
Riaz Ahmad
Section of Genetics, Department of Zoology, Faculty of Life Sciences, Aligarh Muslim University, Aligarh-202 002, Uttar Pradesh, India
Rakesh B. Pandey
Section of Genetics, Department of Zoology, Faculty of Life Sciences, Aligarh Muslim University, Aligarh-202 002, Uttar Pradesh, India
S. Hasan Arif
Section of Genetics, Department of Zoology, Faculty of Life Sciences, Aligarh Muslim University, Aligarh-202 002, Uttar Pradesh, India
Nikhat Nabi
Department of Zoology, Amar Singh College, Gogjibagh, Srinagar (J and K), India
Mumtaz Jabeen
Section of Genetics, Department of Zoology, Faculty of Life Sciences, Aligarh Muslim University, Aligarh-202 002, Uttar Pradesh, India
Absar-ul Hasnain
Section of Genetics, Department of Zoology, Faculty of Life Sciences, Aligarh Muslim University, Aligarh-202 002, Uttar Pradesh, India
Journal of Biological Sciences
Year: 2012 | Volume: 12 | Issue: 2 | Page No.: 98-104







ABSTRACT
SDS-PAGE profiles of soluble eye lens nuclei proteins of walking catfish, Clarias batrachus (Linn.) revealed the existence of eight distinct polymorphs along the western region of Uttar Pradesh, India. On the basis of their molecular weights (Mr) and isoelectric points (pI), the polypeptides were identified as β- and γ-crystallins. The crystallins characteristically focused between pI values of 5.2 to 8.7 and had molecular weights of 19 to 34 kDa. Statistical analysis of polymorphs within random catches indicated a latitudinal pattern of fish distribution. We propose that the observed distribution pattern is the consequence of limited terrestrial migration of C. batrachus. Nocturnal migration to nearby water bodies is its outstanding characteristic as an air-breathing and walking silurid, specifically during rainy season. Population substructure is genetically recognizable into three zones: upper, central and lower; out of which the latter two are most inhabited. F-value according to ANOVA was 1.095 indicating that at different locations, polymorphs existed in differing frequencies (p = 0.375). Present results thus suggest that with the exception of the flood-prone locations, selection pressures which have limited the output of this threatened catfish have not disturbed the homogeneity of population structure in the investigated region.
PDF Abstract XML References Citation
Received: December 09, 2011;
Accepted: February 08, 2012;
Published: March 13, 2012
How to cite this article
Riaz Ahmad, Rakesh B. Pandey, S. Hasan Arif, Nikhat Nabi, Mumtaz Jabeen and Absar-ul Hasnain, 2012. Polymorphic β and γ Lens Crystallins Demonstrate Latitudinal Distribution of Threatened Walking Catfish Clarias batrachus (Linn.) Populations in North-western India. Journal of Biological Sciences, 12: 98-104.
DOI: 10.3923/jbs.2012.98.104
URL: https://scialert.net/abstract/?doi=jbs.2012.98.104
DOI: 10.3923/jbs.2012.98.104
URL: https://scialert.net/abstract/?doi=jbs.2012.98.104
INTRODUCTION
Walking or labyrinth catfish Clarias batrachus (Linn.) has a wide distribution in Indian subcontinent, South Asian countries, Indochina, China extending up to Guam (Axelrod et al., 1971). It is an important food fish and considered not only delicious but of medicinal value, as well. Air-breathing by highly branched and vascular labyrinth imparts C. batrachus an exceptional capability of terrestrial migration to closely located water bodies and surviving short spells of water scarcity by aestivation.
C. batrachus used to be an abundant component of catfish catches in the investigated region about three decades ago. This is a region where two major rivers of Indian subcontinent Ganges and Yamuna, a less prominent Gomti and several of their tributaries flow through (Fig. 1). During past two decades, however, the region has witnessed intermittent periods of drought and devastation of perennial habitats of the catfish. The survival and growth aspects of larvae as well as adult C. batrachus and other clariids are not so well documented (Samad et al., 2005; Adekunle, 2011; Khan et al., 2011). The other suspect is twice as fast growing and exotic sister species, the African catfish Clarias gariepinus (Linn.). However, its projected growth potential has been in general negated by the carnivorous feeding habit which can seriously endangers not only C. batrachus, but virtually all other cohabitant species. Since the catfish breeds during late monsoon months, eventually some or all of these factors must have curtailed abundance of C. batrachus, resulting its listing as a threatened species. A recent record classifies C. batrachus as a vanishing species in a far off but water-resource-rich Southern state of Kerala (Binoy, 2010). Therefore, documentation of diversity by identifying suitable genetic markers will be helpful in assessment of the present resources, their better management and rehabilitation of C. batrachus.
We report here polymorphism in eye lens nuclear crystallins of C. batrachus inhabiting North-Western plains of Uttar Pradesh which is discernible by gradient SDS-PAGE profiles. Polymorphism of this magnitude has not so far been reported for teleost lens proteins.
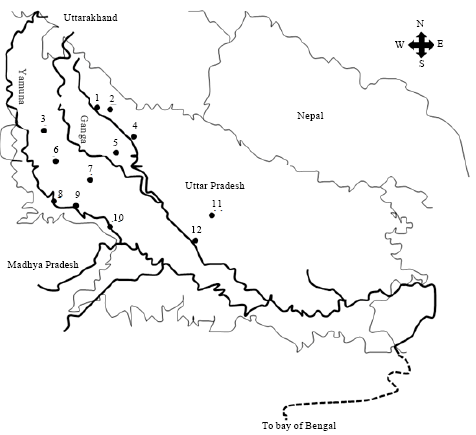 | |
| Fig. 1: | Map of north-western province of Uttar Pradesh, showing sampling locations and the rivers. Samples at each map location represented heterogenous collections from various ponds and ditches. Sample collection locations in the map are: 1: Moradabad, 2: Rampur, 3: Bulandshahr, 4: Bareilly, 5: Badaun, 6: Aligarh, 7: Etah, 8: Agra, 9: Firozabad, 10: Etawah, 11: Lucknow, 12: Kanpur |
There exists a general paucity of literature on genetic variations in fish eye lenses or the nuclei baring some early reports (Barrett and Williams, 1967; Eckroat and Wright, 1969; Smith and Clemens, 1973, 1982; Day and Clayton, 1973; Hasnain et al., 1974). Published evidence has mostly revealed species-specificity of eye lens proteins of teleosts rather than the intraspecies differences (Calhoun and Koenig, 1970; Phan et al., 1985; Suzuki and Phan, 1990; Jamieson and Teixeira, 1991). Taxon-specific eye lens proteins have been identified as multifunctional crystallins (Wistow, 1993) and high resolution IEF further discriminated crystallins in to α, β and γ subtypes (Basaglia, 1989; Basaglia and Di Luca, 1993). Apart from being cost effective, SDS-PAGE has the advantages of resolving polypeptides rather than intact proteins and will dissociate crystallins which might otherwise tend to aggregate (Bon et al., 1968; Srinivas et al., 2011). Our report underlines applicability of SDS-PAGE in elucidating crystallin polymorphism.
MATERIALS AND METHODS
Fish collection and their procurement: During 1998-2005, live samples of Clarias batrachus of 20-30 cm were collected from many sites of North-Western states of Uttar Pradesh (26°-29° latitude and 77½°-81° longitude), India (Fig. 1). Catches from ponds and ditches scattered within a periphery of ~50 km, brought live to city markets (locations shown in the map, Fig. 1), were obtained and brought live to the laboratory. The catfish can be conveniently transported in wet grass due to air-breathing capability. Information about ponds and ditches which were source of samples was recorded from professionals bringing in catches to central locations.
Extraction of eye lens nuclefi proteins: To obtain eye lens nuclei of the fish, cortex was removed by rolling the eye lenses first on dry and then wet analytical grade filter paper (Whatman No. 1). They were washed twice with distilled water and then homogenized in 50 mM Tris-HCl (pH, 7.5) following overnight extraction at 15±1°C with gentle shaking. The homogenate was centrifuged at 10,000 rpm and 4°C for 15 min and clear supernatant was carefully removed.
Protein estimation and polyacrylamide gel electrophoresis: Following protein estimation (Bradford, 1976), it was immediately mixed in appropriate amounts of 4x sample buffer containing SDS and incubated at 100°C for 3 min as per standard protocol (Laemmli, 1970). After keeping overnight at room temperature, they were stored frozen till further analysis. SDS-PAGE was carried out on 15% gels following a modified system in use at this laboratory (Hasnain et al., 1999). After initial screening, representative phenotypes of each lot were run together, visualized by Coomassie Brilliant Blue (CBB) staining and photographed. Densitometric tracing was performed using Scion image software.
Identification of crystallins by isoelectric focusing: Identification of crystallins was carried out by Isoelectric Focusing (IEF) on 5% polyacrylamide gels (M:C = 5:3) using broad range ampholite (pH 3-10). It was performed at 2000 volts for 2 hrs keeping other conditions as per instruction manual of LKB-Pharmacia.
Statistical analysis: The data on the polymorphs was statistically tested through ANOVA using SPSS (Version 11.0) software and phylogenetic inter relationship among them was demonstrated by using Unweighted Pair Group Method with Arithmetic Mean (UPGMA) cluster analysis (Ferguson, 1980).
RESULTS
Eye lenses of Clarias batrachus are quite small and depending on the size of fish used here measured between 1.2-1.8 mm in diameter. To exclude age dependent modifications, a size range of 20-30 cm was chosen which represents fish of about 1 to 1.5 years of age. Central hard core or nucleus after removing soft cortex further reduces the size by about two-third of the lens diameter. Therefore, the amount of soluble fraction has been sufficient mostly for one time analysis. Best resolutions of soluble fraction were obtained in 15% gels. Figure 2a demonstrates the patterns of soluble proteins of eye lens nuclei (Lane 1-4) as well as of whole lenses (Lane 5-8). It was observed that bulk (>90%) of soluble proteins of lens nuclei resolve with molecular weights of 19-34 kDa (Fig. 2b) and pI values of 5.2-8.7 (Fig. 2c) confirming β- and γ-crystallin subtypes. In addition, fluctuating levels of α-crystallins (pI≤5.2) in few of the polymorphs excluded their value as markers. During the present investigations, therefore, bands out of 19-34 kDa range and/or posses pI values below 5.2 or above 8.7 have been ignored. IEF profiles of either of the polymorph demonstrate the presence of many more bands in comparison with the polypeptides resolved by SDS-PAGE. Obviously, several crystallin isoforms (as resolved by IEF) share similar molecular weights (Fig. 2c). Densitometric comparison confirmed that the relative intensity of individual SDS-PAGE polymorph correlates with the intensity (sum of total crystallin bands) of the corresponding IEF profile.
As shown in Fig. 2b, the number of bands in various polymorphs differs from 3-9. Band No. 1, 2, 4, 6, 7, 9, 10, 11 and 13 are frequent in their distribution, while those marked No. 3, 5, 8 and 12 occur, restrictively within the populations of locations other than Moradabad, Etawah, Kanpur and Lucknow. Intraspecies phylogenetic relationship of discovered polymorphs obtained by UPGMA analysis is shown in Fig. 3. It is evident that polymorphs No. 5 and 7 as well as No. 3 and 6 cluster together and become part of bigger clusters constituted by polymorphs No. 8 and 4, respectively. The two big clusters apparently represent two different lineages with the immediate predecessors being polymorphs No. 1 and 2.
DISCUSSION
Published data on population structure of C. batrachus are scarce and somewhat contradictory. While, Islam et al. (2007) found some genetic variability in wild and hatchery populations, Khedkar et al. (2010) reported a lack of genetic diversity even in distant Indian populations. There is thus a need for identifying diverse sets of markers and widening coverage.
Fish eye lens is unique as it maintains lifelong refractivity by continuously recruiting crystallins (Bours, 1980). Ontogenetic and age related changes may also affect electrophoretic profiles of soluble proteins of eye lenses (Zigman and Yulo, 1979; Benz 1980; Smith and Gilman, 1982). Therefore, the information obtained from electrophoretic profiles would greatly depend on the type of recruited proteins (crystallins) their relative concentration, solubility and the techniques employed to resolve the soluble lens proteins. While defining polymorphs of C. batrachus crystallins as genetic, we have attempted to address these concerns: (1): Only fish of adult size were sampled. (2): Soluble lens nuclei proteins were analyzed, since they are more reliable source of genetic information (Smith and Clemens, 1973). (3): Reducing and dissociating conditions of SDS-PAGE resolve aggregates of α-crystallins (Bon et al., 1968; Srinivas et al., 2011). (4): The data were statistically tested to examine the correlation between the observed polymorphism and the population distribution of the fish.
IEF analysis of the selected polymorphs identified α-, β- and γ-crystallins and corresponded with SDS-PAGE profiles stacking as bands of 19-34 kDa (Fig. 2b).
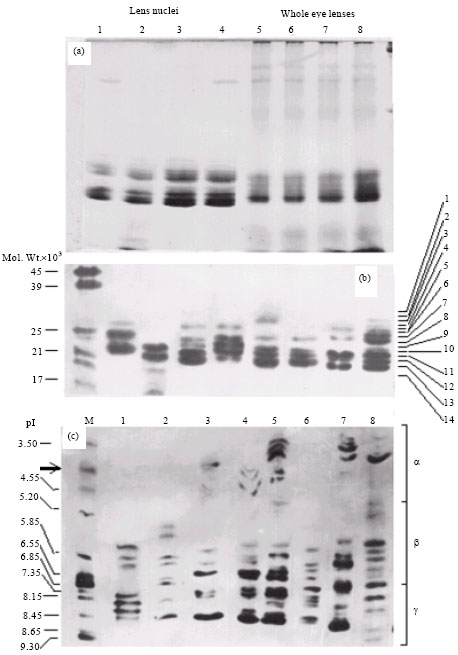 | |
| Fig. 2 (a-c): | Typical SDS-PAGE patterns of soluble proteins of lens nuclei (lanes 1-4) and whole eye lenses (lanes 5-8) of C. batrachus in 15% gels, Eight polymorphs detected in soluble nuclear proteins among populations inhabiting the western belt of Uttar Pradesh and IEF patterns of the eight polymorphs showing multiplicity and types of crystalline isoforms |
Our findings on IEF profiles based subtyping are in agreement with species diagnostic subtypes of some teleosts (Basaglia, 1989) and other vertebrates (Zigler and Sidbury, 1976; Zigman and Yulo, 1979). These bands also stack with pI values of 5.2-8.7 (Fig. 2c) confirming their identity as β- and γ-crystallins (Smith and Gilman, 1982; De Jong and Hendriks, 1986; Piatigorsky and Wistow, 1989; Basaglia and Di Luca, 1993).
We have excluded α-crystallins because of inconsistent quantitative and numerical variations in their electrophoretic/IEF profiles. Alpha-crystallins levels are more likely to fluctuate since they are stress inducible molecular chaperons which keep lens refractive (Fernald, 1990; Basu et al., 2002; Dahlman et al., 2005). They also determine stability of lens proteins of teleosts living at temperature extremes (Kiss et al., 2004) as also the heat stability at high temperatures (Yu et al., 2004). On the other hand, inner core crystallins deposit first in eye lens of teleosts and the sequential deposition proceeds according to an ontogenetic program (Smith and Gilman, 1982).
While cumulatively 14 protein bands were detectable in SDS-PAGE profiles, one or the other of the eight lens nuclei polymorphs of C. batrachus consisted of 3 to 9 crystallin bands or co stacks (Fig. 2b).
 | |
| Fig. 3: | Dendrogram showing the phylogenetic relationship of entire set of eight polymorphs of eye lens nuclei (P-1 to P-8 are polymorphs 1-8) |
Statistical analysis reveals the existence of these polymorphs in three distinct population zones of C. batrachus along this region. Upper Zone (UZ) constituted by locations of Rampur, Moradabad, Bareilly, Bulandshahr, Badaun and Aligarh; Central Zone (CZ) having only Etah and Lower Zone (LZ) comprising Firozabad, Agra, Etawah, Kanpur and Lucknow.
The data presented here demonstrate that distribution of C. batrachus inhabiting seasonal or perennial ponds and ditches, constituting the freshwater non-riverine habitat of the region is not influenced by the riverine barriers of this region. Instead, a latitudinal pattern of distribution is the unique feature, rarely reported for other fish species (Koehn and Rasmussen, 1967). Although, we do not rule out predation by C. gariepinus, our data on crystallin polymorphism in C. batrachus showed no anomaly in distribution pattern justifying such a correlation. The latitudinal distribution of C. batrachus can, however be explained by instinctive nocturnal terrestrial-migration, which is the characteristic of C. batrachus as a walking catfish with accessory air-breathing support. Heterogeneity existed in perennial flood-prone location of Etah and Etawah, while homogeneity (p<0.05) characterized the populations inhabiting majority of other locations. Thus, while homogeneity among populations of C. batrachus is due to characteristic land-walk, large scale intermixing is due to floods and hence different levels of gene flow.
Based on genetic identity two big clusters can be recognized in the dendrogram (Fig. 3). One of them is formed by polymorph No. 4 along with No. 3 and 6, which make up minicluster within this cluster. The other one is constituted by polymorph No. 8, No. 5 and 7 with the latter two forming a minicluster again. The data demonstrates that while retaining some of the characteristics of their immediate progenitors (No. 2 or No. 1), polymorphs of both clusters are typically limited to some latitudinal zone in their distribution. The two main lineages exhibit typical patterns of distribution as polymorph No. 2 is restricted to only LZ whereas polymorph No. 1 to UZ. Out of the two big clusters where polymorphs No. 4 and 8 are the next immediate progenitors, polymorph No. 8 inhabits mainly LZ while No. 4 is nearly uniformly distributed in both of the zones. The minicluster constituted by No. 3 and 6 occupy mainly UZ whereas that formed by polymorphs No. 5 and 7 are uniformly distributed in both of the zones. Apparently, the factors responsible for latitudinal distribution of different polymorphs are yet to favor the equilibrated propagation of the minicluster constituted by the former two polymorphs confined to UZ or it represent a less viable lineage. In spite of selection pressures that are limiting the output of C. batrachus in the investigated region, there is still a definite pattern of distribution wherein different populations appear inclined to reach homogeneity.
CONCLUSIONS
Polymorphism has been detected in SDS-PAGE profiles of eye lens nuclei crystallins of air-breathing walking catfish Clarias batrachus. The detected eight polymorphs were exclusively composed of β and γ-crystallins, which characteristically focus with pI values of 5.2-8.7 and stack as polypeptides of 19-34 kDa in SDS-PAGE profiles. Statistical analyses of random samples collected from a wide area of western Uttar Pradesh (India) suggest a latitudinal pattern of distribution of the polymorphs that we attribute to land migration of the walking catfish to closely located water bodies. We conclude that despite being a threatened catfish, operational selection pressure seems to be ineffective in disturbing the population equilibrium of C. batrachus.
ACKNOWLEDGMENTS
This study was mainly supported by a grant from the Indian Council of Agricultural Research (ICAR), New Delhi, India to the last author (AH). Authors are obliged to Chairman, Department of Zoology, Aligarh Muslim University for necessary facilities. We also thankfully acknowledge Prof. A. Qayyum Siddiqui for a critical revision of the manuscript.
REFERENCES
- Barrett, I. and A. Williams, 1967. Soluble lens proteins of some scombroid fishes. Copeia, 2: 468-471.
CrossRef - Basaglia, F., 1989. Study of the soluble lens proteins from fifteen sparidae species. Comp. Biochem. Physiol., 94: 625-629.
CrossRef - Basaglia, F. and D. di Luca, 1993. A comparative study of vertebrate eye lens crystallins using isoelectric focusing and densitometry. Comp. Biochem. Physiol., 106: 575-586.
PubMed - Basu, N., A.E. Todgham, P.A. Ackerman, M.R. Bibeau, K. Nakano, P.M. Schulte and G.K. Iwama, 2002. Heat shock protein genes and their functional significance in fish. Gene, 295: 173-183.
Direct Link - Benz, G.W., 1980. Progressive ontogenetic changes in the soluble nuclear eye lens proteins of the sandbar shark, Carcharhinus milberti (Valenciennes). Comp. Biochem. Physiol., 67: 191-193.
CrossRef - Binoy, V.V., 2010. Catfish Clarias is vanishing from the waters of Kerala. Curr. Sci., 99: 714-714.
Direct Link - Bon, W.F., G. Ruttenberg, A. Dohrn and H. Batink, 1968. Comparative physicochemical investigations on the lens proteins of fish. Exp. Eye Res., 7: 603-604.
CrossRef - Bours, J., 1980. Species specificity of the crystallins and the albuminoid of the ageing lens. Comp. Biochem. Physiol., 65: 215-222.
CrossRef - Calhoun, W.B. and Y.L. Koenig, 1970. Distribution of soluble proteins in the lenses of some marine vertebrates. Comp. Biochem. Physiol., 34: 71-80.
PubMed - Dahlman, J.M., K.L. Margot, J. Horwitz and M. Posner, 2005. Zebrafish alpha-crystallins: protein structure and chaperon-like activity compared to their mammalian orthologs. Mol. Vis., 11: 88-96.
PubMed - Day, T.H. and R.M. Clayton, 1973. Intraspecific variations in the lens proteins. Biochem. Genet., 8: 187-203.
PubMed - De Jong, W.W. and W. Hendriks, 1986. The eye lens crystallins: Ambiguity as evolutionary strategy. J. Mol. Evol., 24: 121-129.
PubMed - Eckroat, L.R. and J.E. Wright, 1969. Genetic analysis of soluble lens protein polymorphism in brook trout, Salvelinus fontinalis. Copeia, 1969: 466-473.
Direct Link - Islam, M.N., M.S. Islam and M.S. Alam, 2007. Genetic structure of different populations of walking catfish (Clarias batrachus L.) in Bangladesh. Biochem. Genet., 45: 647-662.
CrossRef - Jamieson, A. and A.S. Teixeira, 1991. Eye lens proteins as genetic markers in anguillids. Anim. Genet., 22: 45-52.
CrossRef - Kiss, A.J., A.Y. Mirarefi, S. Ramakrishnan, C.F. Zukoski, A.L. Devries and C.H.C. Cheng, 2004. Cold-stable eye lens crystallins of the antarctic nototheniid toothfish dissostichus mawsoni norman. J. Exp. Biol., 207: 4633-4649.
PubMed - Koehn, R.K. and D.I. Rasmussen, 1967. Polymorphic and monomorphic serum esterase heterogeneity in catostomid fish populations. Biochem. Genet., 1: 131-144.
PubMed - Laemmli, U.K., 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature, 227: 680-685.
CrossRefDirect Link - Phan, V.N., H. Suzuki, V. Gomes and M. Passos, 1985. Notes on electrophorograms of eye-lens muscle proteins and zymograms of esterase of fish collected during the first Brazilian expedition to antarctica. Bolm. Inst. Oceanogr., 32: 201-212.
Direct Link - Piatigorsky, J. and G.J. Wistow, 1989. Enzyme/Crystallins: Gene sharing as an evolutionary strategy. Cell, 57: 197-199.
PubMed - Khan, S., M.A. Khan, K. Miyan and M. Mubark, 2011. Length-weight relationship of nine freshwater teleosts collected from river Ganga, India. Int. J. Zool. Res., 7: 401-405.
CrossRefDirect Link - Samad, M.A., M.A. Islam and M.A. Khaleque, 2005. Effect of stocking density on the growth and survival rate of magur (Clarias batrachus) fry in laboratory and nursery ponds. Pak. J. Biol. Sci., 8: 338-344.
Direct Link - Adekunle, A.I., 2011. Growth and blood chemistry of selected clariid catfishes fed phosphorus supplemented diets. J. Fish. Aquat. Sci., 6: 545-554.
CrossRef - Smith, A.C. and H.B. Clemens, 1973. A population study by proteins from the nucleus of bluefin tuna eye lens. Trans. Am. Fish. Soc., 102: 578-583.
CrossRef - Smith, A.C. and R.L. Gilman, 1982. Electrophoretic study of proteins from solubilized eyelens nuclei of fishes. Comp. Biochem. Physiol. B, 71: 337-343.
PubMed - Suzuki, H. and V.N. Phan, 1990. Electropboretic study on intraspecific variations and interspecific relationships of marine catfishes (Siluriformes, Ariidae) of Cananeia (Sao Paulo, Brazil). 1. General proteins of eyelens and skeletic muscle. Boletim Inst. Oceanogr., 38: 31-42.
Direct Link - Yu, C.M., G.G. Chang, H.C. Chang and S.H. Chiou, 2004. Cloning and characterization of a thermostable catfish alphaB-crystallin with chaperon-like activity at high temperature. Exp. Eye Res., 79: 249-261.
PubMed - Zigler, Jr. J.S. and J.B. Jr. Sidbury, 1976. A comparative study of the β-crystallins of four sub-mammalian species. Comp. Biochem. Physiol., 55: 19-24.
PubMed - Zigman, S. and T. Yulo, 1979. Eye lens ageing in the dogfish (Mustelus canis). Comp. Biochem. Physiol. B, 63: 379-385.
PubMed - Bradford, M.M., 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem., 72: 248-254.
CrossRefPubMedDirect Link