistance and environmentally more friendly can be obtained through the use of biolarvicide3. Bioinsecticide or biological insecticide is an insecticide, its base ingredients derived from plants that contain chemical substances (bioactive) which is toxic to insects and easy to decompose (biodegradable) in nature so it does not pollute the environment and relatively are safe for humans. In addition, biological insecticide is selective4. The toxicity power comes from its toxic substances that contained in biological insecticide. These substances can act as toxins through the digestive tract absorption or through the skin on the animals which have tender body5.
Brusatol is a quassinoid derived from B. javanica (L.) Merr. fruit. Its molecular formula is C26H32O11 with amorphous powder and bitter taste. Zhang et al.6 have proved that brusatol isolated from B. javanica (L.) Merr. has insecticidal and antifeedant properties against instar 3 larvae of Spodoptera exigua. Syahputra7 and Lina et al.8 have also demonstrated that the active material contained in B. javanica (L.) Merr. extract has larvicidal effect against Crocidolomia pavonana (Lepidoptera: Crambidae). Extract of B. javanica (L.) Merr. can inhibit feeding, decrease the rate of growth and inhibit laying C. pavonana imago9. These informations showed that the study of B. javanica (L.) Merr. and brusatol compound as larvicides/insecticides is limited to the use of agricultural pests. Therefore, this study aimed to investigate the more detail on the brusatol larvicidal activity and the effect on behavior of the larvae of A. aegypti after treatment with brusatol concentration and cytotoxicity in normal cells (vero cells).
MATERIALS AND METHODS
Makasar fruit (B. javanica (L.) Merr.) was purchased from a wholesaler of medicinal plants (Herbal Merapi Yogyakarta) and further identified in the Laboratory of Pharmaceutical Biology of the Faculty of Pharmacy Gadjah Mada University to obtained confirmation of the simplisia data information. The instar III larvae of A. aegypti until the beginning of instar IV was acquired and developed at the Laboratory of Parasitology, Faculty of Medicine, Gadjah Mada University. Vero cells were obtained from the Laboratory of Parasitology, Faculty of Medicine, University of Gadjah Mada. The isolation and identification of brusatol were performed in the Laboratory of Biology Pharmacy, Faculty of Pharmacy, Gadjah Mada University. The behavior testing of the larvae of A. aegypti and the brusatol cytotoxicity on vero cells were undertaken at the Laboratory of Parasitology, Faculty of Medicine, Gadjah Mada University in Yogyakarta.
Extraction and isolation of brusatol: The isolation of brusatol from the seeds of B. javanica (L.) Merr. was undertaken using the method of Zhang et al.6 with a slight modification; Brucea javanica (L.) Merr. dried seeds (5 kg) made it into powder, shaken with EtOH-H2O (15 L), the solvent was evaporated in vacuum and the extracts were combined and concentrated, followed by suspending in H2O. The aqueous layer was further extracted with petroleum ether, CHCl3, EtOAc and n-Butanol. The CHCl3 layer was evaporated under vacuum to afford extracts. This extract was re-suspended with CHCl3 and then chromatographed on a silica gel column (400 g, 200-300 mesh) by eluting successively with CHCl3 containing increasing amounts of MeOH (1:0, 50:1, 20:1, 10:1, 5:1 and 0:1). The result in the organic layer is taken and evaporated to obtain a concentrate and then made a solution of MeOH (100-250 mL) at a temperature of 60°C and then stored at room temperature. MeOH solution above when it is allowed to stand at room temperature to form crystalline compounds that expected as brusatol. Further separation is by filtration. The rest of the filtrate/residue is separated by Thin Layer of Chromatography (TLC). The filtrate/residue is evaporated. Further separation is performed if it is necessary.
Larvicidal test: Larvicidal activity test was conducted using bioassay method according to the standard of WHO10 with a slight modification. Initially it was performed a preliminary test to determine the range of brusatol concentration that can be deadly to larvae of A. aegypti on instar III until the beginning of instar IV. Furthermore, the concentrations used in further tests with positive control is temephos 1 ppm whereas the negative control is used only as much as 100 mL of distilled water, repeated 4 times. The larvae of A. aegypti on the end of instar III until the beginning of instar IV are used as much as 25 larvae put in each treatment media and control. After 24 h, the larvae of A. aegypti that were dead were counted. Temperature, pH of the media and humidity in the room were measured at the beginning and the end of the study.
Behavioral change test: Behavioral change was measured using the method described by Sharma et al.11 with a slight modification. The instar III larvae of A. aegypti until the beginning of instar IV were put in a plastic cup filled with brusatol with various concentrations (0.33, 0.6, 1, 3, 9 and 27 ppm), each of 25 larvae, which consisted of four replications with positive control is temephos 1% while the negative control is used only as much as 100 mL of distilled water. Change in larval behavior was monitored every 10 min, 30, 1, 5 and 24 h after treatment. Data on larval behavior was observed by speed/activity of movement, physical response, posture and behavior. Data was recorded and photographed using Nikon D5300 camera. Similar observations were performed in the control group and then compared to the treatment group.
Brusatol cytotoxicity test: Brusatol cytotoxicity test in vero cells was conducted using MTT (3- [4,5-dimethylthiazol-2-yl] -2,5 diphenyltetrazolium bromide) based on the method of Van Meerloo et al.12 with a slight modification. Vero cells were cultured using M199 medium which has added 10% FBS, 2% penicillin-streptomycin and 0.5-1% fungison. Vero cells were taken from liquid nitrogen, heated at 37°C until liquefied. The cell suspension that has liquefied put into conical tube and washed with M199 complete medium. The cell suspension was then transferred into a culture flask and incubated in a 37°C incubator, 5% CO2. Cell growth was observed with inverted microscope every day until the cells almost meet the base wall of flask. Vero cell harvesting is done after the flask was almost completely full. Cells were washed with PBS and added with 0.25% trypsin for the cells detached from the flask wall. The cell suspension was made by adding complete medium, after which the number of cells were counted with a haemocytometer. Cell is inserted into a 96 well microplate at a density of 2×104 cells/well in 100 μL. Then the culture medium containing brusatol for each variation of the concentration (1200, 600, 300, 150, 75, 37.5 μg mL–1) was added. Cell cultures were incubated for 24 h in an incubator temperature of 37°C, 5% CO2. Cell growth was observed using MTT then compared with the control culture (without treatment). Medium discarded after incubation mass ends then added 100 μL of complete medium and 10 μL solution of MTT. Furthermore it was incubated for 4 h in an incubator at 37°C, 5% CO2. Then it was added 100 μL of 10% SDS in 0.01 M HCl to dissolve formazan which was formed and incubated overnight at room temperature. The test result was read by ELISA at a wavelength of 595 nm.
Data analysis: Probit analysis was conducted on larval mortality after 24 h exposure to different concentrations of brusatol using SPSS version 24. If the data were normally distributed and homogeneous then the data were analyzed using one-way ANOVA test, whereas if the data were not normally distributed and not homogeneous the data were tested using the Kruskal-Wallis test, with a reliability level of 0.05 to determine whether there are differences between the mortality of A. aegypti larvae in various concentration of treatment.
The data of viable cell obtained is calculated inhibition percentage using the formula in Eq. 132:
| (1) |
where, A is absorbance of treatment (cells+media +compound), B is absorbance of control samples (media+compound), C is absorbance of control cells (cells+media) and D is absorbance media control.
The value of IC50 was calculated using probit regression analysis using SPSS version 24. The small value of IC50 indicates that the compound has a high cytotoxic effect and vice versa the big value of IC50 suggests that the cytotoxic effect is low.
RESULTS
Larvicidal activity and behavior of A. aegypti after treatment with brusatol: The environment considered in this study was the pH of the media, media temperature and humidity. The results of the measurement of media temperature, pH and humidity in the room of the study respectively was 25°C, 7 and 71-72%. The percentage of the mortality of A. aegypti larvae after observation for 24 h was presented in Table 1. The lethal concentration of 50 and 90 (LC50 and LC90) calculated by probit analysis using SPSS version 24 were 0.669±0.106 and 8.331±0.060 ppm respectively. Kruskal Wallis test results showed the value of p = 0.000 (p<0.05), the average number of deaths of A. aegypti larvae was significantly different.
Observation of behavioral response of A. aegypti larvae after treatment with brusatol was conducted directly by looking at changes in the physical and behavioral response or body movements of A. aegypti larvae as compared with controls. The result of behavioral response observation on the larvae of A. aegypti is shown in Table 2. Observation of behavioral responses to larvae of instar III A. aegypti and early instar IV after brusatol treatment showed a different pattern of behavior compared to the control. Immediately after brusatol exposure, all the larvae of A. aegypti indicated a natural and powerful movement (natural and vigorous movement).
| Table 1: | Average of percentage of the mortality of A. aegypti larvae in the group treated with brusatol in wide range of concentrations, temephos of 1 ppm and control after 24 h of observation |
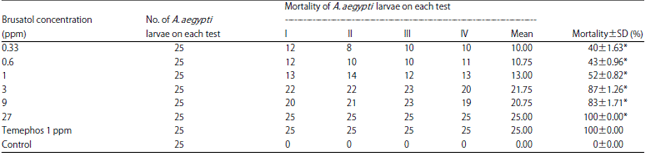 | |
| *p<0.05 with Kruskal Wallis test, ppm: Part per million, SD: Standard deviation | |
But after 10 min of brusatol exposure, larvae of A. aegypti indicated anxiety, excitation and agitation as well as on the provision of temephos 1 ppm. Then the larvae would quickly writhe around 10-15 min and began to sink in the bottom of the water. Most larvae showed heavy anxiety response and very aggressive by biting their gills anal with its mouth so that it looked like a circular motion to form a ring structure (Fig. 1).
Brusatol cytotoxicity on vero cells: The average of alive vero cells observation and the percentage of viable cells after brusatol administration and the control can be seen in Table 3. The percentage of vero cells which were alive reaching 100±0.014% at the lowest concentration of brusatol which is 37.5 μg mL–1, whereas at the highest concentration of brusatol which is 1200 μg mL–1, the percentage of viable cells is 49.897±0.217%. Results of probit analysis using SPSS program version 24 obtained value of IC50 of brusatol on vero cells at 1402.029±0.164 μg mL–1. Kruskal Wallis test results showed the value of p = 0.153 (p>0.05), the average of vero viable cell was not significantly different.
DISCUSSION
Larvicidal activity and behavior of A. aegypti (L.) after treatment with brusatol: The environment that is considered in this study is the pH of the media, media temperature and humidity. The levels of pH, media temperature and humidity measured at the beginning and the end of the study. The result of media temperature measurement shows that the temperature of the media is still in the optimal temperature range (25-27°C) for the development of A. aegypti that is 25°C. The pH value of the media was also in the optimal pH range (6.5-7) for larval development A. aegypti that is 7. The results of the measurement of humidity in the room of the study ranged between 70-71%, that is still within the range of normal humidity for larval development A. aegypti that is 60-80%10. From these results it can be concluded that the mortality of larvae A. aegypti is not caused by environmental factors such as pH media, media temperature and humidity.
The results of 24 h observation showed that there is no larval mortality A. aegypti on control group, so that the number of dead larvae of A. aegypti in the treatment group did not need to be corrected by Abbot formula. At the lowest concentrations of brusatol that is 0.33 ppm larval mortality of A. aegypti is occurred by 40±1.63%. Larval mortality of A. aegypti reached 100% at a brusatol concentration of 27 ppm which is equivalent to giving temephos at a concentration of 1 ppm (Table 1). This means that brusatol is effective as a natural larvicide on the larvae of A. aegypti. The higher the concentration of brusatol , the higher the mortality of larvae of A. aegypti. The mortality of A. aegypti in this study is due to the toxic activity from the brusatol compound. Toxicity of brusatol on A. aegypti larvae stated with the lethal concentration of 50 and 90 (LC50 and LC90) which amounted to 0.669±0.106 and 8.331±0.060 ppm. The smaller the number of lethal concentration the compound is more toxic. Kruskal Wallis test results showed that the average number of deaths of A. aegypti larvae is significantly different. The difference in larval mortality of A. aegypti larvae is likely to be influenced by several factors, namely instar larvae and differences in the sensitivity of each test larvae. Factor larval instar is related to age of the larvae. Although the larvae used were instar III larvae until the early of instar IV, the varieties of larval age is remained. Early instar III larvae are more susceptible to insecticides than the early instar IV larvae13.
| Table 2: | Behavior of A. aegypti larvae after administration of wide range of brusatol concentrations, 1 ppm of temephos and control over the time period of observation |
 | |
| ppm : Part per million | |
 | |
| Fig. 1(a-b): | Digital photomicrographs of early IV instar larvae of A. aegypti, (a) Control larvae and (b) Larvae treated with brusatol 9 ppm |
| Circled larvae show the aggressive anal gill biting behaviour forming ring-shaped structure | |
In addition, differences in larval mortality is influenced also by the differences in the sensitivity of each test larvae. Although using larvae with the same instar and age, each larva has a different level of vulnerability14.
Brusatol is a toxic substances contained in Makasar fruit (B. javanica (L.) Merr.) which gets into the body of A. aegypti larvae through the skin, mouth and respiratory tract. From Table 2 it can be seen that difference in the behavioral response of A. aegypti larvae between the normal larvae (control) and larvae which got treatment with brusatol and 1 ppm of temephos. About 10 min observation after giving brusatol, A. aegypti larvae showed movement agitation, excitation and restlessness, while by giving 1 ppm temephos showed their anxiety and tremor. Meanwhile, normal larvae (control) showed agile and strong mobility. Thirty minutes later looked aggressive and circular movements by biting annals gills and formed a ring on A. aegypti larvae after giving brusatol.
| Table 3: | Average Vero cells were alive and the percentage of viable cells after brusatol administration and control |
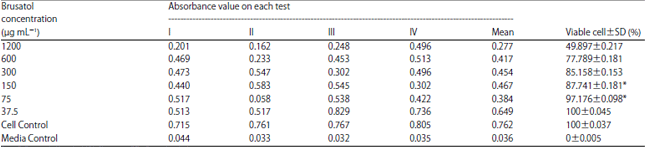 | |
| *p<0.05 with Kruskal Wallis test, SD: Standard deviation | |
The results are consistent with study of Sharma et al.11 who proved that early instar IV of A. aegypti larvae shows behavioral agitation, excitation, restlessness and aggressive movement by biting anal gills after exposure of leaf extract of Achyranthes aspera and stem extracts of A. aspera. These symptoms are similar to symptoms caused by a neurotoxin, meaning that brusatol might act as sitolisin that affect neuromuscular larvae system. Circular movement of the larvae, accompanied by aggressive biting anal gills behavior proved that there are neurotoxic symptoms of brusatol on A. aegypti larvae (Fig. 1b). Anal gills on mosquitoes have some role in controlling electrolyte balance needed for survival15. The effects of brusatol toxic caused electrolyte imbalance in the anal area, causing violent by biting anal gills. The results of this study were similar to results of previous studies which indicate that bioactive plant compounds can act as a neurotoxin although aggressive and un-coordinated movements together with other toxic symptoms were observed at relatively different time intervals16-20. Observation of similar behavior has been reported by Warikoo and Kumar21 on the larvae of A. aegypti tested with root extract of Argemone mexicana. The result of microscopic observation on the early instar IV of A. aegypti larvae after treatment by giving concentration of A. mexicana extract 91.331 and 156.684 ppm for 24 h showed a structural deformity of anal gills. This damage may cause the function anomalies that may have led to interference in the regulation of the osmotic and ionic20. Malfunctioning of this regulation may lead to the death of the larvae of A. aegypti. One hour after treatment with brusatol, larvae of A. aegypti showed the movement of restlessness, tremors, convulsions and paralysis that increasingly powerful that eventually most larvae die, it also occurred in the larvae of A. aegypti after the administration of 1 ppm of temephos. This means that there was a similarity to the behavior of larvae of A. aegypti after administration brusatol with 1 ppm of temephos. Sanjaya and Safaria22 mentioned that the toxic compounds that enter the body can cause a four-stage larvae responses which are excitation, convulsions (spasticity), paralysis (paralysis) and mortality. Brusatol is a toxic substance that can kill larvae of A. aegypti, larvae enters the body through the pores of the larvae’s skin, gastrointestinal tract and respiratory tract. The toxic compounds caused the disruption of the digestive system, the respiratory system and the nervous system in larvae18-20. These compounds caused sensory nerve cells that are on the surface of the larval body disturbed. The mechanism of action of brusatol is performed by inhibiting acetylcholinesterase enzyme which serves to hydrolyze the acetylcholine. In the normal situation, the acetylcholine functions to transmit nerve impulses, after that, acetylcholine is hydrolyzed by the acetylcholinesterase enzyme into choline and acetic acid23. The bound acetylcholinesterase enzyme causes the cumulation of acetylcholine, so that results in disruption and damage to the nervous system24. It makes the muscles continue to contract until exhaustion and then leads to paralysis and death.
Brusatol enters the body of larvae through the skin membrane by diffusion. A number of toxic substances that enter the body cause damage to the skin cells. These toxic compounds hydrolyze the skin membrane by remodeling the proteins of the skin (collagen) into several sections. The damage to the skin cell membrane causes the loss of the skin membrane impermeability. Hence, the other toxic compounds freely enter the body of the larvae. The number of toxic compounds entering the body causes damage to the proteins in the skin membrane so that the function of the skin as a protection of the body is disrupted25.
In addition to the diffusion of the skin membrane, brusatol supposedly enters through the gastrointestinal tract. The gastrointestinal tract of the mosquito larvae consists of three parts, which are the front part (foregut), middle (midgut) and back (hindgut) of the gastrointestinal tract. The process of food digestion and absorption occurs in the middle of the gastrointestinal tract/midgut22. In the middle of the gastrointestinal tract/midgut, it is coated by the epithelial tissue. Toxic substances enter through the mouth into the middle of the gastrointestinal tract/midgut. These toxic substances cause the lysis of the epithelial cells resulting in the reduction of the tension on the surface of the mucous membranes of the middle of gastrointestinal tract/midgut. Thus, the food digestion and absorption might not occur25.
Another way brusatol enter the body of larvae is through the respiratory tract. Air enters through a siphon attached to the surface of the water. This toxic substance is supposed to cover the medium surface, so it blocks the siphon to get oxygen from the medium surface22,25. The nervous tissue of the larvae is very sensitive towards the lack of oxygen, causing the wilting of the nerves and damage of the siphon. Therefore, the larvae face the difficulty in breathing and eventually dying26.
Brusatol as an antifeedant can inhibit the feeding activity of the larvae of A. aegypti. According to Zhang et al.6, brusatol isolated from B. javanica (L.) Merr. have an insecticidal and antifeedant effect to the third instar larvae of Spodoptera exigua. Further study showed that brusatol can induce apoptosis in insect cell lines IOZCAS-Spec-II and Sf21 characterized by the DNA fragmentation, activation of caspase-3 and the enzyme cytochrome-c release from mitochondria. Antifeedant is a chemical compound that inhibits the feeding activity of insects but will not destruct, expel or entrap the insects directly. Antifeedant compound only inhibits the appetite (feeding inhibition) in insects. Antifeedant compound is suppressant (suppress the activity of biting) and a deterrent (prevent the insects for eating continuously) towards insects27. As another antifeedant compounds, brusatol is considered to be able to destruct larvae indirectly by causing disturbances in the signal transmission system of the appetite stimulant (phagostimulant) on the insects. The low feeding activities of the larvae caused the energy to be reduced so that the larval growth process is inhibited and eventually die. In addition, the bitter taste of brusatol6 also caused the larvae lose their appetite which leads to starving and death of the larvae.
Temephos is one of pesticide belongs to class of organic phosphate compounds that can enter the body through the skin, inhaled through breathing and ingested through the mouth. Organic phosphate poisoning in insects followed by restlessness, hyperexcitability, tremors, convulsions and eventually muscle paralysis (paralysis)28. Mortality of A. aegypti larvae reached 100% after 24 h of exposure to brusatol concentration of 27 ppm, while the 1 ppm of temephos giving 100% mortality of larvae of A. aegypti reached after 5 h of treatment. This means that the time required to turn off 100% of A. aegypti larvae is quickly achieved after administration of 1 ppm of temephos than 27 ppm of brusatol. This is because temephos is more quickly absorbed by the larvae of A. aegypti compared with brusatol at the same time. Temephos contains phosphorothionate group which has lipophilic nature so it is able to penetrate to the waxy epicuticular layer of A. aegypti larvae which is hydrophilic so it is more toxic than brusatol. Temephos toxicity against mosquito larvae is closely related to how much the molecule is oxidized or it can be said that the more oxidized, toxicity decreases. Temephos inhibits the cholinesterase enzyme both in vertebrates and invertebrates, causing disruption in neural activity as a continuous muscle contractions, convulsions and ultimately insects will die29. Metabolism of temephos namely fosforotionat group (P = S) in the body of the animal was changed to phosphate (P = O) is more potential as anticholinesterases. The larvae of A. aegypti was able to change the P = S to P = O ester faster than house flies as well as temephos penetration into larvae is rapid where more than 99% of temephos in the medium is absorbed within 1 h after treatment. After the absorption, temephos is converted into metabolic products which some of these metabolic products are excreted into the water28,29.
Brusatol cytotoxicity on vero cells: Cytotoxic test in this study was conducted to determine the brusatol toxicity on normal cells. Cytotoxic test is used to determine the IC50 value parameters. IC50 value indicates the concentration which produces barriers of cell proliferation as much as 50% and demonstrates the potential toxicity of a compound to cell30. IC50 values may indicate the potential of a compound as cytotoxic. The higher the value of IC50 against vero cells means the compounds are less toxic to normal cells31. The end of the cytotoxicity test can provide information on the percentage of cells that are able to survive. Table 3 shows the percentage of viable cells after administration of brusatol growing increasingly at lower concentrations. In brusatol concentration of 1200 μg mL–1, the percentage of viable cells is 49.897±0,217%, while the lowest concentration of 37.5 μg mL–1, the percentage of viable cells is 100±0.045%. Probit analysis results showed IC50 value of brusatol against vero cells is 1402.029±0.164 μg mL–1 but not significantly different (p>0.05). It means brusatol compound has cytotoxic effects categorized as low (low cytotoxicity)26 because it has a value of IC50>30 μg mL–1. Similar observations have been reported by Mangungsong32 who reported that the compound of brusatol isolated from the seed of B. javanica (L.) Merr. has a low cytotoxic effect on vero cells with IC50 value of 1366.55 μg mL–1.
CONCLUSION
This study can be concluded that the potential effect of brusatol as biolarvicide to control the mosquito vector with low levels of cytotoxicity on normal cells (Vero cells). The behavioral response of A. aegypti larvae showed symptoms of a neurotoxic that is excitation, convulsions, tremors and paralysis.
SIGNIFICANCE STATEMENT
Brusatol isolated from the seeds of Makasar fruit (Brucea javanica (L.) Merr.) is potentially effective and safe in controlling mosquito vector especially A. aegypti. Brusatol biolarvicide is proven significantly to have larvicidal effect on A. aegypti with the LC50 of 0.669±0.106 ppm and LC90 that is at 8.331±0.060 ppm.
ACKNOWLEDGMENTS
The authors would like to thank the Rector of University of Diponegoro who has funded this study through PNBP DIPA University of Diponegoro funding No. SP DIPA-042.01.2.400898 / 2016, dated December 7, 2015 of fiscal year 2016.
REFERENCES
- Ndione, R.D., O. Faye, M. Ndiaye, A. Dieye and J.M. Afoutou, 2007. Toxic effects of neem products (Azadirachta indica A. Juss) on Aedes aegypti linnaeus 1762 larvae. Afr. J. Biotechnol., 6: 2846-2854.
Direct Link - Lailatul, K.L., A. Kadarohman and R. Eko, 2010. Biolarvicidal effectivity of ethanol extract from the waste of Akar Wangi (Vetiveria zizanoides) oil destillation towards Aedes aegypti, Culex sp. and Anopheles sundaicus larvae. J. Sains Teknologi Kimia, 1: 59-65.
Direct Link - Isman, M.B., 2006. Botanical insecticides, deterrents and repellents in modern agriculture and an increasingly regulated world. Annu. Rev. Entomol., 51: 45-66.
CrossRefPubMedDirect Link - Zhang, L., X. Feng, D. Ma, J. Yang, H. Jiang, Y. Zhang and W. He, 2013. Brusatol isolated from Brucea javanica (L.) Merr. induces apoptotic death of insect cell lines. Pesticide Biochem. Physiol., 107: 18-24.
CrossRefDirect Link - Syahputra, E., 2008. [Bioactivity of Brucea javanica fruit as nabatical insectiside for agricultural insect pest]. Bul. Littro, 19: 56-67.
Direct Link - Lina, E.C., Dadang, S. Manuwoto, G. Syahbirin and D. Prijono, 2013. Synergistic action of mixed extracts of Brucea javanica (Simaroubaceae), Piper aduncum (Piperaceae) and Tephrosia vogelii (Leguminosae) against cabbage head caterpillar, Crocidolomia pavonana. J. Biopesticides, 6: 77-83.
Direct Link - Lina, E.C., Arnetti, D. Prijono and Dadang, 2010. Insecticidal activity of Brucea javanica L. Merr againts cabbage pests Crocidolomia pavonana (Lepidoptera: Crambidae). Manggaro, 11: 36-39.
Direct Link - Sharma, A., S. Kumar and P. Tripathi, 2015. Impact of Achyranthes aspera leaf and stem extracts on the survival, morphology and behaviour of an Indian strain of dengue vector, Aedes aegypti L. (Diptera: Culicidae). J. Mosquito Res., Vol. 5.
CrossRef - van Meerloo, J., G.J.L. Kaspers and J. Cloos, 2011. Cell Sensitivity Assays: The MTT Assay. In: Cancer Cell Culture, Cree, I.A. (Ed.), Humana Press, Totowa, New Jersey, ISBN: 978-1-61779-079-9, pp: 237-245.
CrossRefDirect Link - Rueda, L.M., K.J. Patel, R.C. Axtell and R.E. Stinner, 1990. Temperature-dependent development and survival rates of Culex quinquefasciatus and Aedes aegypti (Diptera: Culicidae). J. Med. Entomol., 27: 892-898.
CrossRefDirect Link - Barrera, R., M. Amador and G.G. Clark, 2006. Ecological factors influencing Aedes aegypti (Diptera: Culicidae) productivity in artificial containers in Salinas, Puerto Rico. J. Med. Entomol., 43: 484-492.
Direct Link - Choochote, W., U. Chaithong, K. Kamsuk, E. Rattanachanpichai and A. Jitpakdi et al., 2006. Adulticidal activity against Stegomyia aegypti (Diptera: Culicidae) of three Piper spp. Rev. Inst. Med. Trop. S. Paulo, 48: 33-37.
CrossRefDirect Link - Dharmagadda, V.S.S., S.N. Naik, P.K. Mittal and P. Vasudevan, 2005. Larvicidal activity of Tagetes patula essential oil against three mosquito species. Bioresour. Technol., 96: 1235-1240.
CrossRefDirect Link - Chaithong, U., W. Choochote, K. Kamsuk, A. Jitpakdi and P. Tippawangkosol et al., 2006. Larvicidal effect of pepper plants on Aedes aegypti (L.) (Diptera: Culicidae). J. Vector Ecol., 31: 138-144.
CrossRefDirect Link - Kumar, S., R. Warikoo and N. Wahab, 2010. Larvicidal potential of ethanolic extracts of dried fruits of three species of peppercorns against different instars of an indian strain of dengue fever mosquito, Aedes aegypti L. (Diptera: Culicidae). Parasitol. Res., 107: 901-907.
CrossRefPubMedDirect Link - Warikoo, R. and S. Kumar, 2013. Impact of Argemone mexicana extracts on the cidal, morphological and behavioral response of dengue vector, Aedes aegypti L. (Diptera: Culicidae). Parasitol. Res., 112: 3477-3484.
CrossRefDirect Link - Sanjaya, Y. and T. Safaria, 2006. [Toxicity of poison from spider Nephila sp. to Aedes aegypti L. larvae]. J. Biodiversitas, 7: 191-194.
Direct Link - Boublik, Y., P. Saint-Aguet, A. Lougarre, M. Arnaud, F. Villatte, S. Estrada-Mondaca and D. Fournier, 2002. Acetylcholinesterase engineering for detection of insecticide residues. Protein Eng., 15: 43-50.
CrossRefDirect Link - Kim, I.H., R. Suzuki, Y. Hitotsuyanagi and K. Takeya, 2003. Three novel quassinoids, javanicolides A and B and javanicoside A, from seeds of Brucea javanica. Tetrahedron, 59: 9985-9989.
CrossRefDirect Link - Kringer, R., 2010. Handbook of Pesticide Toxicology. Academic Press, California, United States, ISBN-13: 9780080922010, Pages: 2000.
Direct Link - Govindachari, T.R., G.N.K. Kumari, G. Gopalakrishnan, G. Suresh, S.D. Wesley and T. Sreelatha, 2001. Insect antifeedant and growth regulating activities of quassinoids from Samadera indica. Fitoterapia, 72: 568-571.
CrossRefDirect Link - Uthai, U., P. Rattanapreechachai and L. Chowanadisai, 2011. Bioassay and effective concentration of Temephos against Aedes aegypti larvae and the adverse effect upon indigenous predators: Toxorhynchites splendens and Micronecta sp. Asia J. Public Health, 2: 67-77.
Direct Link