
Salah Eldin I. Osman
Department of Zoology, Faculty of Science, Alexandria University, 21511 Alexandria, Egypt
Mohamed H. Swidan
Department of Biology and Geology, Faculty of Education, Alexandria University, 21526 Alexandria, Egypt
Dalia A. Kheirallah
Department of Zoology, Faculty of Science, Alexandria University, 21511 Alexandria, Egypt
Fatma E. Nour
Department of Biology and Geology, Faculty of Education, Alexandria University, 21526 Alexandria, Egypt
Journal of Biological Sciences
Year: 2016 | Volume: 16 | Issue: 3 | Page No.: 93-101







ABSTRACT
Trogoderma granarium is one of the world’s most feared stored-product pests. This study has been conducted to evaluate the histological effects of caraway essential oil and its major monoterpenoid, carvone as well as pyriproxyfen, a juvenile hormone analogue on T. granarium treated as 4th instar larvae. Severe effects were found in the midgut of 4th instar larvae treated with 0.5 mL diluted caraway oil, 2 mL diluted carvone for 2 days. The mid gut and ovarioles of female adults resulted from 4th instar larvae of T. granarium treated with the same concentrations showed also characteristic variations. The histological changes in the midgut and integument of 4th instar larvae treated with 0.05 mL pyriproxyfen for 5 days as well as midgut, integument of delayed 6th instar larvae and ovarioles of female resulted from these treatments after 20 days showed great destruction.
PDF Abstract XML References Citation
Received: January 16, 2016;
Accepted: February 07, 2016;
Published: March 15, 2016
How to cite this article
Salah Eldin I. Osman, Mohamed H. Swidan, Dalia A. Kheirallah and Fatma E. Nour, 2016. Histological Effects of Essential Oils, Their Monoterpenoids and Insect Growth Regulators on Midgut, Integument of Larvae and Ovaries of Khapra Beetle, Trogoderma granarium Everts. Journal of Biological Sciences, 16: 93-101.
DOI: 10.3923/jbs.2016.93.101
URL: https://scialert.net/abstract/?doi=jbs.2016.93.101
DOI: 10.3923/jbs.2016.93.101
URL: https://scialert.net/abstract/?doi=jbs.2016.93.101
INTRODUCTION
Pest insects affect food output directly by reducing the quality and quantity of the crop producted or indirectly by serving as vectors of plant diseases. Insect pests often cause extensive damage to stored grains and their products and this may amount to 5-10% in the temperate zone and 20-30% in the tropical and semitropical environments (Tripathi et al., 2001). The measures to control pest infestation in grain and dry food products depend upon the continuing application of gaseous and liquid insecticides (Ren et al., 2008) which pose possible health hazards to warm-blooded animals, risk of environmental contamination, development of genetically resistant strains of insects and increasing costs of application (Zia et al., 2013). Plants may provide potential alternatives to currently used insect-control agents, because they constitute a rich source of bioactive chemicals (Zia et al., 2013). The benign properties of essential oils permit their use even in sensitive areas, such as schools, restaurants, hospitals and homes because of their lipophilic nature facilitates and their interference with basic metabolic, biochemical, physiological and behavioral functions of insects (Nishimura, 2001). They may also affect some biological parameters such as growth rate, life span and reproduction. Different types of aromatic plant preparations, such as powders, solvent extracts, essential oils and whole plants have been investigated for their insecticidal activity and their action as repellents (Plarre et al., 1997), anti-feedants (Tunc et al., 2000; Lee et al., 2003; Rozman et al., 2007; Ahmed, 2010) and insect growth regulators (Prakash and Rao, 1997) they inhibit the reproduction, growth (development) and possess ovicidal effects in insects (Papachristos and Stamopoulos, 2002). The essential oils are rich in monoterpenes and cause death of insects by inhibiting acetyl cholinesterase activity in nervous system (Tayoub et al., 2012). Monoterpenoid compounds have been considered as potential pest control agents because of their biological activity as ovicides, fumigants (have comparable fumigant action to that of methyl bromide) and contact toxicants (Tsao et al., 1995). Another promising way is the use of Insect Growth Regulators (IGRs) a class of bio rational compounds that mimic insect-produced hormones and potentially reduces pest populations through endocrine disruption. The IGRs are divided into Juvenoids or anti-juvenoids, which mainly affect larval metamorphosis by mimicking juvenile hormone, ecdysteroids, which affect molting and Chitin Synthesis Inhibitors (CSIs), which interfere with cuticle formation (Smet et al., 1990; Das, 2013). The main types of insect growth regulators used commercially are juvenile hormone analogues and chitin synthesis inhibitors (Parrella and Murphy, 1998). Insects have only a few cells that have to undergo division, a process known to be highly susceptible to damage by exposure to insecticides, some of these cells are placed in the epithelium of the midgut, where they renew cells responsible for digestion and absorption of the products of digestion. Division of epithelial cells occurs as new cells grow in to replace old worn-out cells in a very metabolically active tissue. Several applications of essential oils, monoterpenoids and IGRS exhibited histopathological changes in the integument, mid gut and ovaries of insects (Ahmed, 2010; Magdia et al., 2009; Nathan et al., 2008; Ghoneim et al., 2008; Abdullah, 2009; El-Bokl et al., 2010; Bakr et al., 2010).
MATERIALS AND METHODS
Insect culture: Culture of T. granarium was reared at the laboratory for 10 generations before used in the experiments, to get rid of any possible pesticidal contamination. Culture and all test insects were maintained through the experiments under controlled condition of 32°C and 60% R.H. The larvae were reared in 1 L jars on a diet containing crushed wheat. Diet was sterilized in an oven at 60°C for half an hour before using in the tests and kept in the refrigerator at 4°C.
Two days old 4th instar larvae have been used in this study. The newly emerged adults were placed for three days on crushed wheat as an ovipositional site for continuation of the culture and incubated at the same conditions.
Source of essential oil, monoterpenoid and insect growth regulator: Caraway (Carum carvii L.) oil was purchased from phatrade, pharaonic Trade and Distillation. Monoterpenoid Carvone, ((R)-CARVONE, 98%) was purchased from Sigma-Aldrich products (Egyptian International center for Import-EICI). The essential oil was analyzed by gas chromatography with mass spectrum (GC-MS) at Ministry of Trade and Industry, Chemistry Administration, contains 91.05% carvone (Major component) and 8.95% limonene. Pyriproxyfen is a juvenile hormone analogue (commercial name: Admiral) was purchased from Nanjing Essence fine-chemical Co., China.
Histological technique for examination of midgut, integument of larvae and ovaries: Ten 4th instar larvae obtained from each 50 larvae cultures on wheat grain mixed with 0.50 mL diluted caraway oil, 2 mL diluted carvone for 48 h and also 5 larvae from control. The remaining larvae were left in the incubator at the rearing conditions until adult emergence. Newly emerged female adults from treatments and control were collected and all the specimens narcotized by ethyl alcohol. Ten 4th instar larvae were also collected from 50 larvae culture on wheat grain mixed with 0.05 mL of stock pyriproxyfen for 5 days, then after 20 days another delayed 6th instar larvae and the four emerged females as well as parallel control individuals were also taken.
All larvae and adults were decapitated and their last abdominal segments were removed and fixed in freshly prepared alcoholic bouin fixative for 24 h. The specimens were washed well with 70% ethyl alcohol and larvae preserved in 70% ethyl alcohol. The larvae and adults were dehydrated completely in an ascending ethyl alcohol series (70, 80, 90, 96 and 100%) 1/2 h for each and adults stored in terpineol to soften the cuticle.
All the specimens cleared in xylene and were impregnated in hot paraffin: Xylol dish and three wax dishes (wax Ι, wax ΙΙ and wax ΙΙΙ) in 50-52°C oven for 1/2 h each, respectively. Embedding was made in hot paraffin wax by using standard plastic cups. The wax block was so solidified by putting it in cold water. The blocks were then stuck to the holder of the rotary microtome, transverse sections were cut at five micron thickness. The ribbons of the sections were floated on slides coated by smear of egg albumin using drops of hot water. The preparations were kept on hot plate at 40°C to spread the wax ribbons. After complete evaporation of water in a drying oven (for at least 24 h), the slides were dipped in xylol for 3-5 min and in a descending series of ethyl alcohol each for 2 min (100, 90, 80 and 70%). Thereafter the slides were dipped in distilled water and stained in hematoxylin for 30-45 min the slides were transferred to tape water for 2 min and then to distilled water. The slides were then stained in 1% eosin as a counter stain for 5-10 sec and rapidly were rinsed in an ascending alcohol series (70, 80, 90 and 96) and put in absolute alcohol for 10 min to remove any water residues and passed into two changes of xylol for 10-15 min/each. Finally the slides mounted in Canada balsam and covered with cover glass and were dried at 40°C for a day. The sections were examined and photographed by a light microscope.
RESULTS
Severe effects were found in mid-gut of the larvae treated with diluted caraway oil; the epithelial cells were densely (measured 79 μm in compare to control 53 μm), the regenerative cells were fewer and the muscular layers were relatively thicker than the control (Fig. 1a, b). The mid gut of the larvae treated with 2 mL diluted carvone showed also severe effects. The epithelium (measured 34 μm) possessed deeply stained nuclei, the muscular layer was thicker and the regenerative cells were not pronounced as those in the control (Fig. 1a, c).
In the 4th instar larvae treated with 0.05 mL pyriproxyfen for 5 days the epithelium was much thickened, epithelial cells were destroyed, large vacuoles were found between the epithelium and the muscular layer, regenerative cells were separated from each other at the base of the epithelium and the muscular layer become thicker than that of the control (Fig. 2a, b).
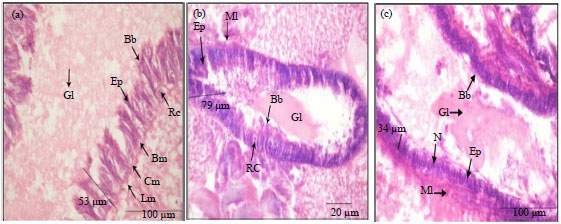 | |
| Fig. 1(a-c): | (a) T.S. in the mid gut of control 4th instar larva of T. granarium, Ep: Epithelium, Rc: Regenerative cells, Bm: Basement membrane, Bb: Brush border, Gl: Gut lumen, Cm: Circular muscle layer, Lm: Longitudinal muscle layer, (b) T.S. in the mid gut of 4th larva of T. granarium treated with 0.50 mL diluted caraway oil, Ep: Elongation of epithelial cells, Rc: Regenerative cells, Bb: Brush border, Ml: Thick muscular layer, Gl: Gut lumen and (c) T.S. in the mid gut of 4th instar larva of T. granarium treated with 2 mL diluted carvone, Ep: Shortened of epithelium, N: Darkly stained nuclei, Bb: Brush border, Ml: Thick muscular layer, Gl: Gut lumen |
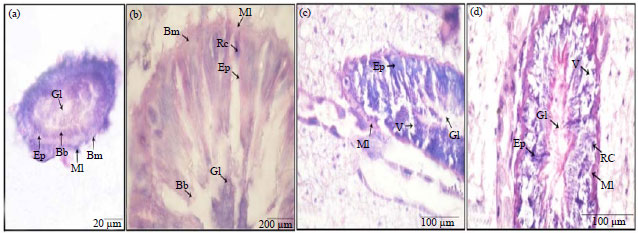 | |
Fig. 2(a-d): | (a) T.S. in mid gut of untreated 6th instar larva of T. granarium, Ep: Epithelial cells, Rc: Regenerative cells, Bb: Brush border, Gl: Gut lumen, Bm: Basement membrane, Ml: Muscle layer, (b) T.S. in mid gut of 6th instar larva of T. granarium treated with 0.05 mL pyriproxyfen for 5 days, Ep: Destructed epithelium, V: Large vacuoles, Ml: Broken muscular layer, Gl: Gut lumen, (c) T.S. in mid gut of untreated late 6th instar larva of T. granarium, Ep: Epithelial cells, Bb: Brush border, Gl: Gut lumen, Bm: Basement membrane, Ml: Muscle layer and (d) T.S. in mid gut of delayed 6th instar larva of T. granarium treated with 0.05 mL pyriproxyfen after 20 days, Ep: Torn epithelial cells, V: Large vacuoles, Rc: Separated regenerative cells, Ml: Thick muscle layer, Gl: Gut lumen |
Severe effects were also found in the mid gut of delayed 6th instar larvae due to treatment with 0.05 mL pyriproxyfen for 20 days (Fig. 2c, d), great destruction in the epithelium, disturbance of epithelial cells, large vacuoles were present and the muscular layer was broken in some places. Regenerative cells could not been identified at the base of the epithelial cells due to the severe destruction of the epithelium, in compare to that of the control.
The cells of hypodermis showed necrological features and blackend (Fig. 3b, d) the cuticular layer appeared brown and no differentiation between exocuticle and endocuticle could be observed (Fig. 3b) in compare to those of the control (Fig. 3a). There was no sign of new cuticle and the old cuticle was separated from the hypodermis (Fig. 3d) in compare to those of control (Fig. 3c).
The mid gut of adults resulted from 4th instar larvae of T. granarium treated with diluted caraway oil showed characteristic variations. The epithelial layer was more elongated; it measures 77 μm in compare to control 14 μm, the lumen appeared narrow, the muscular layer was obviously thicker and the regenerative cells were fewer singly situated at the base of the epithelium (Fig. 4a, b). This lesser number of the regenerative cells could be a result of toxic effect of caraway oil on the insect’s vital activity. It could be concluded that the treatment with caraway oil cause contraction in the mid gut, leading to thickening of the epithelium and muscular layer and narrowing of the gut lumen.
The mid gut of adults resulted from 4th instar larvae treated with diluted carvone showed that the epithelium was columnar measures 42 μm, the nuclei were darkly stained, the regenerative cells were fewer and the muscular layer was thicker than that of the control (Fig. 4a, c).
In the mid gut of females resulted from 4th instar larvae of T. granarium treated with 0.05 mL pyriproxyfen the epithelium was darkened, comparatively thick and showed numerous vacuoles, Regenerative Cells (RC) are located at the base; some of them occupied destroyed spaces in the epithelium and the muscular coat was relatively thick than that of control (Fig. 4a, d).
The ovarioles were affected, due to the treatment with 0.50 mL diluted caraway oil. The germarium and the follicular epithelium of developing oocytes of the ovarioles showed faint nuclei (Fig. 5c, d). This may be due to lesser content of DNA in compare to those of the control (Fig. 5a, b). The basophilic affinity of the nuclei of germarium cells and the follicular epithelium of developing oocytes of the ovarioles was not affected by the treatment with 2 mL diluted carvone (Fig. 5e, f).
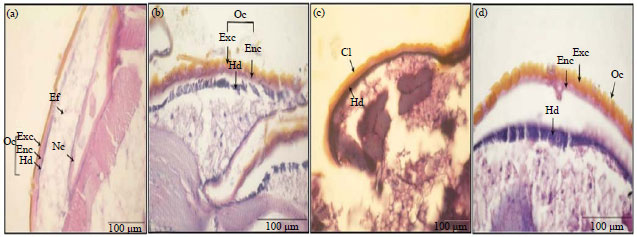 | |
| Fig. 3(a-d): | (a) L.S. of untreated 6th instar larva of T. granarium showing the old and new cuticle, Hd: Hypodermis, Enc: Endocuticle, Exc: Exocuticle, Oc: Old cuticle, Ef: Ecdysal fluid, Nc: New cuticle, (b) L.S. of 6th instar larva of T. granarium treated with 0.05 mL pyriproxyfen showing dead integument, Hd: Dead hypodermis, Cl: Brown cuticular layer, (c) L.S. of untreated late 6th instar larva of T. granarium showing the integument, Hd: Hypodermis, Exc: Exocuticle, Enc: Endocticle, Oc: Old cuticle and (d) L.S. of delayed 6th instar larva of T. granarium treated with 0.05 mL pyriproxyfen after 20 days showing the old cuticle, Oc: Separated old cuticle, Hd: Black hypodermis, Enc: Endocuticle, Exc: Exocuticle |
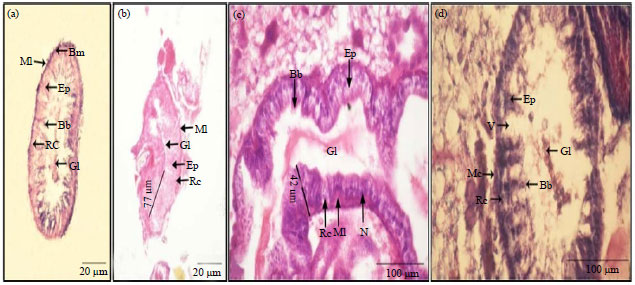 | |
| Fig. 4(a-d): | (a) T.S. in mid gut of untreated female adult of T. granarium, Ep: epithelial cells, Rc: Regenerative celle, Bm: Basement membrane, Bb: Brush border, Gl: Gut lumen, Ml: Muscle layer, (b) T.S. in the mid gut of female adult of T. granarium treated as 4th instar larvae with 0.50 mL diluted caraway oil. Ep: Elongation of epithelial cells, Gl: Narrow gut lumen, Ml: Thick muscle layer, Rc: Few regenerative cells, (c) T.S. in the mid gut of female adult of T. granarium treated as 4th instar larvae with 2 mL diluted carvone. Ep: Columnar epithelial cells, N: Darkly stained nuclei, Rc: Few regenerative cells, Bb: Brush border, Ml: Thick muscle layer, Gl: Gut lumen and (d) T.S. in mid gut of female adult of T. granarium resulted from 4th instar larvae treated with 0.05 mL pyriproxyfen. Ep: Thick epithelium, V: Numerous vacuoles, Rc: Regenerative cells, Mc: Thick muscular coat, Bb: Brush border, Gl: Gut lumen |
The developing oocytes appeared to contain little amount of nutritive material (vitelline) due to treatment with 0.05 mL pyriproxyfen (Fig. 6a, b) and the nuclei of follicular epithelium of the developing oocytes were faintly stained in compare with those of the control (Fig. 5a, b).
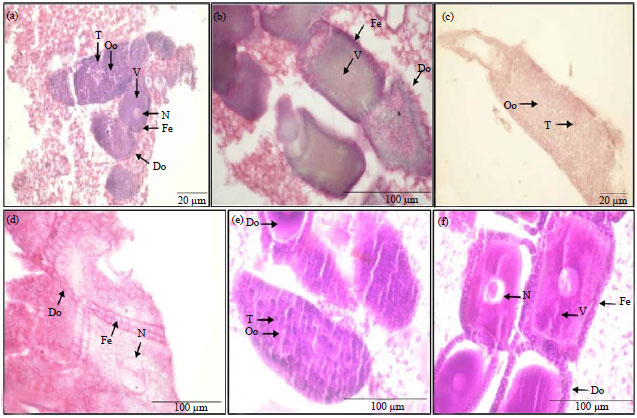 | |
| Fig. 5(a-f): | (a) L.S. in the germarium of an ovariole in control female adult of T. granarium, Oo: Oogonia, T: Trophocytes, Do: Developing oocyte, Fe: Follicular epithelium of developing oocytes, V: Vitelline, N: Nucleus, (b) L.S: Showing developing oocytes in two ovarioles in control female adult of T. granarium, Do: Developing oocytes, V: Vitelline, Fe: Follicular epithelium, (c) L.S. in the germarium of an ovariole in female adult of T. granarium treated as 4th instar larvae with 0.50 mL diluted caraway oil, Oo: Oogonia, T: Trophocytes, (d) T.S. in the ovary of female T. granarium passing through four developing oocytes in female adult of T. granarium treated as 4th instar larvae with 0.50 mL diluted caraway oil, Do: Developing oocyte, Fe: Follicular epithelium nuclei faintly stained, N: Nucleus, (e) Section in three germaria of female adult of T. granarium treated as 4th instar larvae with 2 mL diluted carvone. Oo: Oogonia, T: Trophocytes, Do: Developing oocyte and (f) L.S. through two ovarioles showing developing oocytes of adult female of T. granarium treated as 4th instar larvae with 2 mL diluted carvone, Do: Developing oocyte, V: Vitelline, Fe: Follicular epithelium, N: Nucleus |
DISCUSSION
The present study described the histological changes in the mid gut of the larvae treated as 4th instar larvae with 0.50 mL diluted caraway oil for 2 days, the severe effect observed in the mid gut of the larvae treated with 2 mL diluted carvone and the characteristic variations in the mid gut of adults resulted from treated 4th instar larvae with diluted caraway oil and diluted carvone.
In the mid gut of larvae and adults the regenerative cells were found to be the most susceptible to essential oil and their monoterpenoid, damage to them resulted in the total disruption of the epithelium by preventing the replacement of the functional epithelial cells exhausted by secretory activity (Szczepanik and Ignatowicz, 1998). Our observations coincided with the findings of Abdullah (2009), who observed histopathological lesions, dissolve of nuclei and destroyed of cells of epithelial cells, in 2 days old larvae of R. ferrugineus poisoned by two bio pesticides, Boxus chinensis oil and precocene ΙΙ. Also Ahmed (2010) found an abnormalities in the mid gut and developing oocytes of female of T. granarium treated as 4th instar larvae after two days of treatment with seven essential oil concentrations, especially those prevent new generation. The results also are in agreement with Sharaby et al. (2012) who observed severe effects on the alimentary canal and fat bodies of 1st nymphal instar of H. littoralis after treatment with LC50 concentrations of three essential oils, (garlic, mint and eucalyptus), these effects comprise destruction of epithelial cells, microvilli and the peritrophic membrane were curled and ruptured than those of control group.
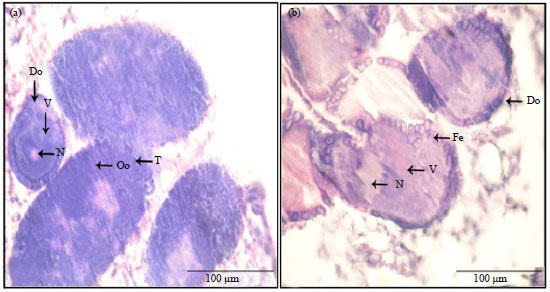 | |
| Fig. 6(a-b): | (a) T.S. showing three germaria and one young developing oocyte of female adult of T. granarium resulted from 4th instar larvae treated with 0.05 mL pyriproxyfen. Oo: Oogonia, T: Trophocytes, Do: Developing oocyte, V: Vitelline, N: Nucleus and (b) T.S. showing developing oocytes of female adult of T. granarium resulted from 4th instar larvae treated with 0.05 mL pyriproxyfen. Fe: Nuclei of follicular epithelium faintly stained, Do: Developing oocyte, V: Poor vitelline, N: Nucleus |
The ovarioles appears to be affected, due to treatment with 0.5 mL diluted caraway oil than in the control while ovarioles appear to be not affected by the treatment with 2 mL diluted cavone. The results come in agreement with Sharaby et al. (2012) who found that the oil of garlic, mint and eucalyptus inhibited oogenesis and egg laying of adults resulting from treating 1st nymphal instars of H. littoralis.
Our observation also revealed histological changes in the 4th instar larvae treated with 0.05 mL pyriproxyfen for 5 days as well as delayed 6th instar larvae and female resulted from these treatments after 20 days. Severe effects were found in the midgut, integument of 6th instar larvae in compare to control and also adults resulted from treated 4th instar larvae showed great destruction. Similar histopathological observations were reported by many investigators Srivastava and Srivastava (1986) who studied histological effects of synthetic juvenile hormone SJH on the midgut cells of the normal and abnormal larval and pupal stages of S. littoralis. Treatment inhibited regeneration of midgut epithelial cells. Ghoneim et al. (2008) assessed histopathological effects of tebufenozide (RH-5992) on the integument and the midgut of last instar nymphs of S. gregaria forsk, detachment of the cuticle from epidermis and undistinguishable epicuticle appeared after treatments. Tebufenozide caused loss in normal architecture of midgut epithelial cells, with dwarf and deformed microvilli. Also, Goncu and Parlak (2011) studied the effects of fenoxycarb on midgut of 5th instar larvae of Bombyx mori with two different doses applied topically. They observed degeneration of larval midgut cells.
As for gonads, the developing oocytes appeared to contain little amount of vitelline and the nuclei of follicular epithelium were faintly stained than those of control. This result find support by Bakr et al. (2010) who tested histopathological changes in the testes on male adults of S. gregaria treated as one day old 5th nymphal instar with LC50 of consult and lufox. The testicular follicles showed damage in zones of reduction and degeneration and necrosis in spermatids and spermatozoa with two compounds.
When insecticides affect the nervous system there are associated effects on the neurosecretory cells which may impair the biological activity of the insect by suppressing the release of the allatotrophic hormone from the neurosecretory cells and thereby affecting the reproductive system of insects (Samaranayaka, 1978). The neurosecretory cells of central nervous system regulate oocyte growth by controlling protein production and by their secretions acting as precursors to stimulate production of the corpus allatum hormone (Mordue, 1965). Reduction of the sexual function may result in part from inhibition of the biosynthesis of DNA (Mitlin et al., 1977).
ACKNOWLEDGMENT
The study was partially funded by Alexandria University. The authors are thankful to Zoology Department, Faculty of science.
REFERENCES
- Tripathi, A.K., V. Prajapati, K.K. Aggarwal and S. Kumar, 2001. Toxicity, feeding deterrence and effect of activity of 1,8,-Cineole from Artemisia annua on progeny production of Tribolium castaneum (Coleoptera: Tenebrionidae). J. Econ. Entomol., 94: 979-983.
CrossRefDirect Link - Ren, Y.L., D. Mahon, J. van Someren Graver and M. Head, 2008. Fumigation trial on direct application of liquid carbonyl sulphide to wheat in a 2500 t concrete silo. J. Stored Prod. Res., 44: 115-125.
CrossRefDirect Link - Zia, S., M. Sagheer, A. Razaq, A. Mahboob, K. Mehmood and Z. Haider, 2013. Comparative bioefficacy of different citrus peel extracts as grain protectant against Callosobruchus chinensis, Trogoderma granarium and Tribolium castaneum. World Applied Sci. J., 21: 1760-1769.
Direct Link - Plarre, R., M. Poschko, S. Prozell, A. Frank, R. Wohlgemuth and J.K. Phillips, 1997. Effects of oil of cloves and citronellol, two commercially available repellents, against the webbing clothes moth Tineola bisselliella Hum. (Lepidoptera: Tineidae). Anzeiger Schadlingskunde Pflanzenschutz Umweltschutz, 70: 45-50.
CrossRefDirect Link - Tunc, I., B.M. Berger, F. Erler and F. Dagli, 2000. Ovicidal activity of essential oils from five plants against two stored-product insects. J. Stored Prod. Res., 36: 161-168.
CrossRef - Lee, S., C.J. Peterson and J.R. Coats, 2003. Fumigation toxicity of monoterpenoids to several stored product insects. J. Stored Prod. Res., 39: 77-85.
CrossRefDirect Link - Rozman, V., I. Kalinovic and Z. Korunic, 2007. Toxicity of naturally occurring compounds of Lamiaceae and Lauraceae to three stored-product insects. J. Stored Prod. Res., 43: 349-355.
CrossRefDirect Link - Papachristos, D.P. and D.C. Stamopoulos, 2002. Repellent, toxic and reproduction inhibitory effects of essential oil vapours on Acanthoscelides obtectus (Say) (Coleoptera: Bruchidae). J. Stored Prod. Res., 38: 117-128.
CrossRefDirect Link - Tayoub, G., A.A. Alnaser and I. Ghanem, 2012. Toxicity of two essential oils from Eucalyptus globulus Labail and Origanum syriacum L. on Larvae of Khapra beetle. Int. J. Med. Arom. Plants, 2: 240-245.
Direct Link - Tsao, R., S. Lee, P.J. Rice, C. Jensen and J.R. Coats, 1995. Monoterpenoids and their Synthetic Derivatives as Leads for New Insect Control Agents. In: Synthesis and Chemistry of Agrochemicals, Baker, D.R., J.G. Fenyes and G.S. Basarab (Eds.). American Chemical Society, Washington, DC., USA., pp: 312-324.
- Smet, H., M. Rans and A. de Loof, 1990. Comparative effectiveness of insect growth regulators with juvenile hormone, anti-juvenile hormone and chitin synthesis inhibiting activity against several stored food insect pests. Proceedings of the 5th International Working Conference on Stored-Product Protection, September 9-14, 1990, Bordeaux, France, pp: 649-658.
- Das, G., 2013. Inhibitory effect of buprofezin on the progeny of rice weevil, Sitophilus oryzae L. (Coleoptera: Curculionidae). Biofertilizers Biopesticides, Vol. 4.
CrossRefDirect Link - Magdia, A.M.H., M.M.S. Alm El-din and E.A.H. El-Akhdar, 2009. The histological and histochemical changes in the gonads of the cotton leaf worm Spodoptera littoralis (Boisd). Isotope Radiat. Res., 41: 1465-1484.
Direct Link - Nathan, S.S., M.Y. Choi, H.Y. Seo, C.H. Paik, K. Kalaivani and J.D. Kim, 2008. Effect of azadirachtin on acetylcholinesterase (AChE) activity and histology of the brown planthopper Nilaparvata lugens (Stal). Ecotoxicol. Environ. Saf., 70: 244-250.
CrossRefPubMedDirect Link - Ghoneim, K.S., R.F. Bakr, M.A. Tanani, A.G. Al Dali and A.S. Bream, 2008. An ultrastructural study on the desert locust (Schistocerca gregaria) as affected by Tebufenozide (RH-5992). Int. J. Agric. Biol., 10: 479-780.
Direct Link - Abdullah, M.A.R., 2009. Toxicological and histopathological studies of Boxus chinensis oil and precocene II on larvae of the red palm weevil Rynchophorus ferrugineus (Oliver)(Coleoptera: Curculionidae). Egypt. Acad. J. Biol. Sci., 2: 45-54.
Direct Link - El-Bokl, M.M., R.F.A. Bakr, H.L. El-Gammal and M.Z. Mahmoud, 2010. Biological and histopathological effects of some insecticidal agents against red palm weevil Rhynchophorus ferrugineus. Egypt. Acad. J. Biol. Sci., 1: 7-22.
Direct Link - Bakr, R.F.A., Mohammed, M.I., A.M. El-Gammal and N.M. Mahdy, 2010. Histopathological change in the testis of the desert locust Schistocerca gregaria (Forskal) induced by the IGR Consult and Lufox. Egypt. Acad. J. Biol. Sci., 1: 23-28.
Direct Link - Sharaby, A., S.A. Montasser, Y.A. Mahmoud and S.A. Ibrahim, 2012. Natural plant essential oils for controlling the grasshopper (Heteracris littoralis) and their pathological effects on the alimentary canal. Ecologia Balkanica, 4: 39-52.
Direct Link - Srivastava, U.C. and U.S. Srivastava, 1986. The effect of a Juvenoid on the acid phosphatase activity in the midgut epithelium of Spodoptera litura (Fabr.). Proc. Indian Natl. Sci. Acad., B52: 723-727.
Direct Link - Goncu, E. and O. Parlak, 2011. The influence of juvenile hormone analogue, fenoxycarb on the midgut remodeling in Bombyx mori (L., 1758) (Lepidoptera: Bombycidae) during larval-pupal metamorphosis. Turkiye Entomoloji Dergisi, 35: 179-194.
Direct Link - Mordue, W., 1965. Neuro-endocrine factors in the control of cyte production in Tenebrio molitor L. J. Insect Physiol., 11: 617-629.
CrossRefDirect Link - Mitlin, N., G. Wiygul and J.W. Haynes, 1977. Inhibition of DNA synthesis in boll weevils (Anthonomus grandis boheman) sterilized by dimilin. Pestic. Biochem. Physiol., 7: 559-563.
CrossRefDirect Link