
Jayant Biswas
National Cave Research and Protection Organization, Central Zone, Raipur 492001, Chhattisgarh, India
Daniel B. Harries
School of Life Sciences, Heriot-Watt University, Edinburgh, Scotland EH14 4AS, UK
Journal of Biological Sciences
Year: 2011 | Volume: 11 | Issue: 7 | Page No.: 459-465







ABSTRACT
The subterranean mode of life always needs a high degree of biological adaptation. The cavernicolous species usually get selected from those which are preadapted with biological traits suitable for cave life and finally occupied a separate taxonomic status. With the ever going exploration of subterranean passages in Jaintia hills, till date several troglobitic species have been recorded from this particular area. Cave biota survey with respect to the explored subterranean passages of the area was conducted in February 2011. Krem Bylliat (cave) is one of them, measure horizontally 600 m having several openings. The cave is situated in the boundary of two major rock formations, nearer to one of the main tributary of river Kopili. Though, the cave is harbour of rich biodiversity, it was found to be relatively impoverished in terms of troglomorphic taxa. Possible factors, obviating the evolution of troglomorphy in this cave have been discussed.
PDF Abstract XML References Citation
Received: August 09, 2011;
Accepted: October 22, 2011;
Published: December 10, 2011
How to cite this article
Jayant Biswas and Daniel B. Harries, 2011. Krem Bylliat: The Harbour of Precedent Cavernicolous Representatives from the Jaintia Hills, Meghalaya, India. Journal of Biological Sciences, 11: 459-465.
DOI: 10.3923/jbs.2011.459.465
URL: https://scialert.net/abstract/?doi=jbs.2011.459.465
DOI: 10.3923/jbs.2011.459.465
URL: https://scialert.net/abstract/?doi=jbs.2011.459.465
INTRODUCTION
Perpetual darkness, sporadic supply of nutrients, near constant environmental conditions and high CO2 percentage are some of the common features for almost all the subterranean caves. Although all these together make caves inhospitable, many organisms inhabit subterranean caves successfully. High degree of physiological adaptations, behavioural adjustments and phenotypic alterations are needed to establish a population permanently in such conditions (Vandel, 1965; Gunn, 2004; Biswas, 2009, 2010) and due to which the most common characters appears in them are absence of pigment, reduction of the optic apparatus (eyes), elongation of appendages and developments of extra sensory organs often referred as troglomorphic traits. On the other hand, low predation pressure, high humidity, easy availability of prey and even the negligible effects of natural environmental calamities (Biswas and Shrotriya, 2011) are some of the other factors of the same cave that altogether attract many opportunistic species to exploit these habitats. In general cave dwelling/adapted organisms usually evolve from ancestral species with pre-adapted biological traits, favouring the cave life (Vandel, 1965; Gunn, 2004). Conclusively, two types of organisms could be found inside any cave; (1) the animals once trapped accidentally or get intentionally into the cave start successfully their phylogenetic race inside the cave, referred as obligatory type, (2) the animal frequently visiting or almost settled inside the cave for fulfillment of their certain needs of life, such as feeding, roosting, hibernating, breeding etc. can be referred as opportunistic. Opportunistic types are often seen to depend on their ambient external environment for some of their biological needs. However, the opportunistic cave dwellers are often acting as empirical models of natural selection and adaptation in the subterranean environment (Kane and Culver, 1992; Sahu et al., 2011).
State Meghalaya contains some of the most significant areas of karst within the Indian subcontinent. The limestone is of Cretaceous to Miocene age and it forms an almost continuous outcrop along the border with Bangladesh on the southern margin of Meghalaya (Fig. 1b). Much of this area has high elevation and the limestone deposits can have a depth potential up to 500 m. The state is also well known for its high rainfall with southern Meghalaya receiving more than 10,000 mm rainfall per year (Rai, 2010). The well developed limestone deposits and high rainfall has resulted in the development of numerous and extensive subterranean drainage passages.
The Jaintia Hills district of state Meghalaya is of particular importance as a cavernous area due to the large extent of karst and general high elevation (up to 1200-1500 m). The area is mostly composed of Tertiary rocks of the Jaintia Group and the group itself represents the entire eocene shelf sediments of Meghalaya (Samanta, 1971).
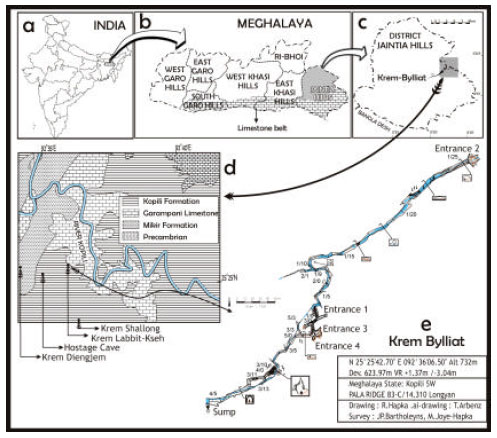 | |
| Fig. 1(a-e): | (a) Subcontinent India; indicating the location of state Meghalaya (b) State Meghalaya; indicating the location of district Jaintia Hills alongwith the existing outcrop Limestone belt (c) District Jaintia Hill; indicating the karst-complex location and where the cave “Krem Bylliat” exist (d) Location where Krem Bylliat and few other caves exists and (e) Map of Krem Bylliat, locating each and every entrance, water canals and dripstone galleries |
Many hundreds of caves have been recorded within the Jaintia Hills and some are internationally significant in terms of extent (e.g., passage lengths up to 30 km and depth range up to 300 m). Each year further caves are added to the records and many kilometers of additional cave passages are mapped. The full extent of cavernous habitat in the Jaintia will not be established for many years. However, the biospeleology of the region is poorly known (Kottelat et al., 2007; Biswas, 2009; Harries et al., 2008; Disney, 2009) and only few caves have been subject to detailed biological study.
In the present study, a biotic survey report of Krem Bylliat; one of the recently explored cave of Jaintia hills has been documented. Its biotic components reflect almost the same picture which has already been documented earlier from the various subterranean passages of that area. However, the lacks of degree of troglomorphism in such taxa are of great interest. The possible factors suppressing the process of such evolution has been tried to correlate with the adjacent abiotic factors.
Surveyed cave description: Krem Bylliat (N 25°25’42.70”; E 92°36’06.50” (WGS84); altitude 732 m above sea level) is situated in Pala-range of Jaintia Hills within 50 m of the banks of the River Kopili. The total length of the cave has been mapped as 624 m with a vertical range of about 4.5 m (Fig. 1e) by Meghalayan Adventurers’ Association. The northern entrance (Entrance 2) of the cave is located at the base of a 3 m deep doline which is densely vegetated by bamboo trees (Fig. 2a). The cave begins as a phreatic tube; the passage floor from the entrance (Entrance 2) to the end of twilight zone is laterally divided into two parts (Fig. 2b). On the left the floor is dry and mostly covered by flowstone deposits whereas on the right there is a stream bedded by mud-clay, alluvial gravel and sandstone pebbles. The structure of the cave passage deeper within the cave remains broadly similar to this entrance area.
 | |
| Fig. 2(a-c): | Images of Krem Bylliat. (a) Entrance-2; surrounded by bamboo trees (b) Twilight zone (Entrance-2) clearly indicating the path divided into two parts; left flawstone formation; right water passage. Roof and side walls are clearly indicating two different formations and (c) One after one rimstone pools nearer to the Entrance 2 |
A small stream from the river tributary flows continuously through the cave and it appears that significant flooding occurs on a seasonal basis. About ~40 m within the cave from Entrance 2 there is a deep (approx. 2 m) canal (Fig. 1e). The twilight zone ends at this canal because three consecutive rimstone dams cross the canal and come sufficiently close to the ceiling to cut out the remaining light (Fig. 2c). Beyond the canal and dams the passage becomes considerably larger. At about 300 m within the cave there are three additional openings to the surface with associated twilight zones (Fig. 1e). Beyond these entrances the passage continues through areas with prominent calcite dripstone formations suspended from the ceiling before terminating at a sump at the southern end of the accessible cave.
Humidity and temperature were measured by digital thermo-hygrometer (Pacer® TH 402) and were found to be constant throughout the cave; i.e., 100% humidity and the atmospheric temperature 16°C.
Faunal components of the cave: In February 2011 the first author participated in an international cave mapping expedition (organized by Meghalayan Adventurers’ Association). The base camp was located in the Pala range of the Jaintia Hills in close proximity to Krem Bylliat. Over a fifteen day period the first author spent each day; three to four hours within the cave directly recording the fauna. All sub-habitats and crevices were carefully scrutinized and the fauna was photographed in situ. Aspects of their ecology, distribution, behaviour and response to disturbance were noted where appropriate.
NOTES ON BIOTA
Vertebrates: Myotis longipes Dobson, 1873, the microchiropteran bat is commonly known as the Kashmir cave-bat, because it was initially thought to be restricted to the caves of Kashmir (India). Subsequently, the species was reported from caves in adjacent countries such as Nepal and China (Talmale and Pradhan, 2009; Bates and Harrison, 1997). In Krem Bylliat; M. longipes was found roosting on cave walls in the twilight zone of Entrance-2. This area of the cave has a relatively low ceiling and frequent observations of partially consumed bat corpses indicate that they are vulnerable to predation (Fig. 3a). Similar reports of apparent bat predation have earlier been noted from Dandak cave of Chhattisgarh, India (Biswas and Shrotriya, 2011). M. longipes appears to have a wide distribution in Meghalaya and has previously been reported from Mawsmai Cave in the East Khasi Hills and Siju Cave in the South Garo Hills (Sinha, 1999a, b). Further, a small colony of rhinolophid bats (unidentified) was noted roosting in the roof of the passage well within the dark zone of the cave.
A small population of about 8-10 individuals of frog; Odorrana chloronota Günther, 1875 (Fig. 3b), was always apparent at the edges of the rimstone pools close to Entrance-2. The species is endemic to the Northeastern part of India and adjacent Southeast Asian countries (Bain et al., 2003; Stuart et al. 2006). Characteristic features include a spectacular vivid green dorsal colour and a peculiar odoriferous skin secretion. This frog was not found to respond any disturbance so quickly. Epigean populations of this frog are known to be nocturnal and occur in wet habitats. Presumably the combination of low light and damp conditions at this transition zone of the cave are favourable to the species.
The outer canal of the twilight zone near Entrance-2 contained a number of unidentified fish species including loach, carp and a species of Glyptothorax. The carp and Glyptothorax was found to occupy the bottom level of the canal and were very sensitive to any disturbance. The smaller fishes, especially the loach were usually apparent at the edges of the canal. Surprisingly, not a single specimen of any fish was found in the isolated rimstone water-pools exist a little further into the cave (Fig. 2c). The larger fish (especially Carp) were 15 cm or more in length, however not a single specimen represented any trace of troglomorphy.
Invertebrates: A shrimp population; Macrobrachium cavernicola Kemp, 1924 (Fig. 3e) was highly abundant in the outer canal of the twilight zone near Entrance-2 and smaller specimens were easily captured near the edges of the canal. The distinctive two-segmented mandible palps helped us to identify it as M. cavernicola. This troglomorphic species was first recorded from Siju Cave of Meghalaya, India (Kemp, 1924). The species has subsequently been recorded from caves throughout Meghalaya including many of the Jaintia caves (Biswas, 2009; Harries et al., 2008). Interestingly, the adult form of this shrimp was also recorded in the inner rimstone pools and in further deeper zone of the cave where no other large aquatic fauna were seen. The adults are completely albinic and have small eyes. The juveniles were always apparent nearer the entrance zones of all the caves surveyed by our team of that area (Fig. 1d).
The walls and roof of the Krem Bylliat was found to be occupied by Huntsman spider; Heteropoda fischeri Jager (2005) (Fig. 3f) which are very large spiders with the leg span of adults reaching up to 15 cm. It was 1st of all reported from the Krem Labbit which exist hardly 10 km apart from this cave.
 | |
| Fig. 3(a-h): | Organisms inhabits in Krem Bylliat. (a) Partially consumed corpses of a bat; Myotis longipes recovered from the twilight zone of Enterance-2, (b) Frog; Odorrana chloronota sitting in the edge of a rimstone pool, (c) Fishes apparent in the 1st water canal exist in the twilight zone of Entrance-2, (d) Crabs (possibly Maydelliathelphusa falcidigitis) present in the 1st water canal exist in the twilight zone of Entrance-2, (e) Partial troglomorphic shrimp; Macrobrachium cavernicola present in the water canal of inner chambers, (f) Huntsman spider; Hetropoda fischeri dominating throughout the cave, (g) Opiliones; apparent in all the enterance zone till twilight range and (h) Cricket; nymph of Rhaphidophoridae sp. |
The predator-prey relation between this spider and the cave crickets was already reported from other Jaintia caves (Harries et al., 2008). Further, the juveniles of an unidentified brown cricket were abundantly observed in the twilight zones of Krem Bylliat and even a group of these juveniles were seen feeding upon the dead body of a bat nearer the Entrance-2. The crickets were more sparse deeper within the cave where adults were also present.
Dark crabs; Maydelliathelphusa falcidigitis Alcock, 1910 (Fig. 3d) were noted in the twilight zone near Entrance-2 and often in some cavities exist in the deeper zone within the cave. It is also a common species reported from almost all the subterranean caves of Meghalaya (Biswas, 2009; Harries et al., 2008). Further, densely aggregated dark black coloured opiliones were frequently encountered on passage walls in every twilight as far back as the transient zones of this cave. They were highly sensitive to any type of disturbance and rapidly dispersed in response to small stimuli. Opiliones are usually omnivorous feeding upon materials such as small invertebrates, detritus, fungi, carrion, guano and other fecal material. They are adapted for life in dark conditions and use their second pair of legs as sensory antennae. These sensory adaptations and the ability to exploit a variety of food sources are likely to be advantageous in a cave habitat. It is possible that they exploit shaded environments simply to avoid desiccation and to link the cave environments is purely incidental. They have been noted in shaded rock gullies several meters beyond cave entrances and this indicates that the habitat preference might be for shade rather than specifically for cave entrances. Earlier also in addition to the troglomorphic Opiliones sp., similar aggregations of black opiliones have been recorded from the threshold area of several other caves in Meghalaya, (Harries et al., 2008). Surprisingly, not a single specimen of both the above species represents troglomorphism.
In addition to the above organisms, time to time few additionals were also seen inside the deep passages, but due to lack of their any permanent population we considered them either habitual or accidental trogloxene for this particular cave.
DISCUSSION
Till date several long and deep subterranean caves have been explored and mapped from Jaintia Hills of the state Meghalaya, India, most of them have passage lengths of many kilometers and a depth range of well over 100 m. This ensures an extensive ‘dark zone’ and a diversity of habitats including stable dry fossil levels as well as flood prone stream passages. In contrast; Krem Bylliat is a relatively small cave has multiple entrances and consequently no part of the cave is much more than 150 m from an entrance. It also has a very limited vertical depth range (~4.5 m) and consists almost entirely of flood prone streamway. These factors probably explain the relatively impoverished community recorded within the cave. In the present survey, none of the taxa in Bylliat exhibited pronounced troglomorphy. Among the tabulated ones, Krem Bylliat found to be shared six taxa with the other Meghalayan caves, few of which are very common. One of the Bylliat species showed partial troglomorphy (Macrobrachium cavernicola). Earlier reports already pleaded the existence of nine partially troglomorphic taxa in other Meghalayan caves and three of these (a species of beetle and two isopods) occur in almost every caves (Harries et al., 2008). In addition, few unidentified species were also noted in the adjacent caves (Fig. 1d) during the survey period (data unpublished). Notably, a troglobitic loach; Schistura papulifera was recorded from the adjacent cave “Hostage” in the region (Fig. 1d) which is slightly longer (746 m) than Krem Bylliat but has a greater vertical range (28 m).
Interestingly during the survey period, O. chloronota which is always present in this cave was not at all conspicuous in its adjacent caves (Fig. 1d). Possibly, the twisted and/or vertical entries of such caves act a hurdle for its establishment. Nevertheless, the lacks of its population from such caves having large entry and big water canals are of great interest. However, there is currently no evidence to suggest, they sustain stable reproducing populations within the caves.
Krem Bylliat contains many of the species that have been previously noted as characteristic components of the Meghalayan cave communities (e.g., bats, cyprinids, loach, Glyptothorax, Macrobrachium, freshwater crabs, Heteropoda, brown rhaphidophorids) and it appears typical of superficial cave habitats in the Jaintia Hills. Though, the distribution of fauna within Krem Bylliat is broadly similar to that of other caves in the Jaintia Hills (Harries et al., 2008), the apparent partial/complete troglomosphism in such species are completely lacking. In Krem Bylliat the fish species were restricted to the threshold area but previous records demonstrate that these taxa also frequently occur deep within caves. The reason for their absence from the dark zone of Krem Bylliat is interesting. It could be speculated that, due to having the multiple entrances in Krem Bylliat the deep zone and/or stagnant zones are completely lacking and due to which the required habitat for complete troglomorphy is absent here. Further, given the degree of water flow during the rainy season it is unlikely that the subterranean and epigean populations are spatially segregated. The species may be sufficiently adaptable to exploit both cave and surface habitats depending on conditions. Further study of the ecology of this species would prove interesting.
Macrobrachium cavernicola did occur in the dark zone of Krem Bylliat and were also found in surface streams in the vicinity of this and other (Krem-Labbit Ksheh, Krem-Diengjem, Krem-Shallong) caves. Earlier besides the deeper zones, a subterrnaen Solvenian shrimps Troglocaris sp. has already been reported to observe in the illuminated parts of the caves (Sket, 1965). Partial troglomorphism in it suggests that the species is highly pre-adapted to dark and other geophysical conditions of subterranean mode of life and thus could be able to adopt quickly the subterranean mode of life.
It is likely that the main energy supply to the biota of Krem Bylliat is derived from material carried on the cave stream. Like many cavernicolous species most of the taxa are probably generalist feeders able to exploit a variety of food sources opportunistically. Of course the Heteropoda are specialist predators but a plentiful food source is available in the form of the brown cave crickets and the close proximity of the surface means that occasional stray surface species are also likely to be available as prey items. The bat populations are another potential energy source and carcasses were certainly seen to provide a food source for the brown cave crickets. No significant guano deposits were found and this is true of many of the Megalayan caves presumably because guano is regularly swept away by seasonal floods. The bats themselves were seen to be subject to predation (Fig. 1a) but the identity of the predator is unknown. It is likely that bats roosting in Krem Bylliat are particularly accessible to external predators due to the limited extent and multiple entrances of the cave.
The study reinforces the conclusion that there is a degree of predictability to the composition of the cavernicole communities in the region and illustrates the relationships between habitat extent and cavernicole diversity and between surface proximity and frequency of occurrence of troglomorphic species. Finally, we have also to accept the hypothesis that the degree of troglomorphy does not absolutely reflect the strength of the “troglobiosis”(Sket, 2008).
ACKNOWLEDGMENT
We are very grateful to Mr. Brain D Kharpran (Meghalayan Adventurers Association), Mr. Thomas Arbenz and Swiss Society of Speleology for allowing us to use the map of Krem Bylliat to use in this manuscript. Thanks are also due towards Dr. Manuel Ruedi for his time to time unreserved help during the field study and the Natural History Museum of Geneva, Switzerland, for financial support to the 1st author conduct this survey.
REFERENCES
- Biswas, J., 2009. The biodiversity of Krem Mawkhyrdop of Meghalaya, India, on the verge of extinction. Curr. Sci., 96: 904-910.
Direct Link - Biswas, J., 2010. Kotumsar Cave biodiversity: A review of cavernicoles and their troglobiotic traits. Biodivers. Conser., 19: 275-289.
CrossRefDirect Link - Biswas, J. and S. Shrotriya, 2011. Dandak: A mammalian dominated cave ecosystem of India. Subterranean Biol., 8: 1-8.
CrossRefDirect Link - Bain, R.H., A. Lathrop R.W. Murphy, N.L. Orlov and C.T. Ho, 2003. Cryptic species of a cascade frog from Southeast Asia: Taxonomic revisions and descriptions of six new species. Am. Museum Novitates, 3417: 1-60.
CrossRefDirect Link - Disney, R.H.L., 2009. Scuttle flies (Diptera: Phoridae) from caves in Meghalaya, India. J. Cave Karst Stud., 71: 81-85.
Direct Link - Harries, D.B., F.J. Ware, C.W. Fisher, J. Biswas and B.D. Kharpran-Daly, 2008. A review of the biospeleology of Meghalaya, India. J. Cave Karst Stud., 70: 163-176.
Direct Link - Jager, P., 2005. New large-sized cave-dwelling Heteropoda species from Asia, with notes on their relationships (Araneae: Sparassidae: Heteropodinae). Revue Suisse de Zoologie, 112: 87-114.
Direct Link - Kottelat, M., D.R. Harries and G.S. Proudlove, 2007. Schistura papulifera, a new species of cave loach from Meghalaya, India (Teleostei: Balitoridae). Zootaxa, 1393: 35-44.
CrossRefDirect Link - Sahu, K.R., J. Biswas, K.V. Achari and K.M. Sinha, 2011. Ecological approval for cave habitat: The occurrence of regressed stridulatory system in cavernicolous Homoeogryllus sp. Int. J. Zool. Res., 7: 369-375.
CrossRefDirect Link - Samanta, B.K., 1971. Early tertiary stratigraphy of the area around Garampani, Mikir-North Cachar Hills, Assam. J. Geol. Soc. India, 12: 318-327.
Direct Link - Sket, B., 2008. Can we agree on an ecological classification of subterranean animals?. J. Nat. Hist., 42: 1549-1563.
CrossRefDirect Link - Stuart, B.L., R.F. Inger and H.K. Voris, 2006. High level of cryptic species diversity revealed by sympatric lineages of Southeast Asian forest frogs. Biol. Lett., 2: 470-474.
CrossRefDirect Link