
B. Siti Balkis
Biomedical Department, Faculty of Allied Health Sciences, Universiti Kebangsaan Malaysia, Jalan Raja Muda Abdul Aziz, Kuala Lumpur, 50300, Malaysia
R. Nor Fadilah
Department of Forensic Science, Faculty of Allied Health Sciences, Universiti Kebangsaan Malaysia, Kuala Lumpur, 50300, Malaysia
A. Ezlan
Biomedical Department, Faculty of Allied Health Sciences, Universiti Kebangsaan Malaysia, Jalan Raja Muda Abdul Aziz, Kuala Lumpur, 50300, Malaysia
O. Khairul
Department of Forensic Science, Faculty of Allied Health Sciences, Universiti Kebangsaan Malaysia, Kuala Lumpur, 50300, Malaysia
A.B. Mokhtar
Department of Pathology, Faculty of Medicines, Universiti Kebangsaan Malaysia, Kuala Lumpur, 50300, Malaysia
M. Jamaludin
Biomedical Department, Faculty of Allied Health Sciences, Universiti Kebangsaan Malaysia, Jalan Raja Muda Abdul Aziz, Kuala Lumpur, 50300, Malaysia
Journal of Biological Sciences
Year: 2008 | Volume: 8 | Issue: 3 | Page No.: 622-627







ABSTRACT
The aim of present study was to investigate the effect of alpha lipoic acid (ALA) supplementation on oxidative stress and DNA damage in streptozotocin-induced diabetic rats. The diabetic rats were divided into two groups; supplemented with ALA (100 mg/kg/day) and non-supplemented with ALA (No Suppl group). Non-diabetic rats (NDM) formed the control group. Following eight weeks of supplementation, fasting blood glucose (FBG) and HBA1c in ALA-supplemented rats was found to be significantly lower than the No Suppl diabetic rats. ALA supplementation also significantly increased plasma superoxide dismutase (SOD) activity and vitamin C level compared to the No Suppl group. The increased in plasma malondealdehyde (MDA) and 4-hydroxynonenal (4-HNE) levels also were inhibited. At the same time, the percentages of comet tail and tail moment of peripheral lymphocytes which indicate the oxidative DNA damage were significantly reduced in the ALA-supplementation group. These results suggest that ALA has protective effects against oxidative stress by increasing antioxidant status and reducing lipid peroxidation and DNA damage in streptozotocin-induced diabetic rats.
PDF Abstract XML References Citation
How to cite this article
B. Siti Balkis, R. Nor Fadilah, A. Ezlan, O. Khairul, A.B. Mokhtar and M. Jamaludin, 2008. Oxidative Stress and DNA Damage in Streptozotocin-Induced Diabetic Rats: Effects of Alpha Lipoic Acid. Journal of Biological Sciences, 8: 622-627.
DOI: 10.3923/jbs.2008.622.627
URL: https://scialert.net/abstract/?doi=jbs.2008.622.627
DOI: 10.3923/jbs.2008.622.627
URL: https://scialert.net/abstract/?doi=jbs.2008.622.627
INTRODUCTION
Diabetes Mellitus (DM) is a metabolic disorder characterized by hyperglycemia resulting from defective insulin secretion, resistance to insulin action or both (Altan, 2003). DM causes alteration in carbohydrate, fats and protein metabolism and increased risk of cardiovascular disease. It is associated with reduced quality of life and increased rate of mortality and morbidity. The cost of caring for patients with DM and its complications will continue to grow with the increasing incidence of the disease (Cefalu, 2004).
Oxidative stress has been suggested as a potential contributor to the development of complications as a results of DM. Growing evidence indicates that oxidative stress is increased in diabetes due to the overproduction of Reactive Oxygen Species (ROS) and decreased efficiency of antioxidant defenses as a results of hyperglycemia (Wiernsperger, 2003). Oxidation of lipids, proteins and other macromolecules such as DNA occurs during the development of diabetes and its complications (Stetina et al., 2006). Hyperglycemia leads to the increased production of free radical, lipid peroxidation products, DNA damage and mutations that contribute to diabetes, cancer and other age-related diseases (Evans et al., 2004).
To control the flux of ROS, aerobic cells have developed an antioxidant defense system which includes enzymatic and non-enzymatic components (Ahmed, 2005). The stability and capacity of antioxidant status during chronic diabetes significantly influences the outcome of the long-term complications caused by oxidative stress (Sasvári and Nyakas, 2003). Alpha Lipoic Acid (ALA) is a naturally occurring short chain fatty acid with sulfhydryl groups that has potent antioxidative activity in a wide variety of experimental systems and is clinically used to treat diabetic neuropathy (Packer et al., 2001). ALA is a unique antioxidant because it has beneficial effects on fuel metabolism and also an essential cofactor of mitochondrial respiratory enzymes, including the pyruvate dehydrogenase (PDH) complex (Packer et al., 2001).
Recently, the use of dietary antioxidants has become a popular approach in preventing DM and to attenuate the complications due to hyperglycemia. Various antioxidants may be helpful in the reduction of oxidative stress and protection of cells against diabetes-induced injury (Karasu et al., 1997). Therefore, the aim of this study was to evaluate the potential of ALA in preventing oxidative damage in DM. We investigated the effects of oral supplementation of ALA on glycaemic status, oxidative stress levels and DNA damage on streptozotocin (STZ)-induced diabetic rats.
MATERIALS AND METHODS
Diabetes induction: Male Spraque Dawley rats weighing 260-290 g were supplied by the Animal Unit of University Kebangsaan Malaysia which is located at the Institute of Medical Research, Kuala Lumpur, Malaysia. This study was conducted from January to December 2003. Animals were housed two per cage. All animals were maintained on a balanced diet and given water ad libitum without restricted. The study was approved by University Kebangsaan Malaysia Animal Ethics Committee (UKMAEC) no; Biomed/2001/Siti/17-May/047 and UKMAEC guidelines on using live animals were followed.
Diabetes was induced in a group of rats after an overnight fast by single intravenous injection (via tail vein) of STZ (Sigma, St Louis, MO, USA), which was freshly dissolved in normal saline. The dose was 50 mg/kg body weight. Another group of rats, which only received saline, formed the non-DM group (NDM) (n = 7). Three days later, blood was collected via tail vein and glucose concentration measured by a strip-operated blood glucose sensor (Companion 2, Medisense Ltd., Birmingham, UK). Rats with blood glucose levels >15.0 mmol L-1 were included in the study and divided into two groups; supplemented with ALA (n = 8) and not supplemented with ALA (No Suppl) (n = 8). ALA (Sigma, St Louis, MO, USA) was administered orally at a dose of 100 mg/kg body weight/day throughout the feeding period of eight weeks and the supplementation was begun on the same day. The treatment dose of ALA was based on a previous study (Arivazhagan and Panneerselvam, 2000). No Suppl and NDM rats were left untreated.
Biochemical analysis: After eight weeks of supplementation, rats were fasted overnight and blood was collected by cardiac puncture under deep anesthesia with diethyl ether. Blood was collected into tubes containing sodium fluoride tube (fasting blood glucose (FBG) analysis) and EDTA.
Plasma glucose levels were analyzed using enzymatic glucose-oxidase kits (Trace Scientific, Melbourne, Australia. Catalogue no TR 15104). Blood HbA1c was determined using method described by Eross et al. (1984) and express as percent of total hemoglobin. Plasma protein concentrations were measured by the method of Bradford (1976) and MDA+HNE levels were assayed by lipid peroxidation using a kit (Calbiochem`s Lipid Peroxidation Assay, catalogue No. 437634). SOD activities in plasma were determined according to Beyer and Fridovich (1987). In this method, SOD activities were measured by the photochemical nitro blue tetrazolium (NBT) reduction; one unit of SOD is the amount which causes a 50% decrease of the SOD-inhibitable NBT reduction.
Plasma vitamin C was measured according to the method of Liau et al. (1993). Briefly, 100 μL plasma was mixed with 100 μL of 10% perchloric acid containing 1% of meta-phosphoric acid. All were mixed in 1.5 eppendorf tubes, wrapped in aluminum foil to ensure the vitamin C would not be degraded by light and kept at -40 °C. Vitamin C analysis was done within one week of storage using high-performance liquid chromatography (HPLC) at room temperature. The HPLC system consisted of a Gilson 307 pump, Rheodyne manual injector, 20 μL sample loop and a Gilson 151 UV/VIS set at 245 nm. Column for separation was a Metaphase Crestpak C18S (4.6 mm ID x 150 mm L-1). Mobile phase consisted of 0.015% metaphosphoric acid in 20 mM ammonium dihydrogen phosphate (pH 2.95). Flow rate was set at 1 mL min-1 with a run time of 10 min. Eluted peak of vitamin C was detected using UV/VIS Gilson set at 245 nm and results were analyzed based on the area under the peak.
Prior to injection of the sample into the system, 200 μL HPLC mobile phase was added to sample, vortex and centrifuged at 20,000 g for 1 min. A total of 10 μL supernatant was then injected into the HPLC for vitamin C analysis. Standard curve to extrapolate true concentration of vitamin C in sample was done daily by diluting 10 mg L-(+)-ascorbic acid (BDH, Poole, UK) in 10 mL fresh mobile phase. The mixed solution was then diluted further with 1% metaphosphoric acid and 10% perchloric acid to produce a stock solution with a concentration of 100 mg mL-1. External control of analysis was done by injecting a vitamin C standard for every 10 injections. An external control concentration deviation of 5% will result in the creation a new standard curve.
Alkaline comet assay: The Alkaline Comet Assay procedure was done as described previously by Singh et al. (1988). Electrophoresis slides observation were made using Leitz Laborlux fluorescence microscope (Nikon) equipped with an epifluorescence mercury lamp source (excitation filter 515, barrier filter 590 nm) and X40 fluorescence objective (numerical aperture 0.85) and data were analyzed using a specialized single-cell gel (SCG) image analysis programme (TriTex Comet ScoreTM (free comet score)).
The image analysis software provided a full range of densitometric and geometric parameters describing the complete comet, as well as the head and tail of DNA portions. Since the comet assay essentially reflects the displacement of fluorescence from the head to the tail in damage cells, we used % tail DNA (comet tail), i.e. the percentage of total nuclear DNA that has migrated to the tail and tail moment (% tail DNA x length) as the parameter to quantify basal levels of DNA damage. Each slide was analyzed in duplicate and 50 cells per slide were scored.
Statistical analysis: All results were expressed as mean ± standard error of the mean (SEM). The significance of the difference between the means of the different groups was analysed using one-way analysis of variance (ANOVA), followed by post hoc LSD test for multiple comparisons. The difference between groups was considered significant when p<0.05.
RESULTS AND DISCUSSION
FBG and HbA1c levels for all groups are shown in Table 1. FBG and HbA1c levels were found to be higher in the No Suppl and ALA groups than in the NDM group. FBG and HbA1c levels in the ALA group were significantly lower compared to the No Suppl diabetic group.
MDA+HNE levels of plasma were significantly increased in No Suppl diabetic group compared with NDM group. Treating diabetic rats with ALA normalized plasma MDA+HNE levels. STZ-induced diabetic rats also had a significant decreased in mean plasma SOD activities as well as in mean plasma vitamin C levels compared with NDM group. However, plasma SOD activities in ALA group were significantly higher compared to No Suppl group. ALA supplementations also inhibited the reduction of plasma vitamin C in diabetic rats (Table 2).
The images of lymphocyte DNA after the Comet Assay are shown in Fig. 1. The mean level of endogenous DNA damage, measured by the % tail DNA (comet tail) of the lymphocytes was significantly higher in Non Suppl rats compared with NDM rats (Table 3). ALA
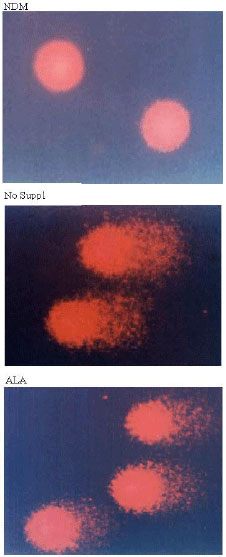 |
| Fig. 1: | Images of comets in different groups of study after eight weeks of study |
| Table 1: | Comparison of FBG and HbA1c levels in the NDM, No Suppl and ALA groups after 8 weeks of study |
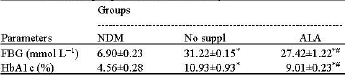 | |
| Data are expressed as the mean ± SEM. *p<0.05 vs NDM group, #p<0.05 vs No Suppl group | |
supplementation normalized the % tail DNA in ALA diabetic group. Tail moment, which is defined as the product of the tail length and the fraction of DNA in the tail was also used as a parameter to describe the extent of DNA damage. Diabetic rats showed significantly higher levels of tail moment compared to NDM rats. ALA supplementation was also able to normalize the tail moment value (Table 3).
| Table 2: | Comparison of plasma MDA+HNE levels, SOD activities and Vitamin C levels in the NDM, No Suppl and ALA groups after 8 weeks of study |
 | |
Data are expressed as the mean ± SEM. *p<0.05 vs NDM group, #p<0.05 vs No Suppl group | |
| Table 3: | Comparison of tail DNA and tail moment levels in the NDM, No Suppl and ALA groups after 8 weeks of study |
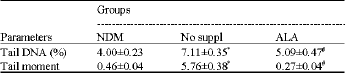 | |
| Data are expressed as the mean ± SEM. *p<0.05 vs NDM group, #p<0.05 vs No Suppl group | |
In the present study, blood glucose levels in STZ-induced diabetic rats were significantly higher compared to control rats. Oral administration of ALA showed hypoglycemic effects against STZ-induced diabetes in rats. ALA supplementation lowered significantly the levels of FBG and HbA1c. These observations are consistent with previous findings (Nagamatsu et al., 1995; Obrosova et al., 1998). Our results support an earlier finding that antioxidant supplementation has beneficial effects in decreasing blood glucose concentration, promoting regeneration of the pancreatic islets and increasing insulin release in STZ-induced diabetic rats (Vessal et al., 2003).
Previous researchers had shown that STZ-diabetes leads to oxidative stress and abnormal antioxidant status (Kamalakkannan and Stanely Mainzen Prince, 2003). In the present study, we observed elevated levels of lipid peroxidation product in plasma of STZ-induced diabetic rats. Elevated levels of MDA+HNE in the plasma are consequences of increased production and liberation of lipid peroxides due to pathological changes (Kamalakkannan and Prince, 2003).
ALA is the most important lipid and water-soluble radical scavenging antioxidant. ALA reduces lipid hydroperoxides generated during the process of peroxidation and protects the cell structure against damage (Packer et al., 2001). Exogenous ALA is reduced intracellular and this form influences a number of cell processes. ALA is capable of scavenging free radicals such as hydroxyl radicals, hypochlorous acid and singlet oxygen (Packer et al., 2001). MDA+HNE levels were significantly decreased in plasma of ALA-supplemented diabetic rats compared to diabetic rats without any supplementation. Further, no significant differences were found compared with the NDM group. This suggests that ALA is capable of preventing the process of lipid peroxidation that normally occurs in diabetic condition. The ability of ALA to decrease lipid peroxidation process confirmed previous reports (Cakatay et al., 2000; Obrosova et al., 2000).
SOD is powerful enzymatic antioxidant that inhibits lipid peroxidation by inactivating the superoxide radicals. Reduced plasma SOD activities in the present study may be due to elevated free radicals. Similar finding was observed in experimental diabetes (Mohamad et al., 2004). Vitamin C is an excellent hydrophilic antioxidant in plasma because it disappears faster than other antioxidant when plasma is exposed to reactive oxygen species (Frei et al., 1989). The observed decrease in plasma vitamin C may be due to increased utilization as an antioxidant defence against increased reactive oxygen species. A second possibility is because of reduction in other antioxidant enzyme such as reduced glutathione (GSH) which is required for the recycling of vitamin C. Even though this study did not measure GSH levels, a previous study was reported that the reduction of GSH occurred in STZ-induced diabetes (Wefers and Sies, 1988).
The elevated levels of vitamin C and SOD activities in ALA group play a protective role against increased peroxidation in diabetes. ALA replenishes vitamin C, glutathione and vitamin E through the reduction of their radicals via the redox cycle (Packer et al., 2001). This observation showed that ALA could improve the antioxidant defence mechanism by reducing free radicals levels and thereby reducing the oxidative damage that occurs in diabetic condition.
In the present study, we used the Alkaline Comet Assay to measure basal levels of DNA damage in freshly isolated lymphocyte. There were significant increases in the levels of DNA damage in the lymphocyte from No Suppl diabetic rats. Our results also indicated that MDA levels were significantly higher in Non Suppl diabetic rats. These findings show that DNA damage is associated with the production of free radicals. Free radicals including hydroxyl radicals, singlet oxygen, peroxyl radicals and peroxynitrite are able to produce modifications to the DNA bases, strand breakage and various other DNA damages (Imaeda et al., 2002a).
Prevention in DNA damage was also observed in diabetic rats after administration of ALA. This finding is in agreement with the previous study which found ALA has the capability in preventing DNA damage induced by palmitic acid (Beeharry et al., 2003). ALA also has the ability to prevent apoptosis process. ALA is functionally efficient in helping cells to recover from oxidative damage (Arivazhagan and Panneerselvam, 2000). Supplementation of ALA had caused a substantial increase in nucleic acid content and protein in aged rats. Previous study also showed that antioxidants such as fluvastatin (Imaeda et al., 2002b) and vitamin E (Claycombe and Meydani, 2001) have protective effect against oxidative DNA damage. The presence of antioxidants can influence the reaction of the cells to external mutagens and also the ability to detoxify the mutagens (Claycombe and Meydani, 2001). Since ALA is an effective antioxidant the protective effect of ALA on oxidative stress induced DNA damage may be mediated through inhibition of free radical formation and also action as a free radical scavenger.
Previous studies have demonstrated a relationship between chronic hyperglycaemia and the development of secondary complications in diabetes patients. It has been postulated that oxidative stress due to chronic hyperglycaemia may play an important role in etiology of diabetic complications (Seghrouchni et al., 2002; Martin-Gallan et al., 2003). In the present study the antioxidant properties of ALA seem to be major contributing factors in reducing oxidative stress and DNA damages. Since glycemic control plays an important role in the progression of diabetes mellitus and the development of secondary complications, the reduction of oxidative stress in ALA supplementation diabetic rats may be associated with the improvement in glycemic index.
In conclusion, ALA may have the potential to reduce oxidative stress and inhibits DNA damage in diabetic rats by improving glycemic status and providing antioxidant activities.
ACKNOWLEDGMENTS
This study was supported by Grants from Malaysian Ministry of Science and Technology (IRPA Grant No. 06-02-02-0111 and Science Fund Grant No. 05-01-02-SF 0014). We thank Prof. K.L. Yap and Ms Hafiza Ariff for helping us editing this manuscript.
REFERENCES
- Ahmed, R.G., 2005. The physiological and biochemical effects of diabetes on the balance between oxidative stress and antioxidant defense system. Med. J. Islamic World Acad. Sci., 15: 31-42.
Direct Link - Altan, V.M., 2003. The pharmacology of diabetic complications. Curr. Med. Chem., 10: 1317-1327.
CrossRef - Arivazhagan, P. and C. Panneerselvam, 2000. Effect of DL-α-lipoic acid on tissue nucleic acid contents in aged rats. Pharmacol. Res., 42: 223-226.
CrossRefDirect Link - Beeharry, N., J.E. Lowe, A.R. Hernandez, J.A. Chambers and F. Fucassi et al., 2003. Linoleic acid and antioxidants protect against DNA damage and apoptosis induced by palmitic acid. Mutat. Res., 530: 27-33.
CrossRefDirect Link - Beyer, Jr. W.F. and I. Fridovich, 1987. Assaying for superoxide dismutase activity: Some large consequences of minor changes in conditions. Anal. Biochem., 161: 559-566.
CrossRefDirect Link - Bradford, M.M., 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem., 72: 248-254.
CrossRefPubMedDirect Link - Cakatay, U., A. Telci, S. Salman, I. Satman and A. Sivas, 2000. Oxidative protein damage in type 1 diabetic patients with and without complication. Endocr. Res., 26: 365-379.
Direct Link - Cefalu, W., 2004. Economic of diabetes-cost impact of not treating diabetes early and intensively. Clin. Cornerstone, 6: 51-60.
PubMed - Claycombe, K.J. and S.N. Meydani, 2001. Vitamin E and genome stability. Mutat. Res., 475: 37-44.
Direct Link - Eross, J., D. Kreutzmann, M. Jimenez, R. Keen, S. Roger, C. Cowell, R. Vines and M. Silink, 1984. Colometric measurement of glycosylated protein in whole blood, red blood cells, plasma and dried blood. Ann. Clin. Biochem., 21: 477-483.
PubMed - Evans, M.D., M. Dizdaroglu and M.S. Cooke, 2004. Oxidative DNA damage and disease: Induction, repair and significance. Mutat. Res./Rev. Mutat. Res., 567: 1-61.
CrossRefDirect Link - Frei, B., L. England and B.N. Ames, 1989. Ascorbate is an outstanding antioxidant in human blood plasma. Proc. Natl. Acad. Sci. USA., 86: 6377-6381.
CrossRefPubMedDirect Link - Imaeda, A., T. Kaneko, T. Aoki, Y. Kondo, N. Nakamura, H. Nagase and T. Yoshikawa, 2002. Antioxidative effects of fluvastatin and its metabolites against DNA damage in streptozotocin-treated mice. Food Chem. Toxicol., 40: 1415-1422.
Direct Link - Imaeda, A., T. Kaneko, T. Aoki, Y. Kondo and H. Nagase, 2002. DNA damage and the effect of antioxidants in streptozotocin-treated mice. Food Chem. Toxicol., 40: 979-987.
Direct Link - Kamalakkannan, N. and P.S.M. Prince, 2003. Hypoglycaemic effect of water extracts of Aegle marmelos fruits in streptozotocin diabetic rats. J. Ethnopharmacol., 87: 207-210.
CrossRefDirect Link - Karasu, C., G. Ozansoy, O. Bozkurt, D. Erdogan and S. Omeroglu, 1997. Antioxidant and triglyceride-lowering effects of vitamin E associated with the prevention of abnormalities in the reactivity and morphology of aorta from streptozotocin-diabetic rats. Metabolism, 46: 872-879.
CrossRefDirect Link - Liau, L.S., B.L. Lee, A.L. New and C.N. Ong, 1993. Determination of plasma ascorbic acid by high-performance liquid chromatography with ultraviolet and electrochemical detection. J. Chromatogr., 612: 63-70.
CrossRefDirect Link - Martin-Gallan, P., A. Carrascosa, M. Gussinye and C. Dominguez, 2003. Biomarkers of diabetes-associated oxidative stress and antioxidant status in young diabetic patients with or without subclinical complications. Free Radic. Biol. Med., 34: 1563-1574.
CrossRefPubMedDirect Link - Mohamad, S., A. Taha, R.N.K. Bameizai, S.F. Basir and N.Z. Baquer, 2004. Lower doses of vanadate in combination with trigonella restore altered carbohydrate metabolism and antioxidant status in alloxan-diabetic rats. Clin. Chim. Acta, 342: 105-114.
CrossRefPubMedDirect Link - Nagamatsu, M., K.K. Nickander, J.D. Schmelzer, A. Raya, D.A. Wittrock, H. Tritschler and P.A. Low, 1995. Lipoic acid improves nerve blood flow, reduces oxidative stress and improves distal nerve conduction in experimental diabetic neuropathy. Diabetes Care, 18: 1160-1167.
CrossRefDirect Link - Obrosova, I., X. Cao, D.A. Greene and M.J. Stevens, 1998. Diabetes-induced changes in lens antioxidant status, glucose utilization and energy metabolism: Effect of DL-α-lipoic acid. Diabetologia, 41: 1442-1450.
CrossRefDirect Link - Obrosova, I.G., L. Fathallah and D.A. Greene, 2000. Early changes in lipid peroxidation and antioxidative defense in diabetic rat retina: Effect of DL-alpha-lipoic acid. Eur. J. Pharmacol., 398: 139-146.
Direct Link - Packer, L., K. Kraemer and G. Rimbach, 2001. Molecular aspects of lipoic acid in the prevention of diabetes complications. Nutrition, 17: 888-895.
CrossRefDirect Link - Sasvári, M. and C. Nyakas, 2003. Time dependent changes in oxidative metabolism during chronic diabetes in rats. Acta Biol. Szegediensis, 47: 153-158.
Direct Link - Seghrouchni, I., J. Drai, E. Bannier, J. Riviere and P. Calmard et al., 2002. Oxidative stress parameters in type 1, type 2 and insulin-retreated type 2 diabetes mellitus: Insulin treatment efficiency. Clin. Chim. Acta, 321: 89-96.
Direct Link - Singh, N.P., M.T. McCoy, R.R. Tice and E.L. Schneider, 1988. A simple technique for quantitation of low levels of DNA damage in individual cells. Exp. Cell Res., 175: 184-191.
CrossRefPubMedDirect Link - Stetina, R., J. Varvarovska, Z. Rusavy, R. Pomahacova and J. Racek et al., 2006. Oxidative stress, DNA damage and DNA repair capacity in children with type 1 diabetes mellitus. Toxicol. Lett., 164: S1-134.
CrossRefDirect Link - Vessal, M., M. Hemmati and M. Vasei, 2003. Antidiabetic effects of quercetin in Streptozocin-induced diabetic rats. Comparat. Biochem. Physiol. Part C. Toxicol. Pharmacol., 135: 357-364.
CrossRefPubMedDirect Link - Wefers, H. and H. Sies, 1988. The protection by ascorbate and glutathione against microsomal lipid peroxidation is dependent on vitamin E. Eur. J. Biochem., 174: 353-357.
CrossRefPubMedDirect Link - Wiernsperger, N.F., 2003. Oxidative stress as a therapeutic target in diabetes: Revisiting the controversy. Diabetes Metab., 29: 579-585.
CrossRefDirect Link