
Amandeep Kaur
Department of Molecular Biology and Biochemistry, Guru Nanak Dev University, Amritsar-143 005, Punjab, India
Rajinder Singh
Department of Chemistry, University of Cape Town, Rondebosch 7701, South Africa
Sukhdev Singh Kamboj
Department of Molecular Biology and Biochemistry, Guru Nanak Dev University, Amritsar-143 005, Punjab, India
Jatinder Singh
Department of Molecular Biology and Biochemistry, Guru Nanak Dev University, Amritsar-143 005, Punjab, India
Girish J. Kotwal
Division of Medical Virology, Institute of Infectious Diseases and Molecular Medicine, University of Cape Town, HSC, Cape Town, 7925, South Africa
Journal of Biological Sciences
Year: 2008 | Volume: 8 | Issue: 7 | Page No.: 1236-1240







ABSTRACT
A monocot mannose-binding Amaryllidaceae lectin from the bulbs of Crinum latifolium, specific for methyl-α-D-mannopyranoside, has earlier been reported as a nonmitogenic, nonglycosylated homodimeric protein with a subunit molecular mass of 12 kDa. In addition to the conventional protein purification techniques, lectin has also been purified using reverse phase high-performance liquid chromatography and evaluated for biological properties such as antiviral and cytotoxic potential. Lectin did not show inhibition against five human cancer cell lines representing different organs and tissues. Interestingly, it was found to be nontoxic towards BSC-1 cells (African green monkey kidney epithelial cells) and exhibited a significant antiviral potential against a strain of vaccinia virus in vitro.
PDF Abstract XML References Citation
How to cite this article
Amandeep Kaur, Rajinder Singh, Sukhdev Singh Kamboj, Jatinder Singh and Girish J. Kotwal, 2008. In vitro Antiviral Activity of Crinum latifolium
Lectin Against Poxvirus Replication. Journal of Biological Sciences, 8: 1236-1240.
DOI: 10.3923/jbs.2008.1236.1240
URL: https://scialert.net/abstract/?doi=jbs.2008.1236.1240
DOI: 10.3923/jbs.2008.1236.1240
URL: https://scialert.net/abstract/?doi=jbs.2008.1236.1240
INTRODUCTION
Plant lectins are a heterogeneous group of proteins of non-immune origin that reversibly bind to mono- and/or oligosaccharides with high specificity, but are devoid of catalytic activity (Rudiger and Gabius, 2001). Their unique ability to recognize and reversibly bind to specific carbohydrate ligands distinguishes them from other carbohydrate-binding proteins and enzymes and makes them invaluable tools in biomedical and glycoconjugate research. Out of seven groups of plant lectins based on their structural and evolutionary relatedness (Van Damme et al., 1998), monocot mannose-binding lectins having exclusive specificity towards poly- and/or oligo-mannose residues, have interesting biological properties. These lectins represent a well-defined class of antiretroviral compounds and exhibit this activity through their binding to mannosylated region of envelope glycoproteins of various reteroviruses such as human immunodeficiency virus, simian immunodeficiency virus, feline immunodeficiency virus (Davidson et al., 2000; Balzarini et al., 2004). Besides, some of these are also potent in vitro inhibitors of cytomegalovirus (Balzarini et al., 1991), human coronavirus (SARS) and feline coronavirus (Keyaerts et al., 2007), herpes simplex (type 1) and influenza (H1N1) type A (Linda et al., 2004) and vaccinnia virus in vitro (Kaur et al., 2007). Hippeastrum hybrid lectin from the family Amaryllidaceae, has been reported as potential microbicide against HIV (Saïdi et al., 2007). Moreover, a few monocot mannose-binding lectins have also been reported to inhibit cell proliferation by interacting with specific carbohydrates on tumor cell surfaces (Wang et al., 2000). Recently, a monocot mannose binding lectin Tryphonium divaricatum, has been reported to exhibit in vitro antiviral activity against HSV-2 and anti-proliferative effect on human cancer cell lines (Luo et al., 2007).
In continuation to the earlier study on isolation and characterization of new monocot lectins (Kaur et al., 2005a, b), present report describes the antiviral activity of Crinum latifolium lectin against a strain of vaccinia virus in vitro.
MATERIALS AND METHODS
Materials: Bulbs of Crinum latifolium were collected from the foothills of Jammu, India, during the months of September-October. The identity of the plant used was confirmed at the herbarium of the Regional Research Laboratory at Jammu, India.
Lectin isolation and purification: Lectin was isolated from the bulbs of the C. latifolium and purified on DEAE-sephacyl anion exchange column followed by gel filtration chromatography using Biogel P-200, as described in our earlier communication (Kaur et al., 2006).
High performance liquid chromatography: Size-exclusion chromatography of ion-exchange and gel-filtration purified lectin was performed using Synchropak GPC-300 column with Advanced Protein Purification System (Waters) attached to UV-monitor and recorder. The column (250x4.60 mm) was washed with double distilled water and the flow rate was set at 0.8 mL min-1. It was equilibrated with 50 mM PBS, pH 7.2 till the baseline was stable. The PBS dialyzed purified lectin was injected into the column and eluted with 50 mM PBS, pH 7.2. Reversed-phase high-performance liquid chromatography was performed using Jupiter C-18 column (Kaur et al., 2007). The column was washed with distilled water, 70% methanol, equilibrated with 0.1% trifluoroacetic acid and water at a flow rate of 1 mL min-1. After loading, the lectin was eluted with a linear gradient (v/v) of buffer A (0.1% trifluoroacetic acid in water) and buffer B (0.1% trifluoroacetic acid in 70% acetonitrile) at a flow rate of 1 mL min-1.
Cytotoxicity assay: In vitro cytotoxic potential of C. latifolium against human cancer cell lines A-549 (breast), HT-29 (colon), SiHa (cervix), SNB-78 (CNS) and PC-3 (prostate) was assessed by the method as in Skehan et al. (1990). The wells in the microtitre plate were seeded in triplicate with 105 cells/well in 100 μL of medium from different cancer cell lines maintained in RPMI-1640 medium, supplemented with 10% FCS followed by incubation for 24-28 h in CO2 incubator. Subsequently, 100 μL of C. latifolium lectin was added at concentrations ranging from 1-100 μg mL and incubated for additional 48 h. The adherent cell cultures were fixed in situ by adding 50 μL of 50% (w/v) trichloroacetic acid (final concentration, 10% TCA) and incubated for 1 h at 4°C. The supernatant was discarded and the plates were washed five times with deionized water and dried. One hundred microlitres of sulphorhodamine B (SRB, 0.4% w/v in 1% acetic acid) were added to each well and incubated for 10 min at room temperature. The unbound SRB was removed by washing five times with 1% acetic acid and plates dried. The bound stain was solubilized with 50 μL Tris-HCl buffer (10 mM, pH 10.5) and absorption was measured at 540 nm using Multiscan EX (Labsystems) ELISA reader to determine relative cell growth or viability in treated and untreated cells. The anticancer drugs 5-fluorouracil, mytomycin-C and paclitaxel were used as standards. All steps were carried out under sterile conditions.
Toxicity and antiviral assays: The inhibitory potential of C. latifolium lectin against poxviral replication was tested on an attenuated strain of vaccinia virus vGK5 (Kotwal et al., 1989). BSC-1 cell line (African green monkey kidney epithelial cells, ATCC No. CCL26) monolayers were grown to 100% confluence in wells of a 96-well culture plate (Costar, USA) in Earle`s base minimal essential medium containing L-glutamine, non-essential amino acids and NaHCO3 (Highveld Biological, Cape Town, South Africa) and Fetal Calf Serum (Highveld Biological) at a final concentration of 10%. Wells were infected with serial dilutions of untreated virus or virus (100 μL aliquots) treated with 5, 10 and 20 μL of C. latifolium with a lectin content of 8.6, 17.2 and 34.4 μg for 15 min at 37°C. Concurrently, wells were directly inoculated with 5, 10 or 20 μL of lectin, with the same net lectin content, to determine toxicity to the cell monolayers. The cultures were incubated at 37°C in a CO2 incubator (5% CO2) for 48 h. During this time period, infectious virus particles destroy cells in culture producing circular breaks in monolayer known as plaques each representing an infectious virus particle. Following incubation, the media from each well was aspirated off and cell monolayers stained with a Crystal Violet (Merck, Germany) solution (10 Crystal Violet, 20 ethanol, 70 dH2O) to allow visualization of viral plaques. Plaques were counted in each well and the antipoxviral activity of the C. latifolium lectin quantified by calculating the percentage reduction in plaque number following treatment. Toxicity of the lectin was assessed by estimating the percentage of intact cell monolayer in each well.
RESULTS AND DISCUSSION
A new monocot mannose-binding lectin from the bulbs of C. latifolium, specific for methyl-α-D-mannopyranoside has earlier been purified using ion-exchange followed by gel filtration chromatography. The purified lectin was 5.546% (w/w) of total extractable protein and 0.033% (w/w) of total bulb weight. The lectin showed a molecular mass of 24 kDa on gel filtration. Absence of disulphide linkages and the homodimeric nature of the purified lectin were revealed by SDS-PAGE. It did not require metal ions for lectin activity and was non-mitogenic and non-glycosylated protein (Kaur et al., 2006) similar to monocot mannose binding lectin Galanthus nivalis (GNA) from the family Amaryllidaceae, which has been exploited as an affinity reagent for the purification of glycoproteins (Shibuya et al., 1988; Rousseau et al., 1997).
| Fig. 1: | (a) Size-exclusion chromatography of ion exchange and gel-filtration purified C. latifolium lectin was performed using Synchropak GPC-300 column using Advanced Protein Purification System (Waters). The elution was performed with 50 mM PBS, pH 7.2. (b) Reversed-phase high performance liquid chromatography of ion exchange and gel-filtration purified C. latifolium lectin was performed using Jupiter C-18 column. Lectin was centrifuged and injected into the column and the elution was performed with a linear gradient (v/v, secondary Y-axis) of buffer A (0.1% trifluoroacetic acid water) and buffer B (0.1% trifluoroacetic acid in 70% acetonitrile) at a flow rate of 1 mL min-1 |
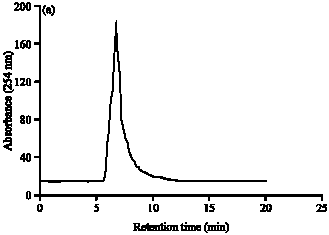
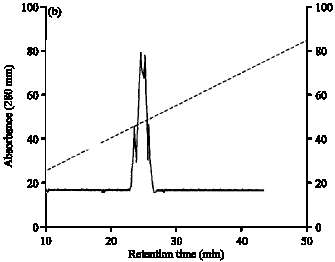
The purified lectin, when subjected to size exclusion and reverse phase chromatography using high performance liquid chromatography yielded a single peak in HP-SEC showing homogenous nature (Fig. 1a). However, in reverse phase chromatography, it gave multiple peaks (Fig. 1b), thereby supporting our earlier results of isoelectric focusing (pI range 5.09-6.18) and alkaline-PAGE at pH 8.3, indicating multiple isoforms (Kaur et al., 2006) similar to other lectins from Amaryllidaceae (Van Damme et al., 1991; Ooi et al., 2000).
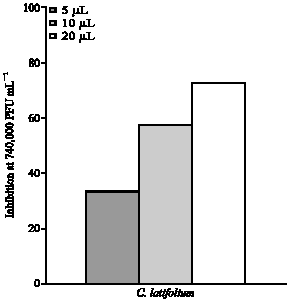 | |
| Fig. 2: | Inhibitory potential of purified C. latifolium lectin against poxviral replication. Percent inhibition is the mean value of experiments conducted three times. The volume 5, 10 and 20 μL-1 with a respective lectin content of 8.6, 17.2 and 34.4 μg was used to evaluate toxicity and antipoxviral potential |
In the light of reports on the antiviral potential of some monocot MBLs from Amaryllidaceae (Balzarini et al., 1991, 2004) and immunomodulatory, cytotoxic and antiviral activities displayed by the aqueous extracts of C. latifloium (Zvetkova et al., 2001); cytotoxic and antiviral potentials of the C. latifloium lectin have been evaluated.
In vitro cytotoxic effect of the C. latifloium lectin was assessed against five human cancer cell lines, which were not inhibited at all the concentrations used (results not shown). Before evaluating its inhibitory potential towards poxvirus replication, it was tested for its toxicity towards BSC-1 cells (African green monkey kidney epithelial cells) and was found to be non-toxic at all the concentrations used. Interestingly, it was a potent inhibitor of poxvirus replication in vitro with a maximum inhibition of 73% at a concentration of 34.4 μg (Fig. 2). At concentrations, 17.2 and 8.6 μg, it gave 63 and 33% inhibition, respectively. The most likely general mechanism of the antiviral activity of carbohydrate-binding proteins has recently been proposed as the interruption of virus fusion with its target cell and may be mediated either by direct binding to the glycans present on the virus envelope or by cross-linking glycans during virus/cell interaction thereby preventing further interaction with the coreceptor (Balzarini, 2006). Carbohydrate-binding molecules such as retrocyclin 2 (a multivalent lectin) inhibit viral fusion and entry by crosslinking membrane glycoproteins. Certain endogenous lectins (defensins and collectins) also crosslink and/or immobilize glycoproteins on viral cellular surfaces, thereby forming a network of immobilized glycoproteins that blocks viral fusion and entry (Leikina et al., 2005). The antiviral activity of C. latifolium lectin may also be attributed to similar interactions with glycoproteins on the viral surface.
It may be noted that potent antiviral agents, asscytovirin and cyanovirin-N (Bolmstedt et al., 2001; Barrientos et al., 2003; O`Keefe et al., 2003) are also mannose-binding proteins other than plant lectins. In addition, the similarity of C. latifolium with other antiviral lectins from Amaryllidaceae including agglutinating rabbit but not murine or human erythrocytes, non-mitogenicity, pH and thermal stability and non-toxic nature; raises the possibility to use the lectin as a new potential candidate against multiple viral diseases caused by enveloped viruses and needs to be further investigated with respect to the mechanistic details at molecular level to explore the basis of biological activity.
REFERENCES
- Balzarini, J., D. Schols, J. Neyts, E.J.M. Van Damme and W.J. Peumans et al., 1991. α-(1-3)- and α-(1-6)-D-mannose-specific plant lectins are markedly inhibitory to human immunodeficiency virus and cytomegalovirus infections in vitro. Antimicrob. Agents Chemother., 35: 410-416.
PubMed - Balzarini, J., S. Hatse, K. Vermeire, K. Princen and S. Aquaro et al., 2004. Mannose-specific plant lectins from the Amaryllidaceae family qualify as microbicides for prevention of human immunodeficiency virus infection. Antimicrob. Agents Chemother., 48: 3858-3870.
CrossRef - Balzarini, J., 2006. Inhibition of hIV entry by carbohydrate-binding proteins. Antiviral Res., 71: 237-247.
CrossRef - Barrientos, L.G., B.R. O'Keefe, M. Bray, A. Sanchez and A.M. Gronenborn et al., 2003. Cyanovirin-N binds to the viral surface glycoproteins, gP1 2 and inhibits infectivity of Ebola virus. Antiviral Res., 58: 47-56.
CrossRef - Bolmstedt, A.J., B.R. O'Keefe, S.R. Shenoy, J.B. McMohan and M.R. Boyd, 2001. HIV-inhibitory natural products part 71-cyanovirin-N defines a new class of antiviral agent targeting N-linked, high mannose glycans in an oligosaccharide-specific manner. Mol. Pharmcol., 59: 949-954.
Direct Link - Davidson, E., J.M.S. Forrest, I.M. Morrison and D. Stewart, 2000. Mannose-Specific Plant Lectins From Plants as Diagnostics, Vaccines and Tools for the Elucidation of Viral Infection Mechanisms in Animals. In: Annual Report for the Scottish Crop Research Institute for 1999/2000, Smith, W.H.M. and T.D. Heilbronn (Eds.). SCRI, Dundee, Scotland, pp: 125-128.
- Kaur, A., J. Singh, S.S. Kamboj, A.K. Saxena and R.M. Pandita et al., 2005. Isolation of an N-acetyl-D-glucosamine specific lectin from the rhizomes of arundo donax with antiproliferative activity. Phytochem, 66: 1933-1940.
CrossRef - Kaur, A., S.S. Kamboj, J. Singh, A.K. Saxena and V. Dhuna, 2005. Isolation of a novel N-acetyl-D-lactosamine specific lectin from alocacia cucullata (Schott.). Biotech. Lett., 27: 1815-1820.
CrossRef - Kaur, A., S.S. Kamboj and J. Singh, 2006. Isolation of a new lectin from the bulbs of Crinum latifolium (L.). J. Biological Sci., 6: 9-14.
CrossRefDirect Link - Kaur, A., S.S. Kamboj, J. Singh, R. Singh and M. Abrahams et al., 2007. Purification of 3 monomeric monocot mannose-binding lectins and their evaluation for antipoxviral activity: Potential applications in multiple viral diseases caused by enveloped viruses. Biochem. Cell. Biol., 85: 88-95.
CrossRef - Keyaerts, E., L. Vijgen, C. Pannecouque, E. Van Damme and W. Peumans et al., 2007. Plant lectins are potent inhibitors of coronavirus by interfering with two targets in the viral replication cycle. Antiviral Res., 75: 179-187.
CrossRef - Kotwal, G.J., A.W. Hugin and B. Moss, 1989. Mapping and insertional mutagenesis of a vaccinia virus gene encoding a 13,800-Da secreted protein. Virology, 171: 579-587.
CrossRefDirect Link - Leikina, E., H. Delanoe-Ayari, K. Melikov, M.S. Cho and A. Chen et al., 2005. Carbohydrate-binding molecules inhibit viral fusion and entry by crosslinking membrane glycoproteins. Nature Immunol., 6: 995-1001.
CrossRef - Luo, Y., X. Xu, J. Liu, J. Li and Y. Sun et al., 2007. A novel mannose-binding tuber lectin from typhonium divaricatum (L.) decne (family araceae) with antiviral activity against HSV-II and anti-proliferative effect on human cancer cell lines. J. Biochem. Mol. Biol., 40: 358-367.
PubMed - O'Keefe, B.R., D.F. Smee, J.A. Turpin, C.J. Saucedo and K.R. Gustafson et al., 2003. Potent anti-influenza activity of cyanovirin-N and interactions with viral hemagglutinin. Antimicrob. Agents Chemother., 47: 2518-2525.
CrossRef - Ooi, L.S., T.B. Ng, Y. Geng and V.E. Ooi, 2000. Lectins from bulbs of the Chinese daffodil Narcissus tazetta (Family amaryllidaceae). Biochem. Cell. Biol., 78: 463-468.
CrossRef - Rousseau, C., M. Felin, M.A. Doyennette-Moyne and A.P. Sève, 1997. CB70, a glycosylated nuclear lectin. J. Cell. Biochem., 66: 370-385.
CrossRef - Rudiger, H. and H.J. Gabius, 2001. Plant lectins: Occurrence, biochemistry, functions and applications. Glycoconjugate J., 18: 589-613.
CrossRefPubMedDirect Link - Saïdi, H., N. Nasreddine, M.A. Jenabian, M. Lecerf and D. Schols et al., 2007. Differential in vitro inhibitory activity against HIV-1 of alpha-(1-3)- and alpha-(1-6)-D-mannose specific plant lectins: Implication for microbicide development. J. Trans Med., 5: 28-28.
CrossRef - Shibuya, N., J.E. Berry and I.J. Goldstein, 1988. One-step purification of murine iGM and human α2-macroglobulin by affinity chromatography on immobilized snowdrop bulb lectin. Arch. Biochem. Biophys., 267: 676-680.
CrossRef - Skehan, P., R. Storeng, D. Scudiero, A. Monks and J. McMahon et al., 1990. New colorimetric cytotoxicity assay for anticancer-drug screening. J. Nat. Cancer Inst., 82: 1107-1112.
CrossRefPubMedDirect Link - Van Damme, E.J.M., I.J. Goldstein and W.J. Peumans, 1991. A comparative study of mannose-binding lectins from Amaryllidaceae and Alliaceae. Phytochemistry, 30: 509-514.
CrossRef - Wang, H., T.B. Ng, V.E.C. Ooi and W.K. Liu, 2000. Effects of lectins with different carbohydrate-binding specificities on hepatoma, choriocarcinoma, melanoma and osteosarcoma cell lines. Int. J. Biochem. Cell. Biol., 32: 365-372.
CrossRef - Zvetkova, E., B. Wirleitner, N.T. Tram, H. Schennach and D. Fuchs, 2001. Aqueous extracts of Crinum latifolium (L.) and Camellia sinensis show immunomodulatory properties in human periphereal blood mononuclear cells. Int. Immunopharmacol., 1: 2143-2150.
CrossRef - Kaur, A., S.S. Kamboj and J. Singh, 2006. Isolation of a new lectin from the bulbs of Crinum latifolium (L.). J. Biological Sci., 6: 9-14.
CrossRefDirect Link - Ooi, L.S.M., S.S.M. Sun and V.E.C. Ooi, 2004. Purification and characterization of a new antiviral protein from the leaves of Pandanus amaryllifolius (Pandanaceae). Int. J. Biochem. Cell. Biol., 36: 1440-1446.
CrossRef