
P. Praveen Reddy
Department of Research and Studies in Biotechnology, Gulbarga University, Gulbarga-585106, India
G.S.N. Reddy
Center for Cellular and Molecular Biology, Uppal-Hyderabad-580032, India
M.B. Sulochana
Department of Research and Studies in Biotechnology, Gulbarga University, Gulbarga-585106, India
Journal of Biological Sciences
Year: 2010 | Volume: 10 | Issue: 3 | Page No.: 237-241







ABSTRACT
β-fructofuranosidases are a group of hydrolytic and transfructosylating enzymes that play an important role in food processing industry to synthesize Fructo Oligosaccharides (FOS). The present study aimed to screening of filamentous fungi strains and growing them in batch cultures to compare their abilities in production of β-Fructofuranosidase (β-FFase) using sole carbon source as sucrose. Radial diffusion method was used to evaluate the potent β-fructofuranosidase producers by using crude enzyme. Among screened strains, Aspergillus niger PSSF21 (2.20±0.100 cm), Chrysonilia sitophila PSSF84 (2.00±0.026 cm) and Aspergillus flavus PSSF22 (1.64±0.040 cm) showed significant hydrolysis zone in the plate. In addition, the β fructofuranosidase of the above strains also showed high fructosyltransferase activity to hydrolytic ratio (Aspergillus niger PSSF21-6.335 U mg-1, Chrysonilia sitophila PSSF84 3.487 U mg-1, Aspergillus flavus PSSF22-3.256 U mg-1). The effects of pH and temperature on the sucrose- β-FFase reaction rate were studied using a 32 experimental design matrix. Acidic pH influences positively on the fructosyl transferase activity of three isolated strains, where as effect of temperature showed remarkable variations on high fructosyltransferase activity-to-hydrolytic activity ratio.
PDF Abstract XML References Citation
How to cite this article
P. Praveen Reddy, G.S.N. Reddy and M.B. Sulochana, 2010. Screening of β-Fructofuranosidase Producers with High Transfructosylation Activity and its 32 Experimental Run Studies on Reaction Rate of Enzyme. Journal of Biological Sciences, 10: 237-241.
DOI: 10.3923/jbs.2010.237.241
URL: https://scialert.net/abstract/?doi=jbs.2010.237.241
DOI: 10.3923/jbs.2010.237.241
URL: https://scialert.net/abstract/?doi=jbs.2010.237.241
INTRODUCTION
From the last decades functional foods have been described as physiologically active food components, which provide benefits beyond basic nutrition and may prevent disease or promote health. A functional ingredient received much attention in the scientific literature is Fructooligosaccharides (FOS). The FOS is easily understood as inulin-type oligosaccharides of D-fructose, attached by β-(2-1) linkages that carry a D-glucosyl residue at the end of the chain. FOS constitutes a series of homologous oligosaccharides derived from sucrose usually represented in the form of GFn. The GFn units of interest are used as food ingredients (Spiegel et al., 1994) in various nutritional foods. In addition it has important physiological properties, as they are low sweetness, calorie free, act as bifidogenic agent, non cariogenic, reduce cholesterol, phospholipids and triglyceride levels in blood serum, respectively (Yun, 1996; Tokunaga et al., 1986). From the literature survey, FOS producing enzymes are classified as β-Fructofuranosidase (β-D-fructofuranoside fructohydrolase, EC 3.2.1.26; β-FFase) which catalyzes the hydrolysis of sucrose as well as the transfructosylation from sucrose (Kim et al., 1998). Hydrolytic action of β-Fructofuranosidase results in production of invert sugar which is completely healthy sweetener for the preparation of confectionery products like Jams, candies etc. (Klein et al., 1989; Vandamme and Derycke, 1983) and transfructosylation action results in synthesis of FOS which has several applications as mentioned above. Fructosyltransferase (EC 2.4.1.9) is another enzyme possessing only transfructosylating activity which is useful for the production of fructooligosaccharides (Chien et al., 2001). Several research papers reported on FOS production using β-FTases by different strains at high sucrose concentrations (Pencillium citrinium FERMP 15994 (Hayashi et al., 2000); Aspergillus japonicus TIT90076 (Hayashi et al., 1992); Pencillum citrinum KCTC 18080P (Lim et al., 2006). To the best of our knowledge only few reports available on screening of β-Ffase possessing FTase activity (Oskar et al., 2005; Dominguez et al., 2006; Jun et al., 2008). As we all familiar too screening of new strains is complicated and tedious work due to large number of evaluations. Considering above points in this study we reported radial diffusion which is cheap and economically viable technique (mentioned below), to screen potent producers of β-fructofuranosidase. The manuscript also describes the transfructosylation property of β-fructofuranosidase produced by different strains and its effect of physical parameters on the reaction rate of enzyme.
MATERIALS AND METHODS
Materials: Triphenyl Tetrazolium Chloride (TTC), 3,5 Dinitro salicylic acid and Sucrose. All other chemicals used were of analytical grade.
Culture media: The basal culture medium for β-fructofuranosidase production was composed of NH4SO4-0.5 g, KH2PO4-3 g, NaNO3-1.5 g, MgSO4.7H2O-0.02 g, Sucrose-2 g and agar -20 g L-1, pH was adjusted to 7.
Isolation and preliminary screening: Different soil samples were collected from agricultural fields, garden soil and compost soils in and around Northern parts of Karnataka (India) in the year 2008. To isolate β-fructofuranosidase producing fungal strains soil samples were serially diluted and spread on basal culture medium mentioned above and incubated at 30°C for 96 h. Petri Plates were constantly observed for the appearance of colonies with greater diameter and cultures were purified by repeated transfer of cultures and maintained on nutrient agar.
Secondary screening of β-fructofuranosidase producing microbes: The isolated organisms from the preliminary screening were selected and fermented in basal culture media at pH 5.0 excluding agar in 250 mL Erlenmeyer flasks. After incubation on a rotary shaker (30°C, 160 rpm) for 7 days, the culture broth was centrifuged (10000 rpm for 10 min) and the supernatant was collected for enzyme assay.
β-fructofuranosidase assay: Radial diffusion method: It was performed by using TTC reagent (0.1% Triphenyl tetrazolium chloride in 0.5 M NaOH) which easily soluble in water. In the presence of reducing sugars TTC reduce in to triphenyformazon which is a red, water insoluble compound. The quantity of formazan formed is directly proportional to the quantity of reducing sugars present. Wells are made aseptically by using cock borer in the Petri plate containing basal culture media (mentioned above). Two hundred microliter of crude enzyme was loaded in to well and kept for incubation at room temperature for overnight. Staining was carried out by spraying TTC reagent to the agar plate and kept incubation for 20 min in the dark. After washing in the 0.1 M acetate buffer (pH-5), the extra cellular production of β-fructofuranosidase was confirmed by the appearance of the red zone around the well.
Quantitative assay: Hydrolytic and Transfructosylating activities: A 2 mL reaction mixture containing 0.6 mL of 50 mM sodium acetate buffer pH 5.0, 1.0 mL of 5% (w/v) sucrose solution and pre-incubated at 30°C for 10 min. A 0.4 mL of appropriately diluted enzyme was added and the incubation continued for further 30 min at 60°C. The reaction was terminated by inactivating the enzyme keeping in boiling water bath at 100°C for 10 min. After cooling, equal volumes of the reaction mixture was used for determining the reducing sugar and glucose by DNS method (Miller, 1959) and Glucose oxidase-Peroxidase (Wood and Bhat 1988), respectively. The colour developed in both cases was read at 540 nm and converted into reducing sugar and glucose using glucose standard curve prepared under identical conditions. The hydrolytic activity and transfructosylation activity was determined by Nguyen et al. (1999) method.
Determination of protein concentration: Proteins were measured by the Lowry et al. (1951) method using Bovine Serum Albumin (BSA) as the standard protein.
Biomass concentration: Cell mass concentration was determined by dry cell weight per volume (g L-1). The cell mass obtained by filtration of the fermentation broth was washed with distilled water and dried at 120°C for 4 h.
Statistical analysis: All the experiments were repeated at least three times and the results were reproducible. The data points represented the mean values within ±1% of the individual values.
Identification of fungal strains: Analysis of morphology a very small sample of a colony was removed from the agar plate, mixed with a drop of water and smeared on a glass slide (Wei, 1979). Then observations were made with a microscope. For further conformation the strains were stored in PDA (potato dextrose agar) slants at 4°C send to Agarkhar research institute, Pune (India).
Temperature and pH effects on the reaction rate: The temperature and pH effects on the sucrose-fructofuranosidase reaction rate were studied using a 32 experimental design. The proposed experimental design matrix is presented in Table 3. All the experiments were performed by preparing the reaction mixture as mentioned above. The enzyme reaction was carried out for 30 min at pH and temperature to be tested. The reaction rate results of fructose transference (At) and of hydrolysis (Ah) were analyzed mathematically and statistically using Instat Graphpad Software.
RESULTS AND DISCUSSION
Thirty strains that can grow on basal culture medium containing sucrose as sole carbon source were isolated in a pure state. By considering large number of evaluations (growth, size etc.) 16 strains are subjected for secondary screening as mentioned above. Radial diffusion method was found significant to determine the hydrolytic activity of crude enzyme.
| Table 1: | Measurements of zone of inhibition produced by β-fructofuranosidase producers by Radial diffusion method |
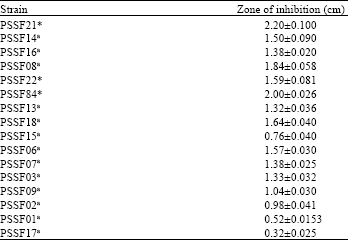 | |
| *Identified strains: PSSF21, Aspergillus niger; PSSF22, Aspergillus flavus; PSSF84, Chrysonilia sitophila. aNon-identified strains | |
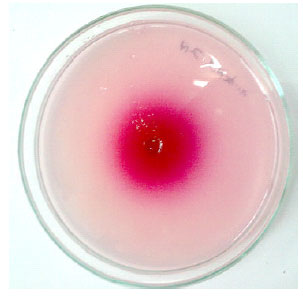 | |
| Fig. 1: | Hydrolysis of sucrose by Aspergillus niger PSSF21 by radial diffusion method |
Activities of β-fructofuranosidase collected from secondary screening were examined to calculate the zone diameter. As shown in Table 1. Zone of hydrolysis produced by strains PSSF21, PSSF84, PSSF08 was found significant comparing with the other isolates producing β-fructofuranosidase (Fig. 1). Based upon morphology and Biochemical characteristics the strains were identified as PSSF21-Aspergillus niger, PSSF84- Chrysonilia sitophila and PSSF22-Aspergillus flavus. Among screened ones PSSF01, PSSF15 and PSSF17 were least producers of β-fructofuranosidase.
In order to determine the relationship between transfructosylation (At) and hydrolytic activity (Ah) the mycelium (ICE) and mycelia broth (ECE) were assayed. The average values of the biomass, enzyme activities of both intracellular (ICE), extra cellular (ECE) and specific activity (SA) were presented in Table 2. The strains PSSF09, PSSF03 PSSF14 reached biomass concentration higher than 7 g L-1. Where as the result with the strains PSSF16, PSSF13, PSSF18, PSSF06 PSSF07, C.sitophila PSSF84, A. flavus PSSF22 and A. niger PSSF21 showed lower biomass concentration (<7 g L-1). Correlation between the biomass concentration and enzyme activity was not found. These results could be caused by the use of different microorganisms that synthesize β-fructofuranosidase with different specific properties and activities, our reports are well in agreement with Hidemasa et al. (1988). In contrast, Fernandez et al. (2007) observed no correlation between cell growth and final pH in shaken flasks.
On the other hand nine strains reached the highest values of extra cellular (ECE) At values which are mentioned in Table 2. Among them A. niger PSSF21 (21.61±0.055 U mL-1), C. sitophila PSSF84 (19.50±0.355 U mL-1), PSSF06 (19.43±0.217 U mL-1) and A. flavusPSSF22 (17.47±0.469 U mL-1) were possessing good transfructosylation activity . But it is known that At/Ah values are the most important parameters to be considered when the objective is screening of strains to produce β-fructofuranosidase with transfructosylation activity (Hidemasa et al., 1988). It is evident from Table 2 significant differences can be observed between the At/Ah ratio values of extra cellular (ECE), intracellular enzyme activities (ECE) and specific activity (S.A). Present observations found similar with previous cited literature (Fernandez et al., 2007). Among twelve screened strains, three of them were selected which shows higher At/Ah values as shown in Table 2. The extra cellular At/Ah ratio of these strains are followed in the order A. niger PSSF21 (11.940 U mL-1) >C. sitophila PSSF84 (8.024 U mL-1) >A. flavusPSSF22 (5.041 U mL-1) were intracellular At/Ah ratio values are in the order C. sitophila PSSF84 (4.042 U mL-1) >A. flavus PSSF22 (3.616 U mL-1) >A. niger PSSF21 (3.038 U mL-1). In contrast PSSF08 strain showed relatively poor At/Ah value (0.657 U mL-1) and high hydrolytic activity (12.84±0.4801 U mL-1) compare to other strains previously listed (Table 2). Lower sucrose concentration might be unfavorable climate to dominate transferase activity in PSSF08 which results in poor At/Ah ratio.
A 32 experimental design was used, to study the effect of pH and temperature on reaction rate of enzyme for three better strains. These strains are selected due to their At/Ah values.
| Table 2: | Biomass, extra cellular, intracellular fructosyltransferase (At) and hydrolytic activities (Ah) reached in shake flask experiments |
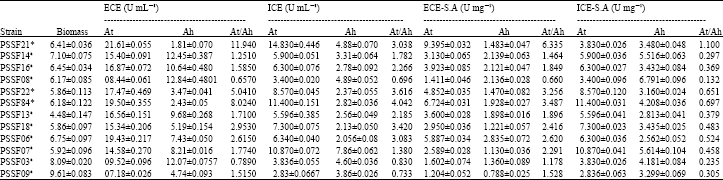 | |
| *Identified strains: PSSF21, Aspergillus niger; PSSF22, Aspergillus flavus; PSSF84, Chrysonilia sitophila. aNon- Identified strains isolated from soil Extra cellular Enzyme = ECE; Intra cellular enzyme = ICE; At = Transfructosylation activity; Ah = Hydrolytic activity; S.A = Specific activity | |
| Table 3: | Effect of physical parameters used to study the β-fructofuranosidase reaction rate by 32 experimental run |
 | |
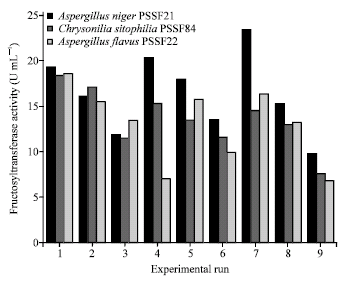 | |
| Fig. 2: | Effects of temperature and pH on fructosyltransferase activity from A. niger PSSF21, C. sitophila PSSF84 and A. flavus PSSF22 |
Experiments were performed under nine conditions using the extra cellular β-fructofuranosidase as described in Table 3. It is evident from Fig. 2. Aspergillus niger PSSF21 achieved higher values of At (23.5±0.75 U mL-1) under experimental conditions of run 7 which indicates good sign of FOS synthesis. Present results are in well agreement with Madlova et al. (1999). Where as the reaction conditions of PSSF84 and PSSF22 strains exhibited higher values of At (18.46±0.537 U mL-1, 18.32±0.285 U mL-1) under conditions of run 1, respectively.
For these microorganisms, the repeated ANOVA was performed (data not shown). The analysis of the obtained results shows that decrease in pH values influences positively on the At value for three strains (A. niger PSSSF21, C. sitophila PSSF84, A. flavus PSSF21), where as temperature effect shown remarkable variation. C. sitophila PSS84 and A. flavus PSSF22 strains showed lower contribution to the At values as the temperature increases and showed positive contribution to Ah values. Concerning the strain A. niger PSSF21 increase in the temperature showed higher At values (mentioned above), besides variation in the pH from 5 to 8 led to decrease in the Ah values and it results remarkable increase in the At/Ah ratio (18.3±0.057 U mL-1) comparing to other two strains (PSSF84 and PSSF22). In contrast Fernandez et al. (2007) reported A. oryzae IPT-301 showed meaningful increase in At/Ah ratio when the variation of pH from 5.5 to 8.0.
CONCLUSION
In view of great demand for FOS production, microbial β-fructofuranosaidase having transfructosylation property has attracted special attention. Researchers showing great interest to survey on new β-fructofuranosidase having unique properties which have ability to synthesis FOS. In this paper, three fungi strains Aspergillus niger PSSF21, Chrysonilia sitophila PSSF84 and Aspergillus flavus PSSF22 have greater potential for FOS production. Considering the results, inspite of non optimization of culture conditions, these microorganisms showed higher values of At/Ah ratio compared to the earlier strains reported. Further studies will be carried out on these strains to meet the demands for FOS production.
ACKNOWLEDGMENTS
The author Praveen Kumar Reddy. Patil would like to thank the Gulbarga University, Gulbarga, India for providing financial support (Junior Research Fellowship) during his work. The author are also grateful to Dr. G. S. N Reddy, Center for Cellular and Molecular Biology, Hyderabad and Dr. Chandrakanth Kelmani, Chairman, Dept of Biotechnology, Gulbarga University, Gulbarga for their constant support and encouragement.
REFERENCES
- Yun, J.W., 1996. Fructooligosaccharides-occurrence, preparation and application. Enzyme Microbial Technol., 19: 107-117.
CrossRefDirect Link - Kim, B.W., J.W. Choi and J.W. Yun, 1998. Selective production of GF4- fructooligosaccharide from sucrose by a new transfructosylating enzyme. Biotechnol. Lett., 20: 1031-1034.
CrossRefDirect Link - Klein, R.D., M.R.Jr. Deibel, J.L. Sarcich, H.A.Z. Neely, I.M. Reardon and R.L. Heinrikson, 1989. Purification and characterization of invertase from a novel industrial yeast Schwanniomyces occidentalis. Prep. Biochem., 19: 283-319.
PubMedDirect Link - Vandamme, E.J. and D.G. Deryke, 1983. Microbial inulinases formentation process, properties and applications. Adv. Applied. Microbiol., 29: 139-176.
PubMed - Chien, C.S., W.C. Lee and T.J. Lin, 2001. Immobilization of A. japonicus by entrapping cells in gluten for production of fructooligosaccharides. Enzyme Microb. Technol., 29: 252-257.
CrossRef - Hayashi, A., K. Matsuzaki, Y. Takasaki, H. Ueno and K. Imada, 1992. Production of β-fructofuranosidase by Aspergillus japonicus. World J. Microbiol. Biotechnol., 8: 155-159.
Direct Link - Lim, J.S., J.H. Lee, J.M. Kim, S.W. Park and S.W. Kim, 2006. Effects of morphology and rheology on neo-fructosyltransferase production by Penicillium citrinum. Biotechnol. Bioproc. Eng, 11: 100-104.
Direct Link - Jun, Y., S. Amachi, H. Shinoyama and T. Fujii, 2008. Production of fructooligosaccharides by crude enzyme preparations of β-fructofuranosidase from Aureobasidium pullulans. Biotechnol. Lett., 30: 535-539.
PubMedDirect Link - Oskar, B., M. del Carmen Ronchel, J.L. Adrio and J. Velasco, 2005. Screening of microorganisms for enzymatic biosynthesis of non digestible oligosaccharides methods in biotechnology. Microb. Enzymes Biotransformation, 17: 105-113.
CrossRef - Dominguez, A., I.M. Santos, A.T. Jose and L. Nelson, 2006. New and Simple plate test for screening relative transfructosylation activity of fungi. Rev. Iberoam Micol., 23: 189-191.
Direct Link - Miller, G.L., 1959. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem., 31: 426-428.
CrossRefDirect Link - Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - Hidemasa, H., H. Masao and S. Naomi, 1988. A fructooligosacharide producing enzyme from Aspergillus niger ATCC 20611. Agric. Boil. Chem., 52: 1181-1187.
Direct Link - Fernandez, R.C., C.A. Ottoni, E.S. da Silva, R.M.S. Matsubara and J.M. Carter et al., 2007. Screening of β-fructofuranosidase-producing microorganisms and effect of pH and temperature on enzymatic rate. Applied Microbiol. Biotechnol., 75: 87-93.
CrossRef