
M.S. Nur Fariza
School of Environmental and Natural Resource Sciences, Faculty of Science and Technology,
Universiti Kebangsaan Malaysia, 43600 Bangi, Selangor Darul Ehsan, Malaysia
S.L. Pang
School of Environmental and Natural Resource Sciences, Faculty of Science and Technology,
Universiti Kebangsaan Malaysia, 43600 Bangi, Selangor Darul Ehsan, Malaysia
C.Y. Choong
School of Environmental and Natural Resource Sciences, Faculty of Science and Technology,
Universiti Kebangsaan Malaysia, 43600 Bangi, Selangor Darul Ehsan, Malaysia
R. Wickneswari
School of Environmental and Natural Resource Sciences, Faculty of Science and Technology,
Universiti Kebangsaan Malaysia, 43600 Bangi, Selangor Darul Ehsan, Malaysia
Journal of Biological Sciences
Year: 2008 | Volume: 8 | Issue: 3 | Page No.: 687-690







ABSTRACT
This study examined DNA sequence variations of coding regions of Cinnamate 4-hydroxylase (C4H) and Cinnamyl Alcohol Dehydrogenase (CAD) in Acacia mangium and A. auriculiformis. cDNA fragments of C4H and CAD with size 1.5 and 1.3 kb, respectively were cloned into pGEM-T Easy Vector and were sequenced. Twenty eight Single Nucleotide Polymorphisms (SNPs) were identified in the coding region of C4H of which 8 caused changes in the amino acids or nonsynonymous mutations and 20 were synonymous mutations. Thirty two SNPs were detected in coding region of CAD. Of these, 12 were nonsynonymous mutations and 20 were synonymous mutations. Two A. mangium individuals (M20 and M22) and two A. auriculiformis individuals (A6 and A3) were used as parents for generating F1 mapping populations. Nucleotide sequence alignment of coding region of CAD detected 28 and 22 SNPs from A3xM22 parental combination and A6xM20 parental combination respectively. Nucleotide sequence alignment of coding region of C4H identified 6 and 23 SNPs for A3xM22 and A6xM20 parental combinations, respectively. For parental combination A3xM22, 11 nonsynonymous mutations were detected while for A6xM20 parental combination 23 nonsynonymous mutations were detected from CAD gene. Amino acid sequence alignment of C4H detected 4 amino acid variations from each parental combination. The putative SNPs can be developed as SNPs markers for Quantitative Trait Loci (QTL) detection. Selecting favourable alleles from progenies which produce desirable lignin profiles would be advantageous in tree breeding programmes for plantation establishment.
PDF Abstract XML References Citation
How to cite this article
M.S. Nur Fariza, S.L. Pang, C.Y. Choong and R. Wickneswari, 2008. Extensive DNA Sequence Variations in Two Lignin Genes, Cinnamate 4-Hydroxylase and Cinnamyl Alcohol Dehydrogenase from Acacia mangium and Acacia auriculiformis. Journal of Biological Sciences, 8: 687-690.
DOI: 10.3923/jbs.2008.687.690
URL: https://scialert.net/abstract/?doi=jbs.2008.687.690
DOI: 10.3923/jbs.2008.687.690
URL: https://scialert.net/abstract/?doi=jbs.2008.687.690
INTRODUCTION
The wood of Acacia mangium and A. auriculiformis is suitable in pulp and paper making as well as for manufacturing of furniture, Medium Density Fiberwood (MDF), veneer and plywood. These species were firstly introduced to Malaysia in 1960s and then have been planted extensively in South East Asia because of their fast growth, good form and economic potential of the wood (Sahri et al., 1993). Spontaneous hybrids of A. mangium and A. auriculiformis were first reported to occur in A. mangium plantations in Sabah (Sim, 1987). Acacia hybrids have shown superior characteristics in growth, adaptation to different types of soils, resistance to disease and higher pulp yield over the parental species (Pinso and Nasi, 1991), showing their importance to the pulp and paper making industry.
Lignin is formed by intracellular synthesis of the monolignol precursors (p?-coumaryl, coniferyl and sinapyl) each giving rise to the hydroxyphenyl (H), guaiacyl (G) and syringyl (S) lignin units respectively. For the production of high quality paper, lignin has to be removed from the cellulose by chemical pulping and the rate of delignification is proportional to the S/G ratio (Piquemal et al., 1998). The benefits of removing as much lignin as possible to avoid residual lignin causes discoloration and reduces paper brightness (Chiang et al., 1988). Cinnamate 4-hydroxylase (C4H) and cinnamyl alcohol dehydrogenase (CAD) are two enzymes that play important role in the beginning and the end of the lignin biosynthesis pathway. C4H catalyzes the hydroxylation of cinnamate to 4-coumarate while the CAD catalyzes the final reduction of cinnamaldehydes to p?-coumaryl, coniferyl and sinapyl alcohols (Lewis, 1999). Down regulation of C4H in Nicotiana tabacum was reported to reduce the lignin content by over 90% and decrease S/G ratio (Blee et al., 2001). The lignin of plants with low CAD activity was more extractable in alkaline chemical pulping process (Baucher et al., 1996). CAD was demonstrated to be a useful target for paper pulping quality improvement in dicotyledons (O`Connell et al., 2002; Pilate et al., 2002).
This study was undertaken to determine the feasibility of using two selected lignin genes, C4H and CAD to find Single Nucleotide Polymorphisms (SNPs) in parental combinations of A. mangium and A. auriculiformis used for generation of segregating mapping populations for fibre length and wood density. Presence of single base differences among the individuals of same species can change the coding of amino acid and protein function resulting in changes in lignin content and composition. SNPs can serve as genetic markers and can be used to characterize genetic resources, map plant genome and specific genes (Dreher et al., 2003). SNP markers have been developed in Eucalyptus nitens and demonstrated that linkage disequilibrium mapping of lignin genes can be used to identify alleles associated with wood quality traits in natural populations (Thumma et al., 2005). SNP discovery and estimation of linkage disequilibrium have been completed in loblolly pine (Pinus taeda) for nineteen candidate genes for wood formation including C4H and CAD (Brown et al., 2004).
MATERIALS AND METHODS
Inner bark tissues of two individuals from A. mangium, M20 and M22 and A. auriculiformis, A3 and A6 were used in this study. Parental combination of A3x M22 and A6xM20 were used in crosses to produce Acacia hybrid mapping population.
Total RNA was extracted from the inner bark tissues using RNeasy Midi Kit (Qiagen, Germany). Total RNA was used as template to synthesize the first strand cDNA and further amplified using a specific primer by reverse transcriptase PCR (RT-PCR) approach. Two primer pairs were designed from full length cDNA of C4H and CAD from Acacia hybrid (Pang et al., 2005). The primer pairs were forward primer (FL-C4H-F: 5` GTC CAC CTT TCT CAG CAG TAT CAA 3`) and reverse primer (FL-C4H-R: 5` CAA TCT CCA AAT CGC CAA CA 3`) to generate the sequence of C4H. CAD was amplified using forward primer (FL-CAD-F: 5` GAC ATT CTT TCT TCT TCT TCT T 3`) and reverse primer (FL-CAD-R: 5` GAC AAA CAT CTG TGA GGC AT 3`). Each reaction consisted of 2 μL of 2.5 p?-mol forward and reverse primers, 1.0 μL of dNTP mix, 2.5 μL of 10xPCR buffer (200 mM Tris-HCl pH 8.8, 100 mM KCl, 1% Triton X-100, 100 mM (NH4)2SO4, 1 μg mL-1 BSA), 0.75 μL of 100 mM MgCl2, 0.04 U YEA Taq DNA Polymerase (Yeastern Biotech Co.) and sterile distilled water to make up 25 μL. C4H amplification was performed in Eppendorf Thermal Cycler (Germany) for 2 min at 95 °C, 35 cycles of 30 sec at 94 °C, 30 sec at 55 °C and 45 sec at 70 °C, followed by final step of 10 min at 70 °C. CAD amplification was carried out by initial denaturing at 95 °C for 2 min, followed by 30 cycles of 94 °C for 30 sec, 46 °C for 30 sec and 70 °C for 45 sec with the final extension time for 10 min.
PCR products were ligated into pGEM-T Easy Vector (Promega) and transformed into competent cell of DH5α from Escherichia coli. Positive clones were cultured in LB Broth supplemented with ampicillin (50 μg μL-1) for overnight. The recombinant plasmids were purified using Wizard Plus SV Minipreps DNA Purification System (Promega). Six to eight positive clones per gene per individual were sent for sequencing using ABI 3730 DNA Analyzer (Applied Biosystem, USA)
The C4H and CAD sequences for the four parental trees (M20, M22, A3, A6) were aligned using the CLUSTALW (http://www.ebi.ac.uk/clustalw/) and visually inspected for base changes. The nucleotide sequence alignments were translated into protein in all reading frames using ORF finder (http://www.ncbi.nlm.nih.gov). The open reading frames of both genes were aligned among four individuals using CLUSTALW to detect the nonsynonymous and synonymous mutations.
RESULTS AND DISCUSSION
cDNA fragments of C4H and CAD of 1.5 and 1.3 kb, respectively (Fig. 1) were cloned into pGEM-T Easy Vector and were sequenced. Six to eight complete cDNA clones for each gene and each individual were sequenced and aligned using CLUSTALW. A3 did not show any nucleotide variation while the others (A6, M20 and M22) showed variations among the clones for C4H. CAD nucleotide sequences showed 20 heterozygous loci which were from A3, M20 and M22. A6 did not show any heterozygous loci.
The consensus sequences of A3, A6, M20 and M22 for both genes were aligned together to detect the SNPs. In total 28 SNPs were found in the coding region of C4H. Of these, 6 SNPs were detected in A3xM22 parental combination while 23 SNPs were detected in A6 x M20 parental combination. Within the coding region of CAD, 32 SNPs were identified. Of these, 28 SNPs were detected in A3xM22 parental combination and 22 SNPs were detected in A6xM20 parental combination. The cDNA sequence of CAD was more polymorphic than that of C4H. On average, one SNP occurred for every 53 bp in C4H compared to one SNP for every 39 bp in CAD (Table 1). The occurrence of SNPs detected in this study
| Table 1: | The number of SNPs in C4H and CAD |
 | |
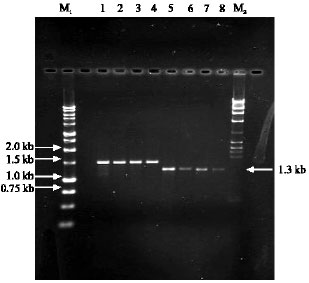 | |
| Fig. 1: | Agarose gel electrophoresis of RT-PCR products of C4H (lane 1-4) and CAD (lane 5-8). M1 is a 1 kb marker and M2 is a Lambda HindIII marker |
for A. mangium and A. auriculiformis are much higher than that detected for coding regions in humans (Cargill et al., 1999), Zea mays (Zhu et al., 2003) and barley (Kanazin et al., 2002). The occurrence of SNPs in Eucalyptus globulus was also reported to be much lower in the coding region of CAD; one SNP occurred for every 147 bp (Poke et al., 2003) compared to that in A. mangium and A. auriculiformis. A. mangium and A. auriculiformis are considered as pioneer species. The high occurrences of SNPs in these species are possible because they have to adapt physiologically to a variety of unpredictable environmental conditions such as infertile acid soils outside their native range. High genetic variation at many physiologically important gene loci may be a strategy for tree species to maintain a higher level of adaptability (Ziehe et al., 1999).
Eight and 12 nonsynonymous mutations were detected in C4H and CAD, respectively (Table 2). Out of these, 4 mutations were detected in A3xM22 parental combination for C4H whereas the remaining 4 mutations were detected in A6xM20 parental combination. Within the open reading frame of CAD, 11 and three nonsynonymous mutations were detected in A3xM22 and A6xM20 parental combination, respectively. The synonymous mutations detected were 20 in both genes.
| Table 2: | The number of synonymous and nonsynonymous mutations detected in C4H and CAD |
 | |
The proportion of nonsynonymous mutations, dN were 0.017 and 0.033 for C4H and CAD, respectively. The proportions of synonymous mutation, dS were 0.042 and 0.056 for C4H and CAD, respectively. The proportion indicates that synonymous mutations were more common than nonsynonymous mutations for both genes. The results were similar to Cargill et al. (1999) and Ramos-Onsins et al. (2008) where the proportions of synonymous mutations were higher than non- synonymous mutations in human coding regions and nine loci involved in the phenylpropanoid pathway of Arabidopsis thaliana, respectively. SNPs of candidate genes for wood properties in P. pinaster. showed one nonsynonymous mutation out of 24 polymorphic sites detected (Pot et al., 2006). Gonzalez-Martinez et al. (2007) also reported that a nonsynonymous substitution for CAD in loblolly pine was in strong association with earlywood specific gravity and causing lignin modification in wild trees.
The putative SNPs can be developed as SNP markers for Quantitative Trait Loci (QTL) detection in Acacia hybrid mapping populations after validation using segregation analysis. A highly polymorphic SNP marker allows the selection of alleles in breeding programmes to produce trees with more favourable lignin profiles for pulping.
ACKNOWLEDGMENTS
This research was supported by funds for postgraduate studies from the Faculty of Science and Technology, Universiti Kebangsaan Malaysia.
REFERENCES
- Baucher, M., B. Chabbert, G. Pilate, J. VanDoorsselaere and M.T. Tollier, 1996. Red xylem and higher lignin extractability by down-regulating a cinnamyl alcohol dehydrogenase in poplar. Plant Physiol., 12: 1479-1490.
Direct Link - Blee, K., J.W. Choi, A.P. O’Connell, S.C. Jupe, W. Schuch, N.G. Lewis and G.P. Bolwell, 2001. Antisense and sense expression of cDNA coding for CYP73A15, a class II cinnamate 4-hydroxylase, leads to a delayed and reduced production of lignin in tobacco. Phytochemistry, 57: 1159-1166.
Direct Link - Brown, G.R., G.P. Gill, R.J. Kuntz, C.H. Langley and D.B. Neale, 2004. Nucleotide diversity and linkage disequilibrium in loblolly pine. Proc. Natl. Acad. Sci. USA., 101: 15255-15260.
CrossRefDirect Link - Cargill, M., D. Altshuler, J. Ireland, P. Sklar, K. Ardlie and N. Patil, 1999. Characterization of single-nucleotide polymorphisms in coding regions of human genes. Nat. Genet., 22: 231-238.
Direct Link - Chiang, V.L., R.J. Puumala, H. Takeuchi and R.E. Eckert, 1988. Comparison of softwood and hardwood kraft pulping. Tappi J., 71: 173-176.
Direct Link - Dreher, K., M. Khairallah, J.M. Ribaut and M. Morris, 2003. Money matters (I): Costs of field and laboratory procedure associated with conventional and marker assisted maize breeding at CIMMYT. Mol. Breed., 11: 221-234.
Direct Link - Gonzalez-Martinez, S.C., N.C. Wheeler, E. Ersoz, C.D. Nelson and D.B. Neale, 2007. Association genetics in Pinus taeda L. I. wood property traits. Genetics, 175: 399-409.
CrossRefDirect Link - Kanazin, V., H. Talbert, D. See, F. DeCamp, E. Nevo and T. Blake, 2002. Discovery and assay of single nucleotide polymorphisms in barley (Hordeum vulgare). Plant Mol. Biol., 48: 529-537.
Direct Link - O’Connell, A., K. Holt, J. Piquemal, J. Grima-Pettenati, A. Boudet, B. Pollet, C. Lapierre, M. Petit-Conil, W. Schuch and C. Halpin, 2002. Improved paper pulp from plants with suppressed cinnamoyl-CoA reductase or cinnamyl alcohol dehydrogenase. Transgenic Res., 11: 495-503.
CrossRefDirect Link - Pilate, G., E. Guiney, K. Holt, M. Petit-Conil and C. Lapierre et al., 2002. Field and pulping performances of transgenic trees with altered lignification. Nat. Biotechnol., 20: 607-612.
Direct Link - Piquemal, J., C. Lapierre, K. Myton, A. O’connell, W. Schuch and J. Grima Pettenati, 1998. Down-regulation of Cinnamoyl-CoA reductase induces significant changes of lignin profiles in transgenic tobacco plants. Plant J., 13: 71-83.
CrossRefDirect Link - Poke, F.S., R.E. Vaillancourt, R.C. Elliot and J.B. Reid, 2003. Sequence variation in two lignin biosynthesis genes, cinnamoyl CoA reductase (CCR) and cinnamyl alcohol dehydrogenase 2 (CAD2). Mol. Breed., 12: 107-118.
CrossRefDirect Link - Pot, D., J.C. Rodrigues, P. Rozenberg, G. Chantre, J. Tibbits, C. Cahalan, F. Pichavant and C. Plomion, 2006. QTLs and candidate genes for wood properties in maritime pine (Pinus pinaster Ait.). Tree Genet. Genomes, 2: 10-24.
Direct Link - Ramos-Onsins, S.E., E. Puerma, D. Balana-Alcade, D. Salguero and M. Aguade, 2008. Multilocus analysis of variation using a large empirical data set: Phenylpropanoid pathway genes in Arabidopsis thaliana. Mol. Ecol., 17: 1211-1223.
Direct Link - Sahri, M.H., F.H. Ibrahim and N.A.A. Shukor, 1993. Anatomy of Acacia mangium grown in Malaysia. IAWA. J., 14: 245-251.
Direct Link - Thumma, B., R., M.F. Nolan, R. Evans and G.F. Moran, 2005. Polymorphisms in Cinnamoyl CoA Reductase (CCR) are associated with variation in microfibril angle in Eucalyptus sp. Genetics, 171: 1257-1265.
CrossRefDirect Link - Zhu, Y.L., Q.J. Song, D.L. Hyten, C.P. Van Tassell, L.K. Matukumalli, D.R. Grimm, S.M. Hyatt, E.W. Fickus, N.D. Young and P.B. Cregan, 2003. Single nucleotide polymorphism in soybean. Genetics, 163: 1123-1134.
Direct Link