
Abdalla M. Abdelatif
Department of Physiology, Faculty of Veterinary Medicine, University of Khartoum, Sudan
Mariam Y. Ibrahim
Faculty of Medicine, Omdurman Islamic University, Sudan
Mohamed E. Elnageeb
Department of Physiology, Faculty of Veterinary Medicine, University of Khartoum, Sudan
Journal of Biological Sciences
Year: 2013 | Volume: 13 | Issue: 4 | Page No.: 196-206







ABSTRACT
The type of dietary fat and thermal environment may influence the metabolic profile and insulin sensitivity in normal and diabetic conditions. The objective of this study was to evaluate the effects of supplementation with anhydrous milk fat, ghee (FS) and season (summer vs winter) on blood metabolites and Body Weight (BW) in alloxan-diabetic and non-diabetic rabbits. In each season, twenty rabbits were assigned to 4 groups of 5 animals each. Two groups were rendered diabetic using alloxan monohydrate injection. Two groups, a normal and a diabetic received orally 2.5 g kg-1 b.wt. of ghee daily for 8 weeks. Blood samples collected weekly were used for determination of insulin and blood metabolite concentrations. During summer, insulin (INS) level for the diabetic and non-diabetic-fat supplemented (FS) rabbits was higher than the non-diabetic rabbits fed Lucerne. Diabetic FS rabbits had slightly higher INS level than diabetic group fed Lucerne. Glucose (GL) level was significantly higher in diabetic groups and GL level of FS diabetic rabbits was significantly higher than respective values of diabetic group fed Lucerne only. GL level of diabetic groups was significantly higher in summer. In both seasons, serum total lipids (TL) was significantly higher in diabetic than non-diabetic groups. TL was significantly higher in FS diabetic group in summer. FS increased triglyceride (TG) levels in diabetic and non-diabetic animals, TG level was significantly lower in winter in non-diabetic groups of rabbits. In both seasons, the FS diabetic significantly higher serum cholesterol (CHOL) level than the diabetic control and non-diabetic groups of rabbits. The CHO L evel in FS non-diabetic rabbits was significantly higher during winter. In both seasons, serum urea (UR) level was significantly higher in diabetic groups. Diabetic groups showed significantly higher UR during winter compared to summer values. Creatinine (CR) level of FS nondiabetic group was higher during summer. During summer, diabetic groups showed significantly lower mean BW than non-diabetic groups, while with FS, non-diabetic group showed significantly greater mean BW than the other groups. Diabetic FS group had significantly lower mean BW during winter. The findings have implications in pathophysiology and nutritional management of DM.
PDF Abstract XML References Citation
Received: March 04, 2013;
Accepted: April 03, 2013;
Published: July 10, 2013
How to cite this article
Abdalla M. Abdelatif, Mariam Y. Ibrahim and Mohamed E. Elnageeb, 2013. Evaluation of Effects of Milk Lipid (Ghee) Supplementation and Thermal Environment on the Metabolic Profile of Alloxan-diabetic Rabbits (Oryctolagus cuniculus). Journal of Biological Sciences, 13: 196-206.
DOI: 10.3923/jbs.2013.196.206
URL: https://scialert.net/abstract/?doi=jbs.2013.196.206
DOI: 10.3923/jbs.2013.196.206
URL: https://scialert.net/abstract/?doi=jbs.2013.196.206
INTRODUCTION
Diabetes Mellitus (DM) is a common metabolic disease with alterations in carbohydrate, protein and lipid metabolism as a consequence of defects in insulin secretion, insulin action or both (American Diabetes Association, 2010). Recently, the incidence of DM has increased drastically in both developed and underdeveloped countries. In Sudan, DM is currently emerging as an important public health problem, especially in urban areas. Previous survey indicated higher prevalence of diabetes in adult population with marked spatial distribution (Elbagir et al., 1996). The diet composition may have significant effect on insulin sensitivity as well as risk and complications of diabetes (Mann, 2006). Medical nutrition therapy is usually adopted as an effective measure to control hyperglycaemia and dyslipidaemia in diabetic patients (Franz, 2004). The dietary recommendations for diabetic patients seems to be similar for insulin dependent DM (IDDM) and non-insulin dependent DM (NIDDM).
The composition and quantity of dietary fat induce metabolic disorders in normal and diabetic subjects. Chronic consumption of high saturated fat diets deteriorates the metabolic profile and impairs glucose tolerance and increases the risk of type 2 diabetes (Feskens et al., 1995). Also prolonged feeding of high-fat diet in rats, induced hyperinsulinaemia and insulin resistance (Ghatta and Ramarao, 2004). Prolonged feeding of high-fat diet in rats (Woods et al., 2003; Winzell and Ahren, 2004) was associated with increased body weight, hyperinsulinaemia and insulin resistance. Jen et al. (1991) reported that alloxan-diabetic rabbits developed hypercholesterolaemia and hypertriglyceridaemia in response to cholesterol feeding. Yin et al. (2002) noted that a diet high in saturated fat and sucrose impaired glucose regulation in rabbits and resulted in high serum levels of total cholesterol and triglyceride.
The NIDDM which is associated with tissue resistant insulin, is strongly linked with obesity; however, the mechanism by which increased adiposity causes insulin resistance has not been clarified. Yin et al. (2004) reported that in miniature pigs, feeding a high-fat and high-sucrose diet resulted in increase in the size of adipocytes and plasma GL levels and insulin resistance. Dysfunctions of lipoprotein lipase (LPL) were associated with dyslipidaemia, obesity and insulin resistance. An increase in LPL activity in transgenic rabbits induced marked lowering of plasma triglycerides (TG) and non-esterified fatty acids, however, the basal levels of glucose and insulin (INS) were not altered (Kitajima et al., 2004). In Golden Syrian hamsters, increased saturated fat intake was associated with disturbance of homeostasis, severe hypertriglyceridaemia and increase in creatinine (CR) concentrations related mainly to nephropathy (Popov et al., 2003).
The anhydrous milk fat (ghee), is an important dietary fat used for culinary purposes including dressing of foods and cooking. Ghee also has medicinal uses including treatment of allergy, skin and respiratory diseases (Sharma et al., 2010). In Sudan, ghee is used for topping of sauce and it is also drunk for therapeutic purposes (Dirar, 1993). The conjugated linoleic acid in milk fat proved to have anticarcinogenic (Pariza, 1991) and antiatherogenic properties (Lee et al., 1994). Ghee constitutes a source of lipid nutrients, fat-soluble vitamins and essential fatty acids (Chand et al., 1986). In milk fat, the short chain and medium chain fatty acids of milk fat are readily digested, absorbed and oxidized by carnitine pathway (Bansal and Kansal, 1996). Kumar et al. (1999) reported lower concentrations of serum triglycerides, total cholesterol and Low Density Lipoprotein (LDL) cholesterol in rats fed anhydrous milk fat.
Epidemiological studies revealed seasonal pattern in the incidence of DM; the highest numbers of cases were diagnosed during the cool months in winter (Nystorm et al., 1990; Elbagir et al., 1996). However, gestational DM risk diagnosis was significantly higher in summer (Vasileiou et al., 2012). Seasonal variation was also reported in glycosylated haemoglobin (HbA1c) (Asplund, 1997; Carney et al., 2000), insulin sensitivity (Berglund et al., 2012) and lipid profile (Kamezaki et al., 2010; Bardini et al., 2012).
Chemical induction of DM in laboratory animals proved to be useful in elucidating metabolic and pathophysiological changes. The rabbit model is widely adopted in experimental and clinical physiology. The present study was designed to evaluate the effects of dietary ghee supplementation and season on blood metabolites in normal and alloxan-diabetic rabbits.
MATERIALS AND METHODS
Animals: Clinically healthy adult male rabbits (Oryctolagus cuniculus) were used. Animals were procured from a private farm and were aged 8-10 months at the commencement of the experiment.
Housing and management: The rabbits were individually caged in a well ventilated animal facility with natural photoperiod at the Department of Physiology. Animals were allowed to adapt to the housing conditions and experimental procedures for two weeks and had free access to fresh Lucerne and clean tap water. Clinical examination was performed before and during the course of the experiment. Animals were given prophylactic anthelmintic injection (Ivomec: 0.02 mL kg-1 b.wt., Alpha Laboratories Ltd., India) and antibacterial injection (Oxytetracycline: 7.5 mg kg-1 b.wt., Alpha Laboratories Ltd, India).
Feeding: The animals were given fresh Lucerne (Medicago sativa) (Dry matter basis: CP 17.5%; ME 7.8 MJ kg-1). Commercial anhydrous milk fat (ghee) purchased from the local market was used for treated groups of animals. The proximate composition of ghee is triglyceride 98.6%, cholesterol 161.7 mg/100 g, phospholipids 10.4 mg/100 g (Kumar et al., 1999).
Climatic conditions: The ambient temperature (Ta) and Relative Humidity (RH) measurements were obtained from the Meteorological Unit located about 500 m from the experimental site. The data for the experimental periods under summer and winter conditions are depicted in Table 1.
Induction of diabetes mellitus (DM): The treated groups of rabbits were made diabetic by a single intravenous injection of 150 mg kg-1 alloxan monohydrate (Sigma, St. Louis, MO) dissolved in 0.9% NaCl.
| Table 1: | The ambient temperature (Ta) and relative humidity (RH) prevailing during the experimental period |
 | |
Therapeutic measures were adopted to secure the survival of animals by administration of glucose to tide over initial hypoglycaemic phase and injection of insulin (INS) during the acute phase of hyperglycaemia. The Glucose Tolerance Test (GTT) was performed to ensure the diabetic status. Animals with plasma GL levels higher than 200 mg dL-1 were regarded as diabetic and used in the study. Animals developed diabetes mostly on the third day following injection of alloxan.
Collection of blood samples: Blood samples (4 mL) were withdrawn from the jugular vein by plastic syringes. One milliliter of blood was kept in a test tube containing sodium fluoride and after centrifugation, the plasma was used for glucose determination. The rest of the blood sample was left at room temperature for 3 h and then centrifuged (Hettich-Germany) at 3000 rpm for 15 min. Haemolysis-free serum samples were stored at -20°C for subsequent analysis.
Analysis of blood metabolites: The plasma glucose (GL) concentration was determined by colorimetric method (Trinder, 1969) using a commercial kit (Spinreact, S.A. Spain). Serum urea (UR) concentration was determined by enzymatic colorimetric test (Berthot) using a kit (Spinreact, S.A., Spain). Serum creatinine (CR) was determined by colorimetric extraction method (Owen et al., 1954). Serum Total Lipids (TL)were estimated according to the method of Frings et al. (1972) measuring the coloured complex with a phosphate ester of vanillin. Serum triglyceride (TG) concentration was determined using the enzymatic method (Liner Chemical, Spain). Serum cholesterol (CHOL) concentration was determined by enzymatic colorimetric method (Spinreact S.A., Spain).
Serum insulin: Serum insulin (INS) concentration was determined by radioimmunoassay kit (Beijing Institute of Atomic Research, Beijing, China).
Experimental procedure: Twenty rabbits were randomly assigned to 4 groups of 5 animals each. Two groups were redered diabetic using alloxan monohydrate as previously explained. Animals in two groups, a normal and a diabetic group were supplemented with milk fat, ghee (FS). Each animal received orally 3 g (2.5 g kg-1 b.wt.) daily. All animals were given free access to fresh Lucerne and tap water. The rabbits were subjected to the dietary experimental protocol for 8 weeks under typical summer and winter climatic conditions. During the experimental period, Body Weights (BW) of rabbits were recorded weekly. Blood samples were collected weekly at 7:00 a.m. The samples were used for the determination of the concentrations of plasma glucose, serum metabolites and insulin.
Data analysis: For each group of animals, the mean values were computed during the course of experimental period. The data are presented as Mean±Standard Deviation (SD). The Analysis of Variance (ANOVA) (SAS, 1998) and Duncans Multiple Range Test (DMRT) were used to evaluate the effects of diabetes, supplementation with fat (FS) and season on the parameters investigated. Differences were considered to be statistically significant at p<0.05.
RESULTS
The results of the effects of milk fat supplementation (FS) on serum insulin, plasma glucose, serum metabolites and Body Weight (BW) in summer and winter are shown in Table 2 and 3, respectively. The effects of season on parameters investigated are depicted in histograms.
Serum insulin (INS): During summer (Table 2), the diabetic groups of rabbits had significantly (p<0.001) lower values of serum INS concentration (7.56-7.79 Mu L-1) compared to the values measured for non-diabetic groups (15.47-16.46 Mu L-1). However, the INS concentration for FS groups was slightly higher than values of groups fed Lucerne only. Winter data (Table 3) indicate that diabetic groups of rabbits maintained significantly (p<0.001) lower insulin levels compared to the respective values obtained for the non-diabetic groups. However, FS did not influence the INS level. Figure 1 shows that seasonal change in thermal environment had no significant effect on serum INS concentration in diabetic and non-diabetic groups of rabbits.
Plasma glucose (GL): During summer (Table 2), the diabetic FS group of rabbits exhibited significantly (p<0.001) higher GL concentration than the respective value of diabetic group fed Lucerne only. The non-diabetic groups of rabbits maintained steady GL level during the experimental period. During winter (Table 3), GL level was highly significantly (p<0.001) higher in the diabetic groups of rabbits compared to non-diabetic rabbits. Figure 2 shows that in both diabetic groups of rabbits, GL level was significantly (p<0.001) higher in summer compared to respective winter value. There was no change related to season in non-diabetic groups of rabbits.
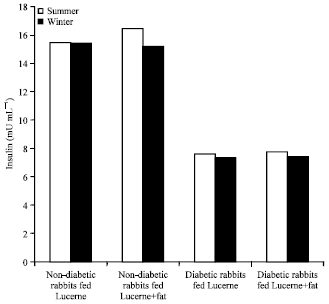 | |
| Fig. 1: | Effect of supplementation with milk fat (Ghee) and season on serum insulin concentration in diabetic and non-diabetic rabbits |
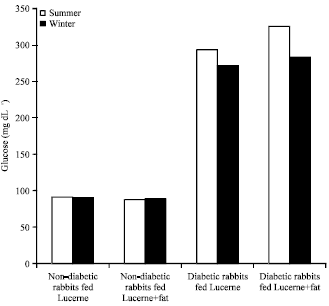 | |
| Fig. 2: | Effect of supplementation with milk fat (Ghee) and season on plasma glucose concentration in diabetic and non-diabetic rabbits |
Serum total lipids (TL): During summer (Table 2), diabetic groups of rabbits had significantly (p<0.01) higher serum TL concentration compared to the non-diabetic groups of rabbits. However, the diabetic FS group of rabbits had significantly (p<0.05) higher TL concentration compared to the diabetic group fed lucerne. In non-diabetic groups, the FS group had slightly higher value. During winter (Table 3), diabetic groups of rabbits had significantly (p<0.01) higher serum TL than the respective values of non-diabetic groups.
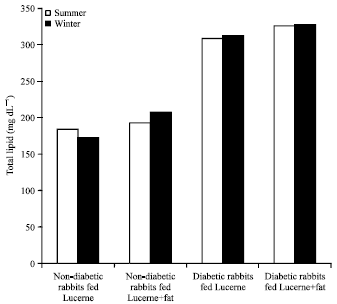 | |
| Fig. 3: | Effect of supplementation with milk fat (Ghee) and season on serum total lipid concentration in diabetic and non-diabetic rabbits |
However, the diabetic FS group of rabbits had significantly (p<0.05) higher serum TL concentration compared to the diabetic group fed Lucerne. Figure 3 shows that seasonal change in thermal environment had no significant effect on TL in diabetic and non-diabetic groups of rabbits.
Serum triglycerides (TG): During summer (Table 2), diabetic FS rabbits had significantly (p<0.05) higher triglyceride concentration. During winter (Table 3), FS had significant (p<0.05) effect on TG concentration in diabetic groups of rabbits, it was higher in the diabetic group of rabbits supplemented with fat. For non-diabetic groups, the FS group had a higher value. Figure 4 shows that TG concentration was significantly (p<0.05) higher during summer compared to the respective winter value in non-diabetic groups of rabbits.
Serum cholesterol (CHOL): During summer (Table 2), the diabetic FS group had significantly (p<0.001) higher CHOL than the diabetic control and non-diabetic groups of rabbits. Diabetic group of rabbits fed Lucerne only had significantly higher CHOL level than the corresponding values of the non-diabetic groups of rabbits. However, serum CHOL concentration was significantly (p<0.05) higher in non-diabetic group supplemented with fat than non-diabetic group fed Lucerne only. During winter (Table 3), the diabetic FS group of rabbits had significantly (p<0.05) higher serum CHOL than the diabetic group fed Lucerne and non-diabetic groups of rabbits. However, there was no significant difference in CHOL concentration of the diabetic group fed Lucerne and the non-diabetic FS group fed Lucerne.
| Table 2: | Effect of supplementation with milk fat (Ghee) on serum insulin, plasma glucose, serum metabolites and body weight (BW) in diabetic and non-diabetic rabbits during summer |
 | |
| Mean values within the same row bearing different superscripts are significantly different, LS: Level of significance, *p<0.05, ***p<001, NS: Not significant | |
| Table 3: | Effect of supplementation with milk fat (Ghee) on serum insulin, plasma glucose, serum metabolites and body weight (BW) in diabetic and non-diabetic rabbits during winter |
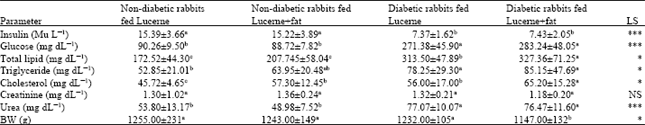 | |
| Mean values within the same row bearing different superscripts are significantly different, LS: Level of significance, *p<0.05, ***p<001, NS: Not significant | |
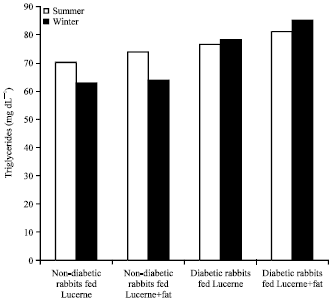 | |
| Fig. 4: | Effect of supplementation with milk fat (Ghee) and season on serum triglycerides concentration in diabetic and non-diabetic rabbits |
The non-diabetic group fed Lucerne exhibited significantly (p<0.05) lower serum CHOL concentration. Figure 5 shows that serum CHOL concentration in non-diabetic FS rabbits was significantly (p<0.05) higher during winter than summer. However, season had no significant effect on serum cholesterol in other groups.
Serum creatinine (CR): During summer (Table 2), the non-diabetic group of rabbits fed Lucerne had slightly higher CR value.
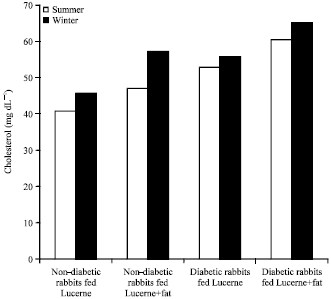 | |
| Fig. 5: | Effect of supplementation with milk fat (Ghee) and season on serum cholesterol concentration in diabetic and non-diabetic rabbits |
During winter (Table 3), the effect of FS on serum CR concentration was not significant in diabetic and non-diabetic rabbits. However, the diabetic FS group of rabbits had lower value. Figure 6 shows that serum CR concentration was significantly (p<0.05) higher during summer in non-diabetic group fed lucerne and diabetic FS group.
Serum urea (UR): During summer (Table 2), the diabetic groups of rabbits maintained significantly (p<0.001) higher serum UR concentration compared to non-diabetic groups of rabbits.
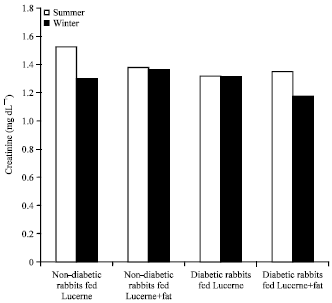 | |
| Fig. 6: | Effect of supplementation with milk fat (Ghee) and season on serum creatinine concentration in diabetic and non-diabetic rabbits |
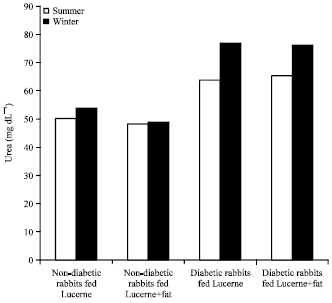 | |
| Fig. 7: | Effect of supplementation with milk fat (Ghee) and season on serum urea concentration in diabetic and non-diabetic rabbits |
During winter (Table 3), the diabetic groups of rabbits maintained significantly (p<0.001) higher UR concentration compared to non-diabetic groups of rabbits. There was no significant variation in UR concentration between diabetic groups. Figure 7 shows that the diabetic FS groups of rabbit and their control showed significantly (p<0.001) higher UR concentration during winter compared to the respective values summer values.
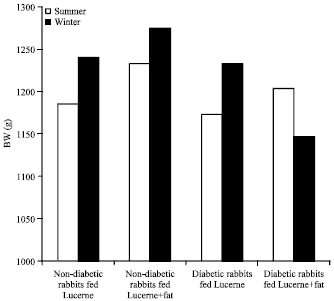 | |
| Fig. 8: | Effect of supplementation with milk fat Ghee) and season on body weight (BW) in diabetic and non-diabetic rabbits |
Body weight (BW): During summer (Table 2), the diabetic groups had significantly (p<0.05) lower mean BW compared to non-diabetic groups. The non-diabetic SF group of rabbits showed significantly (p<0.05) higher mean BW compared to the other experimental groups of rabbits. During winter (Table 3), the non-diabetic group of rabbits maintained significantly (p<0.05) higher BW compared to the values obtained for the other groups. However, the diabetic FS group of rabbits showed significantly (p<0.05) lower mean BW. Figure 8 shows that the diabetic and non-diabetic group of rabbits fed Lucerne and the non-diabetic FS group had significantly (p<0.01) higher BW during winter compared to the respective summer values. The diabetic FS group had significantly (p<0.01) higher BW during summer.
DISCUSSION
This study aimed to evaluate the effects of supplementation with animal fat (FS) and seasonal change in tropical environment on Body Weight (BW), insulin (INS) level and blood metabolites in alloxan-diabetic and non-diabetic rabbits. The current study used the milk fat (ghee) which generally makes appreciable contribution to saturated fat and cholesterol intake in humans.
The lower INS levels in the diabetic rabbits in the present study is clearly the result of induction of experimental diabetes with alloxan. The tendency for increase in serum INS level in non-diabetic FS rabbits during summer (Fig. 1) in the present study is in agreement with Yin et al. (2002) who indicated that the INS level in rabbits fed a diet high in saturated fat and sucrose increased significantly over the first 3 months of experimental period. However, reductions of 50% in circulating INS level have been reported in high fat fed rats and it has been suggested that these diets impair signal transduction mechanism in pancreatic B-cells to reduce INS secretion (Kim et al., 1996). Yin et al. (2003) indicated that there were no significant differences in fasting serum INS levels among rabbits fed high fat and high sucrose diet and their control.
The hyperglycaemia observed in alloxan-treated animals (Table 2, 3) is clearly attributed to INS deficiency. When INS is deficient (absolute or relative), hyperglycaemia develops as a result of increased gluconeogenesis, accelerated glycogenolysis and impaired GL utilization by peripheral tissues (DeFronzo et al., 1994). Increased hepatic GL production is attributed to the high availability of gluconeogenic amino acids precursors associated with enhanced proteolysis (Felig and Wahren, 1971) and lactate (induced by increased muscle glycogenolysis) and glycerol (resulting from increased lipolysis) (Exton, 1987). Conversion of glucose-6-phosphate to glucose is catalyzed by hepatic glucose-6-phosphatase, which is stimulated by an increase in secretion of catabolic hormones and decreased insulin levels (Kitabchi et al., 2001).
The FS diabetic rabbits exhibited significantly higher GL levels than the respective values of diabetic groups fed Lucerne only during summer and winter (Table 2, 3). Ikemoto et al. (1995) indicated that in mice, high dietary fat (safflower oil) induced defective glycaemic control and hyperglycaemia. The higher level of hyperglycaemia in FS diabetic rabbits may be related to inhibition of muscle GL uptake and oxidation. Jequier (1998) indicated that GL metabolism may be inhibited at two important steps. First, the increase in cytoplasmic citrate concentration inhibits phosphofructokinase, which results in an increased glucose-6-phosphate concentration; as a consequence, hexokinase inhibition and impairment of GL uptake. Second, inhibition of pyruvate dehydrogenase reduces the entry of pyruvate into oxidative metabolism and thus contributes to inhibition of GL oxidation. Additionally, the expression of the INS-regulatable GL transporter GLUT-4 (mRNA and protein) was dramatically reduced in skeletal muscle after feeding a high-lard-based diet to mice (Kahn and Pedersen, 1993; Zierath et al., 1997). Reduced INS sensitivity in muscle of high-fat-fed animals was attributed partly to changes in INS signaling, which may ultimately produce a lower GL uptake. Yin et al. (2003) induced increase in GL level by feeding rabbits with high-fat-high-sucrose diet.
The current results indicate that serum INS and GL levels were influenced by season (Fig. 1, 2). There was a tendency for higher INS level in summer in normal FS and in diabetic groups. However, Donahoo et al. (2000) demonstrated that in normal weight humans, GL level was not significantly different between summer and winter.
The data indicate that generally alloxan-treated groups showed higher serum TL and TG levels compared to non-diabetic groups (Table 2, 3). However, FS increased significantly TG levels irrespective of season. The increase in TG is clearly associated with INS deficiency which results in mobilization of fatty acids from adipose tissue. As blood GL is not utilized by tissues, the fatty acids from adipose tissue are mobilized for energy purpose and excess fatty acids which accumulate in the liver are converted to TG (Shih et al., 1997). Suryawanshi et al. (2006) indicated that the blood TG levels change according to the glycaemic control.
High levels of TG may be related to obesity, increase in calorie intake and lack of muscular exercise in diabetic patients (Yagi, 1987). High fat meals exacerbate postprandial hypertriglyceridaemia. Impaired INS action in type Π diabetes may result in loss of suppression of lipolysis in adipose tissue. This leads to an increased release of free fatty acids into the portal circulation and consequently, increased delivery of FFAs to the liver. The effect of this process is increased production of triglycerides by the liver and a decreased production of HDL-C. In addition, there is impaired clearance of TG from the circulation. The resulting hypertriglyceridaemia alters the activity of other enzymes, which leads to the formation of small dense LDL particles and increased catabolism of HDL (Valabhji and Elkeles, 2003). In contrast to the present results, Zulet et al. (1999) indicated that in rats, hypercholesterolaemia was accompanied by a decrease in TG. This reduction in circulating TG could be associated with an inadequate synthesis as Rawashdeh (2002) reported that TG level was reduced after a period of ghee consumption in humans.
Elevated serum TG was noted in rabbits fed high fat/high sucrose diets (Kloeze and Abdellatif, 1975; Yin et al., 2002, 2003). The mobilization of TG seems to be a consequence of INS deficiency. However, in some rabbits the blood lipids remained at low levels during the whole experimental period in spite of an existing INS deficiency and hyperglycaemia. Furthermore, Shafique et al. (2002) in studies on alloxan-induced diabetic rabbit, noted increased serum TG in diabetic placebo group.
Diabetic groups of rabbits fed lucerne only had significantly higher CHOL level than the values of the non-diabetic groups of rabbits (Table 2, 3). The current results are in general agreement with previous studies (Kloeze and Abdellatif, 1975; Khushk et al., 2010) which reported considerable increase in serum CHOL in diabetic rabbits. The diabetic and non-diabetic rabbits had higher serum CHOL levels with FS. A positive relationship between the dietary CHOL intake and serum CHOL was reported in hamsters (Cohen et al., 1994). Rawashdeh (2002) reported that ingestion of ghee significantly increased serum total CHOL level in humans. The hypercholesterolaemic effect of ghee may be related to the high amount of short and medium chain fatty acids which may work as an activator for hepatic hydroxyl-methyl-glutarate-CoA reductase, the rate limiting enzyme in CHOL biosynthesis. Short chain fatty acids with two other fatty acids, lauric and myristic acids may induce hypercholesterolaemia (Elson, 1992) and may cause high rate of CHOL absorption (Ide et al., 1979). Myristic acid is the most potent CHOL raising saturated fatty acid in humans (Hajri et al., 1998). However, paradoxically Kumar et al. (2000) reported that dietary ghee supplementation decreased serum CHOL level that was attributed to increase in the secretion of biliary lipids. The authors reported that dietary ghee did not influence the activity of HMG Co A reductase in liver indicating that it had no influence on biosynthesis of CHOL, but increased significantly the bile secretion and thus reduced CHOL levels.
The current results demonstrated seasonal variation in serum lipid levels. The lower TG level in winter for normal rabbits in the current study (Fig. 4) is in line with previous report in humans (Fuller et al., 1974). However, other researchers reported higher TG level in winter (Donahoo et al., 2000; Kamezaki et al., 2010). Serum CHOL level was higher in winter for non-diabetic rabbits (Fig. 5). This result confirms the hypercholerolaemia reported in normal subjects consistently in winter by other workers (Donahoo et al., 2000; Ockene et al., 2004; Kamezaki et al., 2010). The current results reported for the first time seasonal variation in CHOL level in alloxan-induced diabetic animals (Fig. 4). Recently, Bardini et al. (2012) showed seasonal variation in lipid profile of Type 2 diabetic patients. The mechanisms for seasonal patterns remain largely unknown (Gordon et al., 1987; Woodhouse et al., 1993). It has been suggested that changes in relative plasma volume may explain partly the seasonal differences in lipid profile (Ockene et al., 2004). The climatic data depicted in Table 1 indicate that the mean ambient temperature was 32.71°C in summer and 27.50°C in winter. However, Bardini et al. (2012) indicated that in Type 2 diabetic patients, the peak of serum total CHOL during colder months was associated with changes of diet and lifestyle habits.
The UR and CR levels are generally considered as biomarkers of diabetic nephropathy (De Almeida et al., 2012). In the present study, UR level increased significantly in diabetic rabbits but the data does not show marked change in CR level (Table 2, 3). The high UR level could be associated with impaired renal function (Ronco et al., 2010) and may also indicate an increase in tissue catabolism (De Almeida et al., 2012).The higher UR level during winter in diabetic rabbits (Fig. 7) may be related to haemoconcentration and/or an increase in dietary protein intake.
The results indicate that in both seasons, diabetic groups of rabbits had significantly lower Body Weight (BW) than the corresponding mean values in the non-diabetic groups (Table 2, 3). The reported decrease in BW of diabetic rabbits is attributed mainly to breakdown of structural proteins (Franz et al., 2002). The amino acids produced in proteolysis are oxidized because body cells cannot utilize GL for energy (Sekar et al., 2005). Ramesh et al. (2006) reported BW loss in streptozotocin diabetic rats. The increase in BW in non-diabetic FS group of rabbits during summer is in agreement with previous studies in animals fed high fat diet (Murray et al., 1999). Sandu et al. (2005) reported that in mice exposed to high dietary fat, metabolic processes which lead to BW gain are linked to advanced glycosylation end products. Chen et al. (1992) reported that rats fed a high-saturated fat diet consumed more energy and gained more weight compared with animals on two different high-carbohydrate diets. Other researchers indicated that BW in rats were similar in control and hypercholesterolaemic groups, while a marked decrease in abdominal fat was found in rats fed the CHOL and saturated fat enriched diet; this was attributed to reduction in food intake and ketogenic mechanisms (Tomlinson et al., 1992).
We conclude that the metabolic profile of normal and alloxan-diabetic rabbits was influenced by milk fat intake and seasonal climatic change. The seasonal rhythm of serum lipids implies that control of dyslipidaemia and optimal dietary therapy should be adopted in the cold season. Special clinical attention should be payed to hypercholesterolaemic and diabetic subjects consuming diets with high milk fat content.
REFERENCES
- De Almeida, D.A.T., C.P. Barga, E.L.B. Novelli and A.A.H. Fernandes, 2012. Evaluation of lipid profile and oxidative stress in STZ-induced rats treated with antioxidant vitamin. Braz. Arch. Biol. Technol., 55: 527-536.
CrossRefDirect Link - American Diabetes Association, 2010. Diagnosis and classification of diabetes mellitus. Diabetes Care, 33: S62-S69.
CrossRefDirect Link - Asplund, J., 1997. Seasonal variation of HbA1c in adult diabetic patients. Diabetes Care, 20: 234-234.
PubMedDirect Link - Bardini, G., I. Dicembrini, C.M. Rotella and S. Giannini, 2012. Lipids seasonal variability in type 2 diabetes. Metabolism, 61: 1674-1677.
CrossRef - Berglund, L., C. Berne, K. Svardsudd, H. Garmo, H. Melhus and B. Zethelius, 2012. Seasonal variations of insulin sensitivity from a euglycemic insulin clamp in elderly men. Upsala J. Med. Sci., 117: 35-40.
CrossRefPubMedDirect Link - Carney, T.A., S.P. Guy and C.D. Helliwell, 2000. Seasonal variation in HbA1c in patients with type 2 diabetes mellitus. Diabetes Med., 17: 554-555.
CrossRef - Chand, R., S. Sree Kumar, R.A. Srinivasan, V.K. Batish and H. Chander, 1986. Influence of lactic bacteria cells on the oxidative stability of ghee. Milchwissenschaft, 41: 335-336.
Direct Link - Chen, M.T., L.N. Kaufman, T. Spennetta and E. Shrago, 1992. Effects of high fat-feeding to rats on the interrelationship of body weight, plasma insulin and fatty acyl-coenzyme a esters in liver and skeletal muscle. Metabolism, 41: 564-569.
CrossRef - Cohen, B.I., N. Ayyad, T. Mikami, Y. Mikami and E.H. Mosbach, 1994. Effects of dietary fat and fatty acids on sterol balance in hamsters. Lipids, 29: 503-508.
CrossRef - Donahoo, W.T., D.R. Jensen, T.Y. Shepard and R.H. Echel, 2000. Seasonal variation in lipoprotein lipase and plasma lipids in physically active, normal weight humans. J. Clin. Endocrinol. Metab., 85: 3065-3068.
CrossRefDirect Link - Elbagir, M.N., M.A. Eltom, E.M.A. Elmahadi, I.M.S. Kadam and C. Berne, 1996. A population-based study of the prevalence of diabetes and impaired glucose tolerance in adults in Northern Sudan. Diabetes Care, 19: 1126-1128.
CrossRefDirect Link - Exton, J.H., 1987. Mechanisms of hormonal regulation of hepatic glucose metabolism. Diabetes/Metab. Rev., 3: 163-183.
CrossRef - Felig, P. and J. Wahren, 1971. Influence of endogenous insulin secretion on splanchnic glucose and amino acid metabolism in man. J. Clin. Invest., 50: 1702-1711.
CrossRef - Feskens, E.J., S.M. Virtanen, L. Rasanen, J. Tuomlilehto and J. Stengard et al., 1995. Dietary factors determining diabetes and impaired glucose tolerance: A 20-year follow-up of the Finnish and Dutch cohorts of the seven countries study. Diabetes Care, 18: 1104-1112.
CrossRefDirect Link - Franz, M.J., J.P. Bantle, C.A. Beebe, J.D. Brunzell and J.L. Chiasson et al., 2002. Evidence-based nutrition principles and recommendations for the treatment and prevention of diabetes and related complications. Diabetes Care, 25: 148-198.
CrossRefDirect Link - Franz, M.J., 2004. Evidence-based medical nutrition therapy for diabetes. Nutr. Clin. Pract., 19: 137-144.
CrossRefPubMedDirect Link - Frings, C.S., T.W. Fendley, R.T. Dunn and C.A. Queen, 1972. Improved determination of total serum lipids by the sulfo-phospho-vanillin reaction. Clin. Chem., 18: 673-674.
CrossRefPubMedDirect Link - Fuller, J.H., S.L. Grainger, R.J. Jarrett and H. Keen, 1974. Possible seasonal variation of plasma lipids in a healthy population. Clin. Chim. Acta, 52: 305-310.
CrossRef - Ghatta, S. and P. Ramarao, 2004. Increased contractile responses to 5-hydroxytryptamine and Angiotensin II in high fat diet fed rat thoracic aorta. Lipids Health Dis., Vol. 3.
CrossRefDirect Link - Gordon, D.J., D.C. Trost, J. Hyde, F.S. Whaley, P.J. Hannan, D.R. Jacobs Jr. and L.G. Ekelund, 1987. Seasonal cholesterol cycles: The lipid research clinics coronary primary prevention trial placebo group. Circulation, 76: 1224-1231.
CrossRefDirect Link - Hajri, T., P. Khosla, A. Pronczuk and K.C. Hayes, 1998. Myristic acid-rich fat raises plasma LDL by stimulating LDL production without affecting fractional clearance in gerbils fed a cholesterol-free diet. J. Nutr., 128: 477-484.
PubMedDirect Link - Ide, T., T. Tanaka and M. Sugano, 1979. Dietary fat-dependent changes in hepatic cholesterogenesis and the activity of 3-hydroxy-3-methylglutaryl-CoA reductase in fasted-refed rats. J. Nutr., 109: 807-818.
PubMed - Ikemoto, S., K.S. Thompson, M. Takahashi, H. Itakura, M.D. Lane and O. Ezaki, 1995. High fat diet-induced hyperglycemia: Prevention by low level expression of a glucose transporter (GLUT4) minigene in transgenic mice. Proc. Natl. Acad. Sci. USA., 92: 3096-3099.
CrossRef - Jen, K.L., C. Rochon, S.B. Zhong and L. Whitcomb, 1991. Fructose and sucrose feeding during pregnancy and lactation in rats changes maternal and pup fuel metabolism. J. Nutr., 121: 1995-2005.
PubMedDirect Link - Jequier, E., 1998. Effect of lipid oxidation on glucose utilization in humans. Am. J. Clin. Nutr., 67: 527S-530S.
Direct Link - Kahn, B.B. and O. Pedersen, 1993. Suppression of GLUT4 expression in skeletal muscle of rats that are obese from high fat feeding but not from high carbohydrate feeding or genetic obesity. Endocrinology, 132: 13-22.
CrossRefDirect Link - Kamezaki, F., S. Sonoda, Y. Tomotsune, H. Yunaka and Y. Otsuji, 2010. Seasonal variation in serum lipid levels in Japanese workers. J. Atherosclerosis Thrombosis, 17: 638-643.
CrossRefPubMedDirect Link - Khushk, I., M.U. Dahot, S.A. Baloach and M.A. Bhutto, 2010. The evaluation of soybean extracts in alloxan-induced diabetic rabbits. World Applied Sci. J., 8: 22-25.
Direct Link - Kim, J.K., J.K. Wi and J.H. Youn, 1996. Metabolic impairment precedes insulin resistance in skeletal muscle during high fat feeding in rats. Diabetes, 45: 651-658.
Direct Link - Kitabchi, A.E., G.E. Umpierrez, M.B. Murphy, E.J. Barrett, R.A. Kreisberg, J.I. Maline and B.M. Wall, 2001. Management of hyperglycaemic crises in patients with diabetes. Diabetes Care, 24: 131-153.
CrossRefDirect Link - Kitajima, S., M. Morimoto, E. Liu, T. Koike and Y. Higaki et al., 2004. Overexpression of lipoprotein lipase improves insulin resistance induced by a high-fat diet in transgenic rabbits. Diabetologia, 47: 1202-1209.
CrossRefDirect Link - Kloeze, J. and A.M.M. Abdellatif, 1975. Effects of palm-kernel oil and sunflower-seed oil on serum lipids and atherogenesis in alloxan-diabetic rabbits. Atherosclerosis, 22: 349-367.
CrossRefDirect Link - Kumar, M.V., K. Sambaiah and B.R. Lokesh, 2000. Hypocholesterolaemic effect of anhydrous milk fat ghee is mediated by increasing the secretion of biliary lipids. J. Nutr. Biochem., 11: 69-75.
PubMedDirect Link - Kumar, M.V., K. Sambiah and B.R. Lokesh, 1999. Effect of dietary ghee-the anhydrous milk fat, on blood and liver lipids in rats. J. Nutr. Biochem., 10: 96-104.
CrossRefDirect Link - Lee, K.N., D. Kritchevsky and M.W. Pariza, 1994. Conjugated linoleic acid and atherosclerosis in rabbits. Atherosclerosis, 108: 19-25.
CrossRefPubMedDirect Link - Mann, J.I., 2006. Nutrition recommendations for the treatment and prevention of Type 2 diabetes and the metabolic syndrome: An evidence-based review. Nutr. Rev., 64: 422-427.
CrossRefDirect Link - Nystorm, L., G. Dahlquist, M. Rewers and S. Wall, 1990. The Swedish childhood diabetes study. An analysis of the temporal variation in diabetes incidence 1978-1987. Int. J. Epidemiol., 19: 141-146.
CrossRefDirect Link - Ockene, I.S., D.E. Chiriboga, E.J. Stanek, M.G. Harmatz and R. Nicolosi et al., 2004. Seasonal variation in serum cholesterol levels: Treatment implications and possible mechanisms. Arch. Intern. Med., 164: 863-870.
CrossRefDirect Link - Owen, J.A., B. Iggo, F.J. Scandrett and C.P. Stewart, 1954. The determination of creatinine in plasma or serum and in urine; a critical examination. Biochem. J., 58: 426-437.
Direct Link - Pariza, M.W., 1991. CLA, A new cancer inhibitor in dairy products. Bull. Int. Dairy Fed., 257: 29-30.
Direct Link - Ramesh, B., R. Saravanan and K.V. Pugalendi, 2006. Effect of dietary substitution of Groundnut oil on blood glucose, lipid profile and redox status in streptozotocin-diabetic rats. Yale J. Biol. Med., 79: 9-17.
PubMed - Rawashdeh, A.Y.A., 2002. Influences of olive oil and ghee (samen balady) on serum cholesterol of Jordanians. Pak. J. Nutr., 1: 270-275.
CrossRefDirect Link - Ronco, C., S. Grammaticopoulus, M. Rosner, M. De Cal, S. Soni, P. Lentini and P. Piccinni, 2010. Oliguria, creatinine and other biomarkers of acute kidney injury. Contrib. Nephrol., 164: 118-127.
CrossRefDirect Link - Sandu, O., K. Song, W. Cai, F. Zheng, J. Uribarri and H. Vlassara, 2005. Insulin resistance and type 2 diabetes in high-fat-fed mice are linked to high glycotoxin intake. Diabetes, 54: 2314-2319.
CrossRefDirect Link - Sekar, D.S., K. Sivagnanam and S. Subramanian, 2005. Antidiabetic activity of Momordica charantia seeds on streptozotocin induced diabetic rats. Pharmazie, 60: 383-387.
Direct Link - Shafique , M., M. Ahmed, K.M. Fayyaz and S. Hussain, 2002. Beneficial effects of magnesium supplementation in diabetes mellitus-an animal study. Pakistan J. Med. Res., 41: 73-76.
Direct Link - Sharma, H., X. Zhang and C. Dwivedi, 2010. The effect of ghee (clarified butter) on serum lipid levels and microsomal lipid peroxidation. Ayu, 31: 134-140.
CrossRefDirect Link - Shih, K.C., C.F. Kwok, C.M. Hwu, L.C. Hsiao, S.H. Li, Y.F. Liu and L.T. Ho, 1997. Acipimox attenuates hypertriglyceredemia in dyslipidemic non-insulin dependent diabetes mellites patients without perturbation of insulin sensitivity and glycemic control. Diabetic. Res. Clin. Pract., 36: 113-119.
PubMedDirect Link - Suryawanshi, N.P., A.K. Bhutey, A.N. Nagdeote, A.A. Jadhav and G.S. Manoorkar, 2006. Study of lipid peroxide and lipid profile in diabetes mellitus. Ind. J. Clin. Biochem., 21: 126-130.
Direct Link - Tomlinson, K.C., S.M., Gardiner, R.A. Hebden and T. Bennett, 1992. Functional consequences of streptozotocin-induced diabetes mellitus, with particular reference to the cardiovascular system. Pharmacol. Rev., 44: 103-150.
PubMedDirect Link - Trinder, P., 1969. Determination of glucose in blood using glucose oxidase with an alternative oxygen acceptor. Ann. Clin. Biochem., 6: 24-27.
CrossRefDirect Link - Valabhji, J. and R. Elkeles, 2003. Dyslipidaemia in type п diabetes: Epidemiology and biochemistry. Br. J. Diabetes Vasc. Dis., 3: 184-189.
Direct Link - Vasileiou, V., E. Kyratzoglou, A. Athanasiadou, A. Giakoumi, S. Mouzakioti, M. Alevizaki and E. Anastasiou, 2012. Is there a seasonal variation in the incidence of gestational diabetes mellitus. Endocr. Abs., 29: 599-599.
Direct Link - Winzell, M.S. and B. Ahren, 2004. The high-fat diet-fed mouse: A model for studying mechanisms and treatment of impaired glucose tolerance and type 2 diabetes. Diabetes, 53: S215-S219.
CrossRefPubMedDirect Link - Woodhouse, P.R., K.T. Khaw and M. Plummer, 1993. Seasonal variation of serum lipids in an elderly population. Age Ageing, 22: 273-278.
CrossRefDirect Link - Woods, S.C., R.J. Seeley, P.A. Rushing, D. D'Alessio and P. Tso, 2003. A controlled high-fat diet induces an obese syndrome in rats 1. J. Nutr., 133: 1081-1087.
PubMed - Yin, W., Z. Yuan, Z. Wang, B. Yang and Y. Yang, 2002. A diet high in saturated fat and sucrose alters glucoregulation and induces aortic fatty streaks in New Zealand White rabbits. Int. J. Exp. Diab. Res., 3: 179-184.
CrossRefDirect Link - Yin, W., D. Liao, Z. Wang, S. Xi and K. Tsutsumi et al., 2004. NO-1886 inhibits size of adipocytes, suppresses plasma levels of tumor necrosis factor-α and free fatty acids, improves glucose metabolism in high-fat/high-sucrose-fed miniature pigs. Pharmacol. Res., 49: 199-206.
CrossRefDirect Link - Yin, W., Z. Yuan, K. Tsutsumi, Y. Xie and Q. Zhang et al., 2003. A lipoprotein lipase-promoting agent, NO-1886, improves glucose and lipid metabolism in high fat, high sucrose-fed New Zealand white rabbits. Exp. Diabesity Res., 4: 27-34.
CrossRef - Yagi, K., 1987. Lipid peroxides and human diseases. Chem. Physiol. Lipids, 45: 337-351.
CrossRefDirect Link - Zierath, J.R., K.L. Houseknecht, L. Gnudi and B.B. Kahn, 1997. High-fat feeding impairs insulin-stimulated GLUT4 recruitment via an early insulin-signaling defect. Diabetes, 46: 215-223.
Direct Link - Zulet, M.A., A. Barber, H. Garcin, P. Higueret and J.A. Martinez, 1999. Alterations in carbohydrate and lipid metabolism induced by a diet rich in coconut oil and cholesterol in a rat model. J. Am. Coll. Nutr., 18: 36-42.
CrossRefPubMedDirect Link