
M. Ahmadi
Department of Animal Science, Islamic Azad University, Ilam, Iran
Y. Mohammadi
Department of Animal Science, University of Ilam, Iran
H. Darmani Kuhi
Department of Animal Science, University of Ilam, Iran
R. Osfoori
Agricultural Biotechnology Research Institute of Iran (ABRII), Iran
S. Qanbari
Research Institute of Physiology and Biotechnology, University of Zanjan, P.O. Box 313, Iran
Journal of Biological Sciences
Year: 2008 | Volume: 8 | Issue: 7 | Page No.: 1231-1235







ABSTRACT
The polymorphism in the bovine κ-casein (κ-CN) and β-lactoglobulin (β-LG) genes were analyzed for DNA sequence variants using the polymerase chain reaction-restriction fragment length polymorphism (PCR-RFLP) technique in Iranian Holstein population to find association between the genotypes and milk traits as well as somatic cell score. To this purpose, 2537 test day records of one hundred thirty nine dairy cows were entered in a repeated generalized mixed model. The frequencies of alleles A and B were estimated as 0.81, 0.19 and 0.57, 043 for κ-CN and β-LG loci, respectively. The results of association analysis by the mixed model revealed a significant positive relationship of κ-CN genotype AA on milk yield (p≤0.04). These analyses did not reveal a significant association of κ-CN genotypes with fat percent and SCS traits. In the case of β-LG locus statistical analysis showed a strong relationship between BB genotype and protein percentage (p≤0.007) comparing to the other genotypes. These results did not reveal a significant association of β-LG genotypes with milk yield, milk fat percentage and SCS in studied animals. A tendency to being significant was however observed for β-LG genotype AA (p<0.12) with the SCS trait. Because of the lack of consistency among results of similar studies, we suggest further investigations to determine the precise nature of these associations with the milk proteins to be performed based on haplotypes.
PDF Abstract XML References Citation
How to cite this article
M. Ahmadi, Y. Mohammadi, H. Darmani Kuhi, R. Osfoori and S. Qanbari, 2008. Association of Milk Protein Genotypes with Production Traits and Somatic Cell Count of Holstein Cows. Journal of Biological Sciences, 8: 1231-1235.
DOI: 10.3923/jbs.2008.1231.1235
URL: https://scialert.net/abstract/?doi=jbs.2008.1231.1235
DOI: 10.3923/jbs.2008.1231.1235
URL: https://scialert.net/abstract/?doi=jbs.2008.1231.1235
INTRODUCTION
Genetic markers highly related to production characteristics could be used to predict future animal performance and could thus serve as additional selection criteria. During the last decades genetic polymorphism of milk proteins has been of great interest in animal breeding and the dairy industry due to its relationships with production traits, milk composition and quality (Ng-Kwai-Hang et al., 1984; Lin et al., 1986; Ng-Kwai-Hang et al., 1990; Tsiaras et al., 2005). κ-casein (κ-CN) and β-lactoglobulin (β-LG) are the most important proteins expressed in milk. Due to their potential use as an aid to genetic selection, several tens of studies have been carried out to identify genetic variation and frequencies of their variants in different cattle breeds (Mitra et al., 1998; Soria et al., 2003; Rachagani et al., 2006; Sulimova et al., 2007) and possible associations with economically important traits (Arave et al., 1971; McLean et al., 1984; Ng-Kwai-Hang et al., 1990b; Lundén et al., 1997). In contrast to the numerous studies trying to find the effects of milk protein genetic polymorphism on milk production traits, only a few reports were published regarding their association with Somatic Cell Count (SCC) in studied animals (Lundén et al., 1997). Somatic Cell Count (SCC) in milk constitutes a good diagnostic tool that allows early detection of either subclinical or acute form of mastitis (Green et al., 2004) and is therefore a valuable component of monitoring programs (Schukken et al., 2003). The objectives of this study were to determine the frequencies of κ-CN and β-LG variants and to analyze the relationship between genotypes of these genes on milk production and Somatic Cell Count (SCC) as an indicator of udder health in Iranian Holstein population.
MATERIALS AND METHODS
Data collection: Blood samples were collected by sterile tubes containing EDTA anticoagulant from one hundred thirty nine dairy cows belonging to 6 different dairy farms participating in the recording system of National Animal Breeding Center. For statistical analyses, 2537 test records of milk yield, protein percent, fat percent and somatic cell count traits taken from 2001-2006 were used. These records were taken directly from the national breeding value evaluation for the Holstein population. The DNA was extracted from blood cells by a modified phenol chloroform method. The number of test day records for each animal varied between 8 and 27.
PCR-RFLP assay for β-LG genotypes: The PCR reactions were carried out in a thermocycler (i-Cycler, Biorad). According to Medrano and Aguilar-Cordova (1990a), forward and reverse primers: 5` TGT GCT GGA CAC CGA CTA CAA AAA 3` and 5` GCT CCC GGT ATA TGA CCA CCC TCT 3` were used to amplify a 247 bp DNA fragment by PCR. Each 15 μL PCR reaction contained 50 ng template DNA, 200 μM each of dNTP, 2.5 mM MgCl2. 2.5 pmol each primer, 1X PCR standard reaction buffer and 1.5 unit of Biotherm Taq polymerase. The PCR reaction included the following steps: predenaturation for 3 min at 94°C followed by 30 cycles of 94°C for 1.5 min, 58°C for 1 min, 72°C for 2 min and a final extension for 10 min at 72°C. The amplified PCR product was digested using Hae 3 and 1x reaction buffer at 37°C overnight. The digested product was loaded and visualised on 3% agarose gel after staining with ethidium bromide.
PCR-RFLP assay for κ-CN genotypes: The kappa-casein CSN3 genotypes were determined as described by Medrano and Aguilar-Cordova (1990b) using the polymerase chain reaction-restriction fragment length polymorphism (PCR-RFLP) technique. As a detail, a 350 bp DNA fragment was amplified by adding forward primer (5` ATC ATT TAT GGC CAT TCC ACC AAA G 3`) and reverse primer (5`GGC CAT TTC GCC TTC TCT GTA ACA GA 3`). Each PCR reaction contained 50 ng template DNA, 200 μM each of dNTP, 2.5 pmol each primer, 2.5 mM MgCl2, 1X PCR standard reaction buffer, 1 unit of Biotherm Taq polymerase and sterilised distilled water to make a final volume of 15 μL. The PCR reaction included the following steps: predenaturation for 3 min at 94°C followed by 30 cycles 94°C for 30 sec, 58°C for 1 min 72°C for 2 min and a final extension of 10 min at 72°C. The amplified PCR product was digested using HinfI and 1x reaction buffer at 37°C overnight. The digested product was loaded and visualised on 4% agarose gel after staining with ethidium bromide.
Statistical analysis: A repeated generalized MIXED model (SAS Inst., Inc., Cary, NC) was used to evaluate the fixed effects of herd, parity, season and year of the recorded data, genotypes and random permanent effects of each animal on production traits and SCS in the Holstein data set. For the present study, the milk somatic cell count data was transformed to somatic cell score (SCS = log base 2 (SCC/100,000)+3) and was entered to the statistical model. The mixed repeatability model was:
| Yijklmn = μ+Pi+Sj+Hk+Tl+Gm+Rijklm+eijklmn |
where, Yijklmn is the dependent variable of production traits (Milk yield, fat, or protein percentages) and SCC, μ is the population mean of the analyzed traits, Pi is the fixed effect of the parity i (5 subclasses), Sj is the fixed effect of season j (4 subclasses), Hk is the fixed effect of the herd k (6 subclasses), Tl is the fixed effect of year l (5 subclasses), Gm is the fixed effect of genotype m (3 subclasses for each locus), Rijklmn is the random permanent animal effect with mean 0 and variance s2δ, which is equal to the covariance between repeated measurements within animals and finally eijklmn is the random residual effect with mean 0 and variance σ2. The compound symmetric covariance matrix structure was applied for repeated observations of subclinical mastitis which was fitted based on the Akaike information criterion (Akaike, 1974).
RESULTS AND DISCUSSION
Data statistics and allele frequencies: The means and standard deviations for the milk yield, fat and protein percentages were calculated as 28.64 (9.93), 3.41 (0.68) and 3.13 (0.31), respectively. These values are typical for Holstein cows in the studied population and allow a comparison of the size of the κ-CN and β-LG genotypes effects. Totally, 25% of somatic cell count records were upper than 300,000 cells mL-1. The geometric population mean of the number of somatic cell count, called also as bulk milk SCC value as an indicator of udder health status in studied population was calculated as 268,000 cells mL-1. In the European countries equivalent SCC values in 2002 were 200,000, 132,000 and 221,000 cells mL-1 for Netherlands, Finland and Belgium, respectively (Piepers et al., 2007). For Uruguay this value was reported to be around 450 000-500 000 cells mL-1, indicating about 50% prevalence of sub-clinical mastitis in the studied population (Gianneechini et al., 2002).
Table 1 summarizes the distribution of frequencies of κ-CN and β-LG alleles and genotypes in studied population. The frequencies of alleles A and B were estimated as 0.81, 0.19 and 0.57, 043 for κ-CN and β-LG loci, respectively.
| Table 1: | The frequencies of κ-CN and β-LG alleles and genotypes |
| *Expected heterozygosity | |
The allele frequencies reported in the studies of Lundén et al. (1997) and Bobe et al. (1999) for κ-CN in Sweden and USA as well as Tsiaras et al. (2005) and Hill et al. (1993) for β-LG in Greece and New Zealand, respectively, confirm the frequencies obtained in current study. Undoubtfully, the distribution of Holstein semen worldwide and using of imported semen extensively in the country farms has resulted in the complete accordance with the outcomes of the aforementioned investigations. Possible divergence from Hardy-Weinberg expectations at each locus was evaluated. Both loci showed significant deviation (p<0.05) from Hardy-Weinberg proportions. These results were expected, because a number of basic underlying assumptions of Hardy-Weinberg equilibrium such as random mating, infinite population size and equal parental contribution in a breeding population were violated.
Association analysis: Table 2 summarizes the association results of the effects included in the statistical model. The effects of year, season, herd and parity on production traits were significant (p<0.05). Also the effects of parity and herd on SCS were significant (p<0.05).
The results of association analysis by the mixed repeatability model revealed a significant positive relationship of κ-CN genotype AA on milk yield (p≤0.04). The AA genotype of κ-CN was also negatively associated with protein percentage (p≤0.07); as such the genotypes AB and BB had the higher protein percentage. These analyses did not reveal a significant association of κ-CN genotypes with fat percent and SCS traits. In the case of β-LG locus, statistical analysis showed a strong relationship between BB genotype and elevated protein percentage (p≤0.007) comparing to the other genotypes. These results did not show a significant association of β-LG genotypes with milk yield, milk fat percent and SCS in studied animals. A tendency to being significant was however observed for genotype AA (p<0.12) with the SCS trait (Table 2).
Associations between milk protein variants and production traits have been documented by several authors. As a general result, B κ-CN variant has a favorable effect on protein yield (Ng-Kwai-Hang et al., 1984; Van Eenennaam and Medrano, 1991) and protein content (Gonyon et al., 1987; Ng-Kwai-Hang et al., 1990b; Van Eenennaam and Medrano, 1991; Bovenhuis et al., 1992; Van den Berg et al., 1992). The AA genotype of β-LG has also been shown to have a favorable effect on protein yield (Ng-Kwai-Hang et al., 1984; Aleandri et al., 1990; Bovenhuis et al., 1992), whereas positive effects of the BB genotype on fat content have been reported (McLean et al., 1984; Ng-Kwai-Hang et al., 1984, 1986; Aleandri et al., 1990; Bovenhuis et al., 1992; Hill, 1993).
| Table 2: | The least square means and standard deviation of κ-CN and β-LG genotypes with analyzed traits |
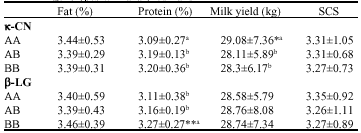 | |
| a,b:Values with different superscripts are significantly different, *p<0.05, **p<0.01 | |
Although, published reports regarding the effects of variants of κ-CN and β-LG on milk production traits apparently, have reached to a consensus result, in the most cases they were conflicting with respect to the significance and the size of genotype effects. For example the B variant of κ-CN has been associated with higher (Ng-Kwai-Hang et al., 1986; Lin et al., 1989; Van Eenennaam and Medrano, 1991) and lower (Bovenhuis et al., 1992) milk yield. whereas other studies have indicated no effect (Arave et al., 1971; McLean et al., 1984; Ng-Kwai-Hang et al., 1984; Lin et al., 1986; Ng-Kwai-Hang et al., 1990b; Lundén et al., 1997). There are also some studies that failed to find any relationships between κ-CN genotypes with production traits (Arave et al., 1971; McLean et al., 1984; Ng-Kwai-Hang et al., 1984; Lin et al., 1986; Ng-Kwai-Hang et al., 1990; Lundén et al., 1997).
In the case of β-LG also, several authors have reported no significant associations of genotypes on milk yield (McLean et al., 1984; Ng-Kwai-Hang et al., 1984; Lin et al., 1986, 1989; Van Eenennaam and Medrano, 1991; Lundén et al., 1997; Ojala et al., 1997). However, reports exist where β-LG genotype AA (Aleandri et al., 1990; Bovenhuis et al., 1992), AB (Pupkova, 1980), or BB (Jairam and Nair, 1983) have been positively associated with milk yield.
Contrary to the production traits, there are only a few studies in literature trying to find an association between the polymorphism of κ-CN and β-LG with milk somatic cell count records. As one of the rare studies on this subject, Lunde΄n et al. (1997) reported no association between the polymorphism of κ-CN and β-LG with milk SCC. This observation is in coincidence with the result of the current study.
Like with the other reports, the results presented in this study are in coincidence with and in some cases differ from the results of previous studies. The observed inconsistency could be explained by different reasons. The reason might be that the associations are due to the effects of linked genes rather than to the effects of the milk protein loci themselves (Bovenhuis et al., 1992). In this case, the associations in different populations might differ. The conflicting observations may also be a consequence of different management and feeding regimens, population size and breed, as well as availability and accuracy of data in the various studies. The kind of data used in different studies (breeding values, daughters yield deviations or daily production records) or the statistical model used to analyze could be also other reasons.
In conclusion, it is suggested, because of the lack of consistency among results of similar studies, further investigations to determine the precise nature of these associations with the milk proteins to be performed based on haplotypes.
ACKNOWLEDGMENTS
This study was financially supported by Ilam Azad University (grant No. 52015840501002). The authors wish to thank to National Animal Breeding Centre, for providing the recorded data.
REFERENCES
- Aleandri, R., L.G. Buttazzoni, J.C. Schneider, A. Caroli and R. Davoli, 1990. The effects of milk protein polymorphisms on milk components and cheese-producing ability. J. Dairy Sci., 73: 241-255.
CrossRefDirect Link - Akaike, H., 1974. A new look at the statistical model identification. IEEE Trans. Autom. Control, 19: 716-723.
CrossRefDirect Link - Arave, C.W., R.C. Lamb and H.C. Hines, 1971. Blood and milk protein polymorphisms in relation to feed efficiency and production traits of dairy cattle. J. Dairy Sci., 54: 106-112.
CrossRefPubMedDirect Link - Bobe, G., D.C. Beitz, A.E. Freeman and G.L. Lindberg, 1999. Effect of milk protein genotypes on milk protein composition and its genetic parameter estimates. J. Dairy Sci., 82: 2797-2804.
CrossRefPubMedDirect Link - Bovenhuis, H., J.A.M. van Arendonk and S. Korver, 1992. Associations between milk protein polymorphisms and milk production traits. J. Dairy Sci., 75: 2549-2559.
CrossRefPubMedDirect Link - Gianneechini, R., C. Concha, R. Rivero, I. Delucci and J.M. Lopez, 2002. Occurrence of clinical and sub-clinical mastitis in dairy herds in the west littoral region in Uruguay. Acta Vet. Scand., 43: 221-230.
Direct Link - Gonyon, D.S., R.E. Mather, H.C. Hines, G.F.W. Haenlein, C.W. Arave and S.N. Gaunt, 1987. Associations of bovine blood andmilk polymorphisms with lactation traits: Holsteins. J. Dairy Sci., 70: 2585-2598.
Direct Link - Green M.J., L.E. Green, Y.H. Schukken, A.J. Bradley and E.J. Peeler et al., 2004. Somatic cell count distributions during lactation predict clinical mastitis. J. Dairy Sci., 87: 1256-1264.
Direct Link - Jairam, B.T. and P.G. Nair, 1983. Genetic polymorphism of milk proteins and economic characters in dairy animals. Indian J. Anim. Sci., 53: 1-8.
Direct Link - Lin, C.Y., A.J. McAllister, K.F. Ng-Kwai-Hang and J.F. Hayes, 1986. Effects of milk protein loci on first lactation production in dairy cattle. J. Dairy Sci., 69: 704-712.
CrossRefDirect Link - Lin, C.Y., A.J. McAllister, K.F. Ng-Kwai-Hang, J.F. Hayes and T.R. Batra et al., 1989. Relationships of milk protein types to lifetime performance. J. Dairy Sci., 72: 3085-3090.
CrossRefPubMedDirect Link - Lunden, A., M. Nilsson and L. Janson, 1997. Marked effect of β-lactoglobulin polymorphism on the ratio of casein to total protein in milk. J. Dairy Sci., 80: 2996-3005.
CrossRefDirect Link - McLean, D.M., E.R.B. Graham, R.W. Ponzoni and H.A. McKenzie, 1984. Effects of milk protein genetic variants on milk yield and composition. J. Dairy Res., 51: 531-546.
CrossRefPubMedDirect Link - Medrano, J.F. and E. Aquilar-Cordova, 1990. Genotyping of bovine kappa-casein loci following DNA sequence amplification. Biotechnology, 8: 144-146.
PubMed - Medrano, J.F. and E. Aquilar-Cordova, 1990. Polymerase chain reaction amplification of bovine beta-lactoglobulin genomic sequences and identification of genetic variants by RFLP analysis. Anim. Biotechnol., 1: 73-77.
CrossRef - Mitra, A., P. Schlee, I. Krause, J. Blusch, T. Werner, C.R. Balakrishnan and F. Pirchner, 1998. Kappa-casein polymorphisms in Indian dairy cattle and buffalo: A new genetic variant in buffalo. Anim. Biotechnol., 9: 81-87.
PubMed - Ng-Kwai-Hang, K.F., J.F. Hayes, J.E. Moxley and H.G. Monardes, 1984. Association of genetic variants of casein and milk serum proteins with milk, fat and protein production by dairy cattle. J. Dairy Sci., 67: 835-840.
CrossRefDirect Link - Ng-Kwai-Hang, K.F., J.F. Hayes, J.E. Moxley and H.G. Monardes, 1986. Relationships between milk protein polymorphisms and major milk constituents in Holstein-Friesian cows. J. Dairy Sci., 69: 22-26.
CrossRefDirect Link - Ng-Kwai-Hang, K.F., H.G. Monardes and J.F. Hayes, 1990. Association between genetic polymorphism of milk proteins and production traits during three lactations. J. Dairy Sci., 73: 3414-3420.
CrossRefDirect Link - Ojala, M., T.R. Famula and J.F. Medrano, 1997. Effects of milk production genotypes on the variation of milk production traits of Holstein and Jersey cows in California. J. Dairy Sci., 80: 1776-1785.
PubMed - Piepers, S., L. De Meulemeester, A. de Kruif, G. Opsomer, H.W. Barkema and S. de Vliegher, 2007. Prevalence and distribution of mastitis pathogens in subclinically infected dairy cows in Flanders, Belgium. J. Dairy Res., 74: 478-483.
CrossRefDirect Link - Rachagani, S., I.D. Gupta, N. Gupta and S.C. Gupta, 2006. Genotyping of β-lactoglobulin gene by PCR-RFLP in Sahiwal and Tharparkar cattle breeds. BMC Genet.
CrossRefDirect Link - Schukken, Y.H., D.J. Wilson, F. Welcome, L. Garrison-Tikofsky and R.N. Gonzalez, 2003. Monitoring udder health and milk quality using somatic cell counts. Vet. Res., 34: 579-596.
CrossRefDirect Link - Soria, L.A., G.M. Iglesias, M.J. Huguet and S.L. Mirande, 2003. A PCR-RFLP test to detect allelic variants of the bovine kappa-casein gene. Anim. Biotechnol., 14: 1-5.
PubMed - Sulimova, G.E., M.A. Azari, J. Rostamzadeh, M.R.M. Abani and O.E. Lazebnyi, 2007. Allelic polymorphism of kappa-casein gene (CSN3) in Russian cattle breeds and its informative value as a genetic marker. Genetika, 43: 88-95.
PubMed - Tsiaras, A.M., G.G. Bargouli, G. Banos and C.M. Boscos, 2005. Effect of kappa-casein and beta-lactoglobulin loci on milk production traits and reproductive performance of holstein cows. J. Dairy Sci., 88: 327-334.
CrossRefDirect Link - Van Eenennaam, A. and J.F. Medrano, 1991. Milk protein polymorphisms in California dairy cattle. J. Dairy Sci., 74: 1730-1742.
CrossRefDirect Link