
Kitherian Sahayaraj
Department of Advanced Zoology and Biotechnology, Crop Protection Research Centre, St. Xavier`s College (Autonomous), Palayamkottai-627 002, Tamil Nadu, India
Nithyanatham Auxelia
Department of Advanced Zoology and Biotechnology, Crop Protection Research Centre, St. Xavier`s College (Autonomous), Palayamkottai-627 002, Tamil Nadu, India
Journal of Biological Sciences
Year: 2012 | Volume: 12 | Issue: 3 | Page No.: 192-196







ABSTRACT
UV 254 nm radiation generally considered as a germicide and its impact has not been studied against highly mobile agricultural pest, Dysdercus cingulatus Fab. Newly hatched egg of D. cingulatus Fab. was exposed with UV 254 nm radiation and the consequence such as nymphal development and survival, morphogenetic effect and whole body protein and genomic DNA content were evaluated under laboratory conditions. Exposure of UV 254 nm radiation prolonged the nymphal developmental period from 1-7 days, reduced the nymphal survival rate and caused both in nymphs and adults morphological deformities. However, the total body protein contents increased up to third instar of D. cingulatus and then it decreased. Whereas, the whole body genomic content increased from 5-20 min UV-C exposure. Hence, UV 254 nm radiation has been considered for its impact on terrestrial animals including insect pests.
PDF Abstract XML References Citation
Received: February 26, 2012;
Accepted: March 12, 2012;
Published: June 12, 2012
How to cite this article
Kitherian Sahayaraj and Nithyanatham Auxelia, 2012. Stress Response of Different Exposure Time by UV 254 on the Biology and Body Total Protein and Genomic DNA Content of Red Cotton Bug. Journal of Biological Sciences, 12: 192-196.
DOI: 10.3923/jbs.2012.192.196
URL: https://scialert.net/abstract/?doi=jbs.2012.192.196
DOI: 10.3923/jbs.2012.192.196
URL: https://scialert.net/abstract/?doi=jbs.2012.192.196
INTRODUCTION
In the visible spectra (VIBGYOR) of the light, the region of radiation from 280-400 nm is called Ultraviolet Radiation (UV). Within the UV portion is possible to distinguish three types of wavelengths: UV-C (100-280 nm), UV-B (290-320 nm) and UV-A (320-400 nm) (Diaz and Fereres, 2007). Unlikely the radiation present in the terrestrial sunlight, the division between the UV-B and UV-C of 290 nm has shorter wavelength (Hollosy, 2002). It is a general observation recorded by many scientist that shorter wavelength light has more energetic and cause maximum damaging to microbes (Ozcelik, 2007; Sini et al., 2007; Rachh et al., 2010); plants (Yuan et al., 2006; Farokh et al., 2010) and animals. However, it has been used for germicidal use (Wells et al., 2011).
UV-A, UV-B and UV-C radiations causes damage to all the living organisms. UV-C radiation is the most potent radiation among these three types of UV radiation and acts at short range. Though, UV-C caused maximum damage, however, it does not pose any risk to earth life creatures (Smith et al., 2009). Orientation, navigation, host finding and feeding behaviors have been altered by various wavelength of UV regions (Antignus and Ben-Yakir, 2004). It is essential to evaluate the impact of various UV regions exposure on insects. Because they are always crawling and moving over the plants both in day and night time. Impact of UV radiation affects both freshwater (Hader and Sinha, 2005) and marine water dwelled organisms (Hernandez et al., 2007) have been documented elsewhere. Impact of UV-C on stored products pests (Faruki, 2005; Faruki et al., 2005, 2007) and on other insect pests (Zhou and Steller, 2003; Needham et al., 2006; Weintraub et al., 2008) including Dysdercus koenigii heteroptera were available in the literature.
Dysdercus cingulatus Fab. (Pyrrhocoridae) is the most serious and predominant insect pest of cotton in Southeast Asian countries and having many alternative hosts belongs to Malvaceae and Bombacaceae family (Kohno and Thi, 2004; Pandey et al., 2010). Both nymphs and adults are moving from one part of the plant to another for feeding and hence, there are a lot of possibilities of phasing sunlight directly. Usually neem has been used for the management of this pest (Pandey and Tiwari, 2011). Moreover, the possibility of direct impact of UV-C to insect biology, morphology and whole body total protein content has received little attention. This study was undertaken under the laboratory conditions, to study the impact of UV-C radiation on the biology, morphology and total protein content of a common cotton pest, red cotton bug D. cingulatus.
MATERIALS AND METHODS
Insects: Life stages of D. cingulatus were collected from cotton and bhendi fields of Tirunelveli District, Tamil Nadu, India and were maintained under laboratory condition 27±2°C temperature, 70-75% RH, 11 L and 13 D h Photoperiod in plastic containers 20x10x15 cm. on water soaked cotton seeds. The laboratory emerged first to fifth instar nymphs were used for the experiments.
UV-C light exposure: Twenty newly emerged first, second, third and fourth instar D. cingulatus >6 h. nymph was randomly selected from the stock culture and expose them directly to UV-C lamp of 254 nm (G30T8 TUV 30W, Philips) were 895 mm length, applied at a distance from the lamb of approximately one foot for different duration of 05, 10 and 20 min. Previously insects were exposed to 1, 2 and 4 min, since no impact was recorded, we selected 05, 10 and 20 min. For exposure six replications n = 120 were maintained for each nymphal instar. After the UV-C treatment, insects were transferred into a plastic container (20x10x15 cm) and maintained till their death using water soaked cotton seeds. Molting times, abnormalities if any, mortality of the insect was recorded from all the exposure categories.
Total body protein and genomic DNA analysis: Moribund/dead insects were collected, maintained at -4°C in a freezer and used for the total protein estimations. Randomly select 6 dead and alive insects each and homogenised with 1.0 mL of phosphate buffer, in a chilled pestle and mortar and centrifuged at 12,000 rpm for 6 min at 4°C. The supernatant was used for the estimation of total body protein (Bradford, 1976). Whole body total genomic DNA was extracted from 4-6 insects from each treatment using a modification of a general procedure for extraction with phenol (Sambrook et al., 1989). The insects were crushed and incubated at 40°C in 0.6 mg mL-1 Proteinase K and 300 μL TNES buffer 50 mM tris-HCl, pH 7.5, 0.4 M NaCl, 20 mM EDTA, 0.5% SDS for 4-8 h. DNA was then purified by washing with organic solvents: once with a chloroform: isoamyl mix 24:1 v/v.; once with a chloroform: phenol mix 1:1 v/v. and once with chloroform only. DNA was then precipitated with absolute ethanol and OD was taken in UV-Visible spectrophotometer at 580 nm.
Data analysis: Statistical comparison was made between control category to 0.5, 10.0 and 20.0 min UV-C radiation exposed individually by Tukey Multiple Range Test (TMRT). and the significance were expressed at 5% level using SPSS ver 11.5.
RESULTS AND DISCUSSION
Egg hatching: Firstly, analyze the hatchability of the irradiated D. cingulatus eggs. The egg hatchability of the irradiated egg was drastically reduced 45.0, 5.0 and 0% for 5, 10 and 20 min exposure, respectively when compared to non-irradiated eggs (91.66%). Similarly, an increase in UV-C exposure time gradual decreased the percentage of Tribolium castaneum and T. confusum egg hatching (Faruki et al., 2007). These results suggest that the irradiation to the eggs may induce some stresses in hatchability of the eggs.
Nymphal development and survival: Nymphal developmental period of D. cingulatus was gradually increased from first instar to fifth nymphal instars (Table 1). Irrespective of the UV-C exposure time, D. cingulatus total nymphal developmental period was significantly increased (3, 4 and 6 days for 5, 10 and 20 min exposure, respectively; p<0.05) than the control category. However, no difference was observed in the total nymphal developmental time of Myzus persicae Sulzer. (Homoptera: Aphididae), when reared under various UV-absorbing films methods (Chyzik et al., 2003). They further reported that the elimination of UV from the light spectrum has no significant effect on the generation development time of Myzus persicae. However, it is interesting that the death rates of the aphids were 1.5-2 times higher under the regular films than under the UV-absorbing ones, both in cages and in walk-in tunnels. To the best of our knowledge this is the first report available about the inhibitory effect of UV-C radiation on D. cingulatus.
Nymphal survival following irradiation was 78.3, 72.6 and 70.6% for 5, 10 and 20 min UV-C exposure, respectively and non-irradiated nymphal survival was 94.9%. Overall manner, there was does and time of exposure dependent radiation effect on hatchability and nymphal survival (Fig. 1).
| Table 1: | Impact of UV-C radiation exposure time (min) on the nymphal developmental period (days) of red cotton bug |
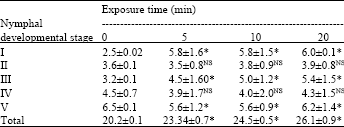 | |
| Different exposure period data of individual nymphal instar compared with the control data, Significance at 5% level by TMRT, NS: Significant | |
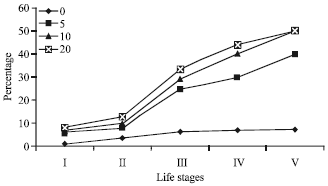 | |
| Fig. 1: | Impact of UV-C radiation exposure periods (min) on the nymphal mortality percent of red cotton bug |
It was previously reported that adult emergence was significantly decreased when larvae of T. castaneum (Faruki, 2005) and Alphitobius diaperinus were exposed to UV radiation as observed in the present study 94.8, 78.0, 72.6 and 70.2% for control, 5, 10 and 20 min UV exposed insects, respectively. A study by Needham et al. (2006) using dust-mite Dermatophagoides farinae, has shown that an exposure of 5 sec is sufficient to kill 100% of newly laid eggs. This is likely to be due to the induction of self-destruction as demonstrated using UV-C and eggs of the fly Drosophila (Zhou and Steller, 2003).
Several studies have shown that whiteflies (Matteson et al., 1992), thrips (Ben-Yakir et al., 2008), Myzus persicae and its hymenopteran parasite (Chyzik et al., 2003) and leafhoppers (Weintraub et al., 2008) either chooses environments where UV radiation is present or disperse less where there is no UV radiation. It has been shown that thrips have maximum visual acuity in the UV range and move toward UV-A radiation source (Sinha and Hader, 2002). Thus, these physiological data and recent reports (Ben-Yakir et al., 2008), strengthen our results, where thrips invasion is limited in UV-absorbing covers. The general effect of UV-absorbing covers that we observed, the reduction in the immigration rate of insects, is in agreement with other studies (Chyzik et al., 2003).
Morphogenesis: When UV-C radiation exposure time was increased, 18.02% adults were emerged with abnormalities in wings, legs and antennae. 7.66% of nymphs are unable to molt their exoskeleton and it leads to the death. In addition to the wing abnormalities, the wing color was also changed from red to pale red and this might be due to the interruption of UV-radiation in pigment production. Further studies are needed to confirm this hypothesis. Moreover, Raviv and Antignus (2004) and Krizek et al. (2005) have also reported that UV radiation interrupts the life cycle of several pathogenic fungi and alters the visual behavior of many insects.
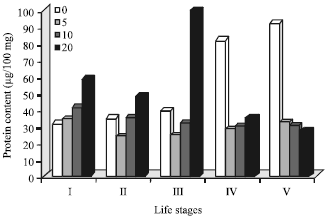 | |
| Fig. 2: | UV-C radiation exposure time (min) on the total body protein content (μg/100 mg) of red cotton bug nymphal status |
Whole body protein and genomic DNA content: Impact of UV-C radiation on the whole body protein content of D. cingulatus life stages is presented in Fig. 2. It was very clear from the results that, protein content was gradually increased from the first instar to fifth instar. Though maximum DNA content was observed in the fifth nymphal instar of D. cingulatus, it was highly reduced (70%) in 5 min UV-C radiation treatment. It was further reduced 71.4 and 75.53 times when D. cingulatus fifth instar exposed to UV-C for 10 and 20 min, respectively. Similar trend was also observed for fourth instar too. Both in first and second instars also the total DNA content was gradually increased from control to 20 minutes UV radiation exposed insect. UV-damaged DNA and induce the alternation of binding proteins (Sugasawa et al., 2005).
UV is able to create two of the most mutagenic and cytotoxic DNA lesions: cyclobutane-pyrimidine dimers and 6-4 photoproducts (Sinha and Hader, 2002). Organisms have therefore developed a number of repair mechanisms to counteract the DNA damage caused by UV. These repair mechanisms will however be unable to cope if the UV dose applied is higher than the repair capacity. As the result total genomic content has been gradually decreased from 5 min UV-C exposure times to 20 min in all the nymphal instars of the pest except the first instars. Furthermore, the whole body DNA content was gradually increased from the first instar to the fifth instar in the non-irradiated D. cingulatus. However, the DNA content was gradually increased from 5-20 min irradiated first to fourth instar nymphs. However, DNA content in 5 minutes treatment was lesser (p<0.05) than control categories (Table 2). Though, maximum DNA content was observed in the fifth nymphal instar of D. cingulatus, it was highly reduced (70%) in 5 min UV-C radiation treatment. It was further reduced 71.4 and 75.53 times when D. cingulatus fifth instar exposed to UV-C for 10 and 20 min, respectively.
| Table 2: | Influence of UV-C exposure time (min) on the whole body genomic DNA content (μg mL-1) of D. cingulatus nymphal instars |
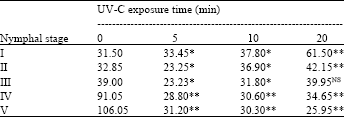 | |
| Different exposure period data of individual nymphal instar compared with the unexposed insect data, Vlalues are significant at *5 and **10% level by TMRT, NS: Not significant | |
The DNA is one of the key targets for UV-induced damage as reported by Sinha and Hader (2002).
CONCLUSION
In conclusion, Dysdercus cingulatus is a serious pest of cotton and distributed all the cotton growing region of many Asian countries. It is highly mobile pest, moving from place to place and exposed to sun light frequently where UV-C is reported. Results revealed that UV 254 irradiated eggs altered the developmental parameters, drastically reduce the total genomic DNA content except in the first instar and total protein content which leads to morphological changes wing deformity which lead to the death of the organisms. This study highlighting that germicidal radiation UV 254, not only having surface sterilization capacity, it would have penetration capacity to affects higher organisms which would provide a technical or researchable advance in a irradiation impact on higher organisms study.
ACKNOWLEDGMENTS
Authors thank Ministry of Earth and Sciences Ref. No. MRDF/01/33/P/07. for the financial assistance and authorities of St. Xavier’s College Autonomous., Palayamkottai for the laboratory facilities and encouragements.
REFERENCES
- Ben-Yakir, D., M.D. Hadar, Y. Offir, M. Chen and M. Tregerman, 2008. Protecting crops from pests using Optinet® screens and Chromatinet® shading nets. Acta Hortic., 770: 205-212.
Direct Link - Ozcelik, B., 2007. Fungi/bactericidal and static effects of ultraviolet light in 254 and 354 nm wavelengths. Res. J. Microbiol., 2: 42-49.
CrossRefDirect Link - Bradford, M.M., 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem., 72: 248-254.
CrossRefPubMedDirect Link - Chyzik, R., S. Dobrinin and Y. Antignus, 2003. Effect of a UV-deficient environment on the biology and flight activity of Myzus persicae and its hymenopterous parasite Aphidius matricariae. Phytoparasitica, 31: 467-477.
CrossRefDirect Link - Diaz, B. and A. Fereres, 2007. Ultraviolet-blocking materials as a physical barrier to control insect pests and plant pathogens in protected crops. Pest Technol., 1: 85-95.
Direct Link - Farokh, P., H. Mahmoodzadeh and T.N. Satari, 2010. Response of seed germination of safflower to UV-B radiation. Res. J. Environ. Sci., 4: 70-74.
CrossRefDirect Link - Faruki, S.I., D.R. Das and S. Khatun, 2005. Effects of UV-radiation on the larvae of the lesser mealworm, Alphitobius diaperinus (Panzer) (Coleoptera: Tenebrionidae) and their progeny. J. Biol. Sci., 5: 444-448.
CrossRefDirect Link - Faruki, S.I., D.R. Das, A.R. Khan and M. Khatun, 2007. Effects of ultraviolet (254nm) irradiation on egg hatching and adult emergence of the flour beetles, Tribolium castaneum, T. confusum and the almond moth, Cadra cautella. J. Insect Sci., 7: 1-6.
CrossRefPubMedDirect Link - Hader, D.P. and R.P. Sinha, 2005. Solar ultraviolet radiation-induced DNA damage in aquatic organisms: Potential environmental impact. Mutation Res./Fundam. Mol. Mech. Mutagen., 571: 221-233.
CrossRefDirect Link - Hernandez, K.L., R.A. Quinones, G. Daneri, M.E. Farias and E.W. Helbling, 2007. Solar UV radiation modulates daily production and DNA damage of marine bacterioplankton from a productive upwelling zone (36°S), chile. J. Exp. Mar. Biol. Ecol., 343: 82-95.
CrossRefDirect Link - Hollosy, F., 2002. Effects of ultraviolet radiation on plant cells. Micron, 33: 179-197.
CrossRefPubMedDirect Link - Kohno, K. and N.B. Thi, 2004. Effects of host plant species on the development of Dysdercus cingulatus (Heteroptera: Pyrrhocoridae). Applied Entomol. Zool., 39: 183-187.
CrossRefDirect Link - Krizek, D.T., H.D. Clark and R.M. Mirecki, 2005. Spectral properties of selected UV-blocking and UV-transmitting covering materials with application for production of high-value crops in high tunnels. Photochem. Photobiol., 81: 1047-1051.
CrossRefPubMedDirect Link - Yuan, L., Z. Yanqun, H. Yongmei, C. Jianjun, C. Haiyan, H. Lilian and L. Fusheng, 2006. Intraspecific responses in flavonoid contents of 28 wild sugarcane clones to enhanced ultraviolet-b radiation under field conditions. Am. J. Plant Physiol., 1: 151-159.
CrossRefDirect Link - Matteson, N., I. Terry, A. Ascoli-Christensen and C. Gilbert, 1992. Spectral efficiency of the western flower thrips, Frankliniella occidentalis. J. Insect Physiol., 38: 453-459.
CrossRefDirect Link - Abu-Sini, M., K.H. Abu-Elteen and A.Z. Elkarmi, M.A. Hamad and R.F. Khuzaie, 2007. Influence of various ultraviolet light intensities on pathogenic determinants of Candida albicans. Biotechnology, 6: 210-217.
CrossRefDirect Link - Needham, G., C. Begg and S. Buchanan, 2006. Ultraviolet C exposure is fatal to American house dust mite eggs. J. Allergy Clin. Immunol., 117: S28-S28.
CrossRefDirect Link - Pandey, J.P. and R.K. Tiwari, 2011. Neem based insecticides interaction with development and fecundity of red cotton bug, Dysdercus cingulatus Fab. Int. J. Agric. Res., 6: 335-346.
CrossRefDirect Link - Rachh, P.R., M.R. Rachh, M.M. Soniwala, V.D. Joshi and A.P. Suthar, 2010. Effect of gymnema leaf extract on uv-induced damage on Salmonella typhi. Asian J. Biol. Sciences, 3: 28-33.
CrossRefDirect Link - Raviv, M. and Y. Antignus, 2004. UV radiation effects on pathogens and insect pests of greenhouse-grown crops. Photochem. Photobiol., 79: 219-226.
CrossRefDirect Link - Sinha, R.P. and D.P. Hader, 2002. UV-induced DNA damage and repair: A review. Photochem. Photobiol. Sci., 1: 225-236.
CrossRefPubMedDirect Link - Smith, W.K., W. Gao and H. Steltzer, 2009. Current and future impacts of ultraviolet radiation on the terrestrial carbon balance. Front. Earth Sci. China, 31: 34-41.
CrossRefDirect Link - Sugasawa, K., Y. Okuda, M. Saijo, R. Nishi and N. Matsuda et al., 2005. UV-induced ubiquitylation of XPC protein mediated by UV-DDB-ubiquitin ligase complex. Cell, 121: 387-400.
CrossRefDirect Link - Weintraub, P.G., S. Pivonia and A. Gera, 2008. Physical control of leafhoppers. J. Econ. Entomol., 101: 1337-1340.
CrossRefDirect Link - Wells, J.B., C.D. Coufal, H.M. Parker, A.S. Kiess, J.L. Purswell, K.M. Young and C.D. McDaniel, 2011. Hatchability of broiler breeder eggs following eggshell sanitization by repeated treatment with a combination of ultraviolet light and hydrogen peroxide. Int. Poult. Sci., 10: 421-425.
CrossRefDirect Link - Zhou, L. and H. Steller, 2003. Distinct pathways mediate UV-induced apoptosis in Drosophila embryos. Dev. Cell, 4: 599-605.
CrossRefDirect Link