
U. Batlang
Department of Crop Science and Production, Faculty of Agriculture, Botswana College of Agriculture, University of Botswana, P/Bag 0027, Gaborone, Botswana
Journal of Biological Sciences
Year: 2008 | Volume: 8 | Issue: 3 | Page No.: 659-662







ABSTRACT
A two year study was conducted to establish effects of a commercial preparation (BA plus GA4+7) called accel, on growth and yield of hot pepper. Five concentrations: 0, 5, 10, 15, 20 and 25 mg L-1 of the preparation were used. Concentrations of 10, 15, 20 and 25 mg L-1 significantly (p<0.001) increased yield in a two year pooled data. Accel treatment did not significantly affect number of fruits, number of branches and plant height. The increase in yield due to accel treatment was associated with significant increase in fruit fresh weight (p<0.001) and length (p<0.001). Therefore the increase in yield was attributed to increase in fruit size. Accel application has the potential to be used as a management practice in greenhouse production of hot pepper.
PDF Abstract XML References Citation
How to cite this article
U. Batlang, 2008. Benzyladenine plus Gibberellins (GA4+7) Increase Fruit Size and Yield in Greenhouse-Grown Hot Pepper (Capsicum annuum L.). Journal of Biological Sciences, 8: 659-662.
DOI: 10.3923/jbs.2008.659.662
URL: https://scialert.net/abstract/?doi=jbs.2008.659.662
DOI: 10.3923/jbs.2008.659.662
URL: https://scialert.net/abstract/?doi=jbs.2008.659.662
INTRODUCTION
Hot pepper (Capsicum annuum L.) (syn.chili pepper) is among the species of the genus Capsicum, domesticated for vegetable and industrial (oleoresin and capsaicin) purposes. Available literature indicates that hot pepper is widely cultivated throughout the world, mainly in Asia, South and North America and parts of Africa (Contreras-Padilla and Yahia, 1998; Kumar et al., 2006). The fresh fruits are used in pickles and sauces, while the red dried and powdered fruits are used as hot spices.
Plant Growth Regulators (PGR) are used extensively in horticulture to enhance plant growth and improve yield by increasing fruit number, fruit set and size. Improvement in vegetative growth and yield attributes may enhance crop productivity. Productivity in horticultural systems is often dependent on manipulation of physiological activities of the crop by chemicals means (Yeshitela et al., 2004) and this is modulated by the interaction of the PGR with plant developmental processes. According to Gianfagna (1987), plant growth regulators can modify development by interfering with biosynthesis, metabolism or translocation of endogenous hormones, or may supplement endogenous hormones when their levels are reduced.
Developmental processes reported to be influenced by PGR application include; induced flowering in Arabidopsis thaliana by gibberellins (Richards et al., 2001), increased flowering in Aquilea species by GA4+7 (Gianfagna and Merrit, 1988), increased fruit fresh weight in cucumber by BA plus GA4+7 in cucumber (Batlang et al., 2006) and GA4+7 (Yang et al., 1992) and stimulation of individual branch growth by BA plus GA4+7 in apple (Voltz et al., 1994). Increase in flowering may lead to more fruits and coupled with increase in fruit size and yield in fruit crops. Fruit size may be determined by increased cell layer formation and cell division and increased sink strength of the fruit (Dyer et al., 1990). Fruit size increase due to cytokinins and gibberellins applications to apples, cucumber and grapes was suggested to be caused by increased cell division and elongation and cell wall extensibility (Emongor and Murr, 2001; Yu et al., 2001).
In the semi-arid areas such as Botswana, there is low rainfall and low fertility and as such there is a need to practice controlled environment horticulture. Intensive production of hot pepper requires the utilization of high yielding cultivars or improved management of the locally available genetic materials. This situation validated the evaluation of accel as a management avenue for intensive production of hot pepper.
MATERIALS AND METHODS
Plant culture: Seeds of a commonly grown landrace were obtained from a farmer in Bobonong (21° 58' S, 28° 27' E), in the Central district of Botswana. The experiments were conducted at Botswana College of Agriculture greenhouses in Gaborone (24° 33' S, 25° 54' E). The landrace has not been genetically characterized and developed into a cultivar and will be referred to as Bob-02; where Bob is Bobonong and 02 is collection year of 2002. Fruits were collected and air dried on a laboratory bench and seeds were removed, washed under a running tap water and germinated in vermiculite in a growth chamber at 25 °C in the dark. After five days uniformly emerged seedlings were transplanted in seedling trays filled with compost soil and grown for four weeks in the greenhouse.
The plants were then transplanted into 8 L polyethene bags filled with soil mixture (4 river sand: 2 top soil and 1 compost) on per volume basis. When plants showed first sings of nutrient deficiency they were irrigated with two hydroponics solutions containing 110 g L-1 hydrogrow (85 g N kg-1, 45 g P kg-1, 240 g K kg-1, 30 g Mg kg-1, 60 g S kg-1, 1.63 g Fe kg-1, 0.4 g Mn kg-1, 0.2 g Zn kg-1, 0.03 g Cu kg-1, 0.05 g Mo kg-1, 0.5 g B kg-1 and 10 g L-1 CaNO3 (26% N) fertilizers. Each fertilizer solution was applied to field capacity at weekly intervals.
Accel treatment: Accel (Abbot Laboratories, North Chicago, IL) is a plant growth regulator that contains the cytokinin 6-benzyladenine (1.8%) and gibberellins (GA4+7) (0.18%). Accel treatment was applied 56 days after transplanting to the vegetatively growing plants. Aqueous solutions were prepared at 0 (control), 5, 10, 15, 20 and 25 mg L-1 of active ingredient (a.i) and applied to run-off with a hand-held sprayer. The control treatments were sprayed with distilled water. Five plants were sprayed for each treatment. Plant became dormant in winter (between May and July) 2003, during which minimal irrigation was applied to carry them through the dormancy period. There was re-growth in the summer beginning August, when hydro-grow and CaNO3 fertilizers regime was resumed, accel application was re-applied at the same rate as in the first year.
Data collection and analysis: Data such as plant height, branch number, marketable fruit number, fruit fresh weight and fruit length were collected between January and May 2003. In this case yield refers to marketable fruits. Fruits were harvested approximately every fourteen days from fruit set until plants ceased to bear fruits. Fruit fresh weight at each time of harvest was divided by the number of fruits to obtain fresh weight per fruit. Data for each parameter was divided by five plants to obtain per plant basis for each level of PGR treatment. In the second year (2003 to 2004) the same data was collected. However, for comparison purposes between years, only data collected between January and May was used for analysis, to minimize seasonal confounding effects.
The experiment was laid out in a randomized complete block design replicated four times. Means were calculated across the five plants in each replication and were subjected to analysis of variance using the general linear models (Proc GLM) of the Statistical Analysis System program (SAS Institute, Carey, NC). Treatment means were separated using the Least Significant Difference (LSD) at p = 0.05. Appropriate linear regression models were used to determine the yield response to fruit number, fruit length and fruit fresh weight per fruit.
RESULTS AND DISCUSSION
It was observed that hot pepper plants did not grow in the winter, even though temperatures were kept at 27 ± 2 °C throughout the experimental period. This observation can be ascribed to the fact that there was low irradiance in winter and high irradiance is required for optimal growth as reported in sweet pepper (Ulvskov et al., 1992). Accel treatment significantly increased yield, fruit fresh weight and fruit length, while plant height, number of branches and fruits were not affected (Table 1). In order to determine how accel might affect chili pepper due to plant age, repeated application of accel was performed on the same plants used in the first year and results were compared. Vegetative growth (plant height and number of branches), fruit number and yield increased with plant age and the increase of the respective parameters was at least 60%. This was to be expected, since in the second year plants had grown more than in the first year and fruiting branches were increased. However, there were no interactions between accel treatments and plant age for all parameters that were assessed (Table 1). The effect of accel treatment and plant age were additive, indicating that the PGR treatment did not affect plant behaviour differently in both years. As a result data was pooled across two years before mean separations.
The PGR treatment at 10, 15, 20 and 25 mg L-1 concentrations increased yield, with 20 and 25 mg L-1 giving highest yield and there were no significant increase in yield between the controls and 5 mg L-1 (Table 2). It was also observed that the increase in yield
| Table 1: | The ANOVA (F-values) of the effects of accel treatment and plant age and their interaction on plant growth, yield and yield components |
 | |
| ***Significant at p<0.001. FW: Fruit Fresh Weight | |
| Table 2: | Effects of accel concentration on yield and its respective components (fruit number, fruit length, fruit fresh weight) in a two year pooled data. |
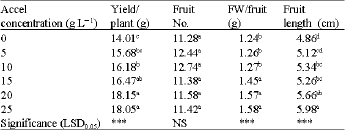 | |
| ***significant at p<0.001. NS: Non-Significant at p = 0.05. FW: Fruit Fresh Weight, Means within columns followed by the same letter(s) are not significantly different at p = 0.05, LSD0.05 | |
| Table 3: | Regression analysis of the relationship between yield and its respective components (fruit number, fruit length, fruit fresh weight) in a two year pooled data |
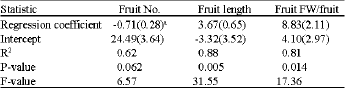 | |
| aValues in parentheses are standard errors | |
due to PGR treatment was generally accompanied with increase in fruit fresh weight per fruit and fruit length, while fruit number was not increased (Table 2, Fig. 1). This is confirmed by regression analyses testing the influence of fruit number and size on yield as a result of PGR treatment (Table 3). It can therefore be reasoned that PGR treatment increased fruit size, which ultimately increased yield on per plant basis. This can be attributed to both cytokinins and gibberellins in their role in cell division and elongation, respectively. The two hormones, which are the components of accel, are known to promote cell division and elongation, respectively (Salisbury and Ross, 1996; Wismer, 1994). During tomato fruit development, there is endogenous gibberellins accumulation, which coincides with activation of cell division and expansion (Gillaspy et al., 1993). The increase in cell volume due to expansion may contribute to the final size of the fruits as observed in this study.
Recently accel was applied to cucumber plants and this caused increased yield through increase in fruit number and size (Batlang et al., 2006). In other fruits such as pear (Stern and Flaishman, 2003) and apple (Wismer et al., 1995; Yuan and Greene, 2000), the increase in fruit size was accompanied by thinning effect due to benzyladenine. While this established the effect of thinning on bigger fruit development, in our case with hot pepper, the increase in fruit size is probably not due to thinning as there was no significant reduction in fruit number due to accel treatment (Table 1, 2). This further implicates the involvement of cytokinins and gibberellins in accel on fruit development in hot pepper.
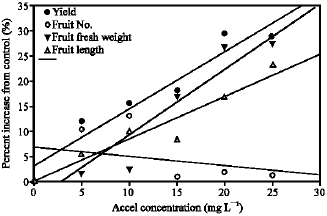 | |
| Fig. 1: | Effects of accel concentration on the increase of yield and its component |
Foliar application of both benzyladenine and GA3 to citrus was reported to enhance assimilate (sucrose) export by the foliage to the developing fruit and GA3 was especially active in promoting fruit sink activity (Mauk et al., 1986). Exogenous application of benzyladenine and GA3 increased expansion of excised leaf discs in sweet pepper (Nielsen and Ulvskov, 1992; Ulvskov et al., 1992). According to Yuan and Greene (2000) application of benzyladenine to apples increased levels of zeatin riboside in fruits, suggesting that it might promote cell division through the activity of zeatin riboside, which is one of the dominant endogenous cytokinins found in pepper (Ulvskov et al., 1992).
Although no attempts have been made to measure cell division in this experiment, it is hypothesized that accel influenced yield in hot pepper directly through processes that increased cell division and improved fruit sink capacity. The next experiments will address direct involvement of accel on fruit growth and development in hot pepper.
ACKNOWLEDGMENTS
The author thanks Botswana College of Agriculture, Department of Crop Science and production for financial and material support and Ms. Biganani Chalegwa for her invaluable technical assistance. Thanks are due to Motshwari Obopile for his critical review of the Manuscript.
REFERENCES
- Batlang, U., V.E. Emongor and F. Pule-Meulenberg, 2006. Effect of benzyladenine plus gibberellic acid on yield and yield components of cucumber (Cucumis sativus L. cv. tempo). J. Agron., 5: 418-423.
CrossRefDirect Link - Contreras-Padilla, M. and E.M. Yahia, 1998. Changes in capsaicinoids during development, maturation and senescence of chile peppers and relation with peroxidase activity. J. Agric. Food Chem., 46: 2075-2079.
CrossRefDirect Link - Gianfagna, T.J. and R.H. Merrit, 1988. GA4+7 promotes stem growth and flowering in a genetic line of Aquilegia x hybrida Sims. Plant Growth Regul., 24: 1-5.
CrossRefDirect Link - Gillaspy, G., H. Ben-David and W. Gruissem, 1993. Fruit: A developmental perspective. Plant Cell, 5: 1439-1451.
Direct Link - Kumar, S., S. Kumar, M. Singh, A.K. Singh and M. Rai, 2006. Identification of host plant resistance to pepper leaf curl virus in chilli (Capsicum species). Scient. Hortic., 110: 359-361.
CrossRefDirect Link - Mauk, C.S., M.G. Bausher and G. Yelenosky, 1986. Influence of growth regulator treatments on dry matter production, fruit abscission and 14C-assimilate partitioning in citrus. J. Plant Regul., 5: 111-120.
CrossRefDirect Link - Nielsen, T.H. and P. Ulvskov, 1992. Cytokinins and leaf development in sweet pepper (Capsicum annuum L.) II. Sink metabolism in relation to cytokinin-promoted leaf expansion. Planta, 188: 78-84.
CrossRef - Richards, D.E., K.E.T. Ait-Ali and N.P. Harberd, 2001. How gibberellin regulates growth and development: A molecular genetic analysis of gibberellin signaling. Annu. Rev. Plant Physiol. Plant Mol. Biol., 52: 67-88.
Direct Link - Stern, R.A. and M.A. Flaishman, 2003. Benzyladenine effects on fruit size, fruit thinning and return yield of Spadona and Coscia pear. Sci. Hort., 98: 499-504.
Direct Link - Ulvskov, P., T.H. Nielsen, P. Seiden and J. Marcussen, 1992. Cytokinins and leaf development in sweet pepper (Capsicum annuum L.) I. Spatial distribution of endogenous cytokinins in relation to leaf growth. Planta, 188: 70-77.
CrossRef - Volz, R.K., H.M. Gibbs and J. Popenoe, 1994. Branch induction on apple nursery trees: Effects of growth regulators and defoliation. N. Z. J. Crop Hortsci., 22: 277-283.
CrossRefDirect Link - Wismer, P.T., J.T.A. Proctor and D.C. Elfving, 1995. Benzyladenine affects cell division and cell size during apple fruit thinning. J. Am. Soc. Hortsci., 120: 802-807.
Direct Link - Yang, Y., D. Lin and Z. Guo, 1992. Promotion of fruit development in cucumber (Cucumis sativus) by thidiazuron. Sci. Hort., 50: 47-51.
CrossRef - Yeshitela, T., P.J. Robbertse and P.J.C. Stassen, 2004. Paclobutrazol suppressed vegetative growth and improved yield as well as fruit quality of Tommy Atkins mango (Mangifera indica) in Ethiopia. N. Z. J. Crop Hortsci., 32: 281-293.
Direct Link - Yu, J.Q., Y. Li, Y.R. Qian and Z.J. Zhu, 2001. Cell division and cell enlargement in fruit of Lagenaria leucantha as influenced by pollination and plant growth substances. Plant Growth Regul., 33: 117-122.
Direct Link - Yuan, R. and D.W. Greene, 2000. McIntosh apple fruit thinning by benzyladenine in relation to seed number and endogenous level in fruit and leaves. Sci. Hort., 86: 127-134.
Direct Link