
A.N.M.A. Rahman
Animal Biotechnology-Embryo Laboratory, Institute of Biological Sciences, University of Malaya, Lembah Pantai, 50603 Kuala Lumpur, Malaysia
R.B. Abdullah
Animal Biotechnology-Embryo Laboratory, Institute of Biological Sciences, University of Malaya, Lembah Pantai, 50603 Kuala Lumpur, Malaysia
W.E. Wan-Khadijah
Animal Biotechnology-Embryo Laboratory, Institute of Biological Sciences, University of Malaya, Lembah Pantai, 50603 Kuala Lumpur, Malaysia
Journal of Biological Sciences
Year: 2008 | Volume: 8 | Issue: 7 | Page No.: 1129-1137







ABSTRACT
The study was intended to review the recent developments and advances of estrus synchronization and superovulation protocols in goats with a view to improve oocytes/embryo recovery for In Vitro Production (IVP) efficiencies. Although a number of estrus synchronizing protocols has been developed in goat, the most widely used one is the treatment of progesterone for 9-11 days followed by a luteolytic dose of prostaglandin administered in the period 48h prior to removal of intravaginal sponge. Until now, Controlled Internal Drug Release (CIDR) device and subcutaneous implants are more preferable than sponges. Ovulation in goat can be synchronized more precisely by administering Gonadotrophin Releasing Hormone (GnRH) around the time of estrus that improves the success of fixed-time Artificial Insemination (AI) and oocytes or embryos collection at a controlled stage. Superovulation is the hormonal treatment for increasing a large number of oocytes that ultimately accelerate genetic improvement in any species. Generally an exogenous follicle-stimulating gonadotrophin is administered that mimics the effect of Follicle Stimulating Hormone (FSH) near the end of the luteal phase of the cycle (9-11 days) or around 48 h before the end of the synchronizing treatments. The major commercial products applied are equine Chorionic Gonadotrophin (eCG) or Pregnant Mare Serum Gonadotrophin (PMSG) and FSH. This review paper describes estrus synchronization, ovarian superovulation as well as the normal physiology of estrous cycle and ovulation in goats.
PDF Abstract XML References Citation
How to cite this article
A.N.M.A. Rahman, R.B. Abdullah and W.E. Wan-Khadijah, 2008. Estrus Synchronization and Superovulation in Goats: A Review. Journal of Biological Sciences, 8: 1129-1137.
DOI: 10.3923/jbs.2008.1129.1137
URL: https://scialert.net/abstract/?doi=jbs.2008.1129.1137
DOI: 10.3923/jbs.2008.1129.1137
URL: https://scialert.net/abstract/?doi=jbs.2008.1129.1137
INTRODUCTION
In this modern era Assisted Reproductive Technologies (ARTs) plays a major role in animal reproduction and production. The estrus synchronization and ovarian superovulation already become one of the popular ARTs in goat industry. Estrus synchronization plays a major role in fixed time breeding, Artificial Insemination (AI), Multiple Ovulation-Embryo Transfer (MOET), Laparoscopic Ovum Pick-up (LOPU) for oocyte or embryo collection and Embryo Transfer (ET). The value of estrus synchronization is vital in goats as the duration of both estrous cycle and estrus is variable and estrus detection cannot be accomplished safely without a buck (Jainudeen et al., 2000; Rahman et al., 2008). On the other hand, superovulation is the hormonal treatment for harvesting increased number of oocytes from the ovary than normal. This will ultimately accelerate genetic improvement in goat like any other livestock. It is a means to induce maturation, ovulation and increase the number of follicles available for oocyte recovery.
A complete and scientifically detailed understanding of any physiological process is vital for its successful manipulation. The basic premises of any estrus synchronization and superovulation protocol pivots around the predictable control of events associated with the animal`s reproductive physiology. Control of reproductive events can be achieved through pharmacological administration of biologically active agents. Usually the agents used in estrus synchronization and superovulation protocols are either based on or are the hormones that occur in the female goat or doe at various times during her cycle. Therefore, works related to estrus synchronization and superovulation treatments of the doe have been reviewed briefly in this paper following a short description of estrous cycle.
This study was conducted in Animal Biotechnology-Embryo Laboratory (ABEL), Institute of Biological Sciences, University of Malaya, Lembah Pantai, 50603 Kuala Lumpur, Malaysia from 2004 to 2008.
ESTROUS CYCLE OF THE DOE
The signs of estrus; length and duration of different phases of estrous cycle; hormonal changes during estrous cycle; follicular dynamics, waves and dominant follicles and ovulation in the doe are described following:
Signs of estrus: Signs of estrus are important indicators of onset of estrus or heat and, therefore, very important for estrus detection. These signs are mainly two types, primary and secondary signs. The primary signs are the most reliable and well accepted for indication of estrus behavior in most female animals. The best method of estrus detection is the observation of primary signs exhibited by the doe in response to the buck. However, the doe seldom mount as often as the cow, but demonstrate some behavior when they are in heat such as seeking out bucks, waggling of the tail when being exposed to the buck and also bleating, more frequently if the buck is absent. Apart from the primary signs, physical signs, for example, redness and swelling of the vulva and a clear mucous discharge from the vulva also indicative of estrus. Restlessness, frequent urination, isolation from others, general loss of appetite and constant vocalizations are the secondary signs of estrus. However, secondary signs are less reliable as they vary in length and may be confused with the symptoms of any minor illness. Although it is possible to detect estrus signs in the doe as described above, but a doe in heat may not exhibit all the signs at the same time. These signs appear and disappear progressively with the onset and termination of estrus behavior. Therefore, estrus detection is dependant upon the careful observation and close attention of such phenomena, including the primary and physical signs of the doe experiencing estrus. Therefore, a routine check-up of heat twice daily (morning and evening) is indispensable to obtain optimal results. Estrus detection is a valuable and useful tool for natural mating, AI, predicting parturition date as well as prediction of ovulation time for LOPU.
Length and duration of different phases of estrous cycle: Like ewes, does are also seasonally polyestrus as their reproductive cycle responds to changes in day length (Attwood, 2007). Sexual activity is usually greatest during autumn and winter. Estrus activity of doe is greater in the tropics than in temperate climates. The average length of estrous cycle in the doe is 21 days, but can vary from 18-22 days depending on the breed differences, stage of breeding season and environmental stress (Jainudeen et al., 2000). The abnormally short cycles that are observed in the doe early in the breeding season may be associated with premature regression of the corpus luteum. The estrus lasts for 24-48 h in the doe and the duration of estrus can be influenced by breed, age, season and presence of the buck (Jainudeen et al., 2000). Angora does have a shorter duration of estrus (22 h) than the dairy breeds. Estrus is of shorter duration at the end of breeding season and in the first breeding season of young does. The characteristic events of estrous cycle in doe are shown in Table 1. The complete estrous cycle in doe is divided into 4 well marked phases, namely proestrus, estrus, metestrus and diestrus. In prepubertal and aged does anoestrus (stage of sexual quiescence characterized by lack of estrus behavior) normally occurs. This may be due to the complete suppression of ovarian activity or silent ovulatory cycles without behavioral signs of estrus. In all domestic animals including the doe, anoestrus also may occur as a pathological condition (Pineda, 2003). The duration of each event associated with the estrus cycle and pregnancy in the doe is illustrated in Fig. 1.
| Table 1: | Duration of characteristics events of estrous cycle in doe |
 | |
| Source: Modified from Devendra and McLeroy (1982) and Jainudeen et al. (2000) | |
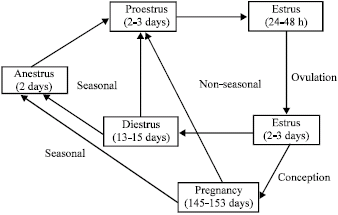 | |
| Fig. 1: | The estrous and reproductive cycles of the female goat. Illustration is based on Pineda (1989) using data cited in Jainudeen et al. (2000), Massita (2003) and Attwood (2007) |
Hormonal changes during estrous cycle in doe: Like other domestic farm animals, improvement in the efficiency of estrus synchronization in the doe depends on a better knowledge of the endocrine patterns that occur during induced and natural estrus (Chemineau et al., 1981). Cyclic changes in the circulating levels of the ovarian hormones, progesterone and estrogen have direct effects on the growth and metabolism of cells in the reproductive tissues. Unlike the ewe, data about the endocrinology of the estrous cycle in the doe is sparse. The main events of the estrous cycle are related to the periods of growth of the ovarian follicles and the corpus luteum. The sequence of hormonal events during the estrous cycle is similar in both doe and ewe, but the doe has a longer progesterone phase than the ewe (Jainudeen et al., 2000). During the 21 days cycle, progesterone dominates for about 15 days and estrogen dominates for 5-6 days.
D0 (day 0) of the cycle is generally designated as the first day of behavioral estrus, which is the result of increasing estradiol-17β levels produced by the developing pre-ovulatory follicle. The rise in estradiol-17β levels and the maximum values before natural estrus in the doe are similar to that of the ewe (Chemineau et al., 1981). The rise of estradiol-17β is followed in less than 12 h by simultaneous peaks of Luteinizing Hormone (LH), Follicle Stimulating Hormone (FSH) and prolactin as reported for the ewe (Pant et al., 1977; Herriman et al., 1979; Cahill et al., 1981). These peaks occur 8.5 h after the onset of natural estrus (Chemineau et al., 1981). Within 48 h of the first peak, a second FSH peak occurs (Chemineau et al., 1981) which is similar in the ewe (Pant et al., 1977). High estradiol-17β levels are believed to cause a surge of Gonadotrophin Releasing Hormone (GnRH) and consequently an LH peak at estrus resulting in spontaneous ovulation towards the end of estrus. Following ovulation the ruptured follicle becomes a functional corpus luteum, which is the main source of progesterone in the cycling doe. Blood levels of progesterone are low at estrus (less than 1.0 ng mL-1) through to 2 days of diestrus and then rapidly increase to maximal levels at 7 days and remain elevated until 13-15 days in natural estrus (Fig. 1). Regression of the corpus luteum (luteolysis), induced by prostaglanding F2α (PGF2α), occurs if an embryo is not present in the uterus, resulting in a rapid drop in plasma progesterone (reviewed in Rahman, 2006). Ovarian oxytocin stimulates endometrial secretion and release of prostaglandins. The onset of the follicular phase of the next cycle is characterized by low progesterone levels and increasing GnRH and LH levels, while the FSH levels present at the onset of this phase are progressively decreased. These events are controlled by estrogen and inhibin, which are produced in increasing amounts by the developing follicles (Rahman, 2006).
Follicular dynamics, waves and dominant follicles: The mature ovary of farm animals contains varying numbers of follicles in different stages of development. Unlike other farm animals and ruminants, reports of caprine follicular waves and their association with hormones during estrous cycle are limited. The term follicular wave is defined as one or more antral follicles growing from 3 mm to ≥ 5 mm in diameter before regression (Ginther et al., 1995; de Castro et al., 1999; Bartlewski et al., 2000). Individual follicle emerging within a maximum of 48 h can be regarded as a single follicular wave (Medan et al., 2003). However, distinct differences exist in the follicular dynamics in the ovary among different farm animals. In cow and mares, distinct groups of follicles develop during the estrous cycle. Although 1 or 2 waves occur during the estrous cycle in mare by Pierson and Ginther (1987a), in cow 2 or 3 waves were detected (Pierson and Ginther, 1987b). The phenomenon called follicle dominance is generally present in each wave of follicular development, both in mare and in cow (Pierson and Ginther, 1987a, b, 1988; Savio et al., 1988; Sirois and Fortune, 1988). Dynamics of follicular development in ewe is a bit different and characterized by an initial wave of follicular activity at the beginning and another at the end of the estrous cycle (Bobes et al., 2003). However, the growth of ovarian follicles in the ovary of doe is characterized by the presence of 4 or more waves of follicle growth in the same cycle and it is in the final wave where the dominant follicle ovulates (Ginther and Kot, 1994; de Castro et al., 1999; Medan et al., 2005). Unlike other farm animals, the subsequent follicular wave begins even though the dominant follicle of the previous wave is still in its peak of development. This behavior strongly suggests that follicular dominance is less apparent in the ovary of doe (Ginther and Kot, 1994). Each follicular wave is preceded by an increase in FSH secretion (Medan et al., 2005). It is found that progesterone treatment affects follicular dynamics in dairy does (Menchaca and Rubianes, 2002).
Ovulation: Ovulation can be defined as the rupture of the mature ovarian follicle on the surface of the ovary and the release of its contents, including the maturing oocyte (Pineda, 2003) with adhering corona radiata cells, Cumulus Cells (CCs) and Follicular Fluid (FF). Ovulation is the most significant event of estrus. The point of ovulation can be seen in the resulting corpus luteum on the ovary days after ovulation. Ovulation is controlled by gonadotrophins: FSH is predominant during the phase of follicular growth and LH is generally regarded as ovulation inducer and also responsible for the formation of corpus luteum (Perry, 1971). In the doe, LH is released from the pituitary in a surge (50 ng mL-1) (Bono et al., 1983), which induces final preparation of the follicle 24 h prior to ovulation. At ovulation, LH level in the peripheral blood circulation of the doe reduces rapidly and FSH level begins to increase (Bono et al., 1983). Following rupture, external part of the follicle collapses and the follicular cavity becomes filled with clotted blood or serous fluid. The ruptured follicle reduces in size, the granulosa and theca internal cells begin to proliferate exuberantly under the influence of LH and form the corpus luteum (Sanga et al., 2002). Ovulation in the doe is spontaneous and most goat breed ovulates between 24-36 h after onset of estrus, the Nubian goat ovulates later, which is possibly due to a longer estrous cycle in this breed (Jainudeen et al., 2000). The average ovulation rate in the doe is 1-3 oocytes, but can vary from 1-5 depending upon the breeds and management conditions (Pineda, 2003).
ESTRUS SYNCHRONIZATION
Estrus synchronization is a key element of all the ART-protocols in livestock animals and has a major influence to increase the overall efficiencies of these programs (Baldassarre and Karatzas, 2004). Estrus synchronization plays a major role in fixed time breeding, AI, LOPU for oocyte or embryo collection and Embryo Transfer (ET). The value of estrus synchronization is vital in does as the duration of both estrous cycle and estrus is variable and estrus detection cannot be accomplished safely without a buck (Jainudeen et al., 2000). This technique has been developed in the early 1960s and since then a number of synchronizing methods has been developed for goats. Approaches towards synchronizing estrus in livestock have to focus on either the manipulation of the luteal or the follicular phase of the estrous cycle. In the doe, the window of opportunity is generally greater during the luteal phase, which is of longer duration and more responsive to manipulation. Different approaches have been concerned with either extending the luteal phase by supplying exogenous progesterone or with shortening this phase through regression of the corpus luteum. Successful techniques must not only establish synchrony, but also provide a reasonable level of fertility in the synchronized cycle.
A number of synchronization methods for goats have been evaluated under research conditions. The most widely used method in the doe is the treatment of progesterone or progestagen for 9-11 days followed by a luteolytic dose of prostaglandin (or an analogue) administered in the period 36-48h prior to removal of intravaginal sponge (Baldassarre and Karatzas, 2004). The progesterone or progestagen treatment can be delivered through an intravaginal sponge, a Controlled Internal Drug Release (CIDR) device or a subcutaneous implant (Evans and Maxwell, 1987; Ritar et al., 1989; Freitas et al., 1997). Although sponges are widely used either in conjunction with Pregnant Mare Serum Gonadotrophin (PMSG), FSH or prostaglandin to more tightly synchronize and/or induce a superovulatory response, but sponges are not preferred as these frequently cause discomfort and may adhere to the vaginal wall causing problems with removal (Holtz, 2005). An alternative means of supplying continuous, exogenous progesterone has been the CIDR developed for goats in New Zealand. The CIDR device is constructed from natural progesterone impregnated medical silicone elastomer molded over a nylon core. Currently the CIDR and subcutaneous implants are preferable than sponges because these are easy to use (Holtz, 2005). Ovulation in the does can be synchronized more precisely by administering GnRH around the time of estrus (Pierson et al., 2003), which improves the success of fixed-time AI and the collection of oocytes or embryos at a controlled stage of development for specific applications such as oocytes for In Vitro Production (IVP), Intracytoplasmic Sperm Injection (ICSI) or Somatic Cell Nuclear Transfer (SCNT) and zygotes for pronuclear microinjection (Baldassarre and Karatzas, 2004). Although, in the past, a considerable attention was focused in estrus synchronization, however, there is an urgent need for additional research conducted in a well-organized and systematic fashion to help establish guidelines for efficient breeding, AI, oocyte or embryo recovery and ET programs. Timeline of significant finding in estrus synchronization of does has been depicted in Table 2.
OVARIAN SUPEROVULATION
As multiple litter bearing animals, ovulation rate and litter size have a major impact on the reproductive efficiency of goat. Ovulation rate is influenced by the stage of breeding season, nutrition, genotype and parity. However, it can also be manipulated by pharmacological means which is known as superovulation. Superovulation is the hormonal treatment for increasing a large number of ova released by the ovary, which ultimately accelerate genetic improvement in any species. It is a means to induce maturation, ovulation and increase the number of ovulating or antral follicles (>2-3 mm) on the surface of the ovary available for oocyte recovery. Principles of inducing superovulation in doe are the same as in cow and ewe. An exogenous follicle-stimulating gonadotrophin is administered that mimics the effect of FSH near the end of the luteal phase of the cycle (9-11 days) or around 48 h before the end of the synchronizing treatments. There are a number of ways to superovulate doe, each of which has its advantages and disadvantages. However, superovulation in goats is frequently restricted by the cost of gonadotrophin or the handling requirements. The major commercial products applied are equine Chorionic Gonadotrophin (eCG) or PMSG and FSH used in higher (pharmacological) doses to elicit a superovulatory response; commercial preparations are partly purified from mare`s serum and porcine pituitary gland, respectively. PMSG is used to stimulate ovarian activity during seasonal anoestrus (Gordon, 1997) and usually used concurrently following estrus synchronization. PMSG is available in most countries and tended to be a practical hormone to administer for superovulation (Amoah and Gelaye, 1990). PMSG is preferred due to its lower cost and easy availability; it can be more easily administered than FSH, usually as a single injection of up to 1500-2000 IU, but the superovulatory response to PMSG can be quite variable and is usually lower than in a FSH-induced superovulation (Amoah and Gelaye, 1990). Problems associated with PMSG-induced superovulation are a high number of non-ovulated follicles, early regression of CL, short or irregular estrous cycles and potential risk of embryo expulsion (Amoah and Gelaye, 1990). A combination of eCG and human Chorionic Gonadotrphin (hCG) has also been widely used to superovulate does (Medan et al., 2003). FSH is a better choice of hormone for superovulating does as it provides more oocytes than PMSG. FSH is usually administered in decreasing doses of 1-5 mg, injected in 12 h intervals over a period of 3-5 days around the time of termination of the progestagen treatment. Like in cows and ewes, a number of experiments have been performed to compare the superovulatory response between FSH and PMSG, the evidence favors the use of FSH than PMSG (Tsunada and Sugie, 1989; Pendelton et al., 1992). In their study, Tsunada and Sugie (1989) reported that average number of oocyte recovery (OR) was significantly higher in FSH-treated does (9.4) than that in PMSG-treated ones (5.7). Studies in our laboratory also support of this finding. Earlier in our laboratory, PMSG alone or in combination with hCG was used to superovulate does. However, due to higher variability of stimulation and lower OR rate, a combination of recombinant ovine FSH (Ovagen™; ICPbio Limited, New Zealand) and hCG (Ovidrel; Laboratories Serono, Switzerland) as single doses was later introduced in our laboratory (Rahman et al., 2007a; Abdullah et al., 2008).
| Table 2: | Timeline of significant finding in estrus synchronization and superovulation in the goats |
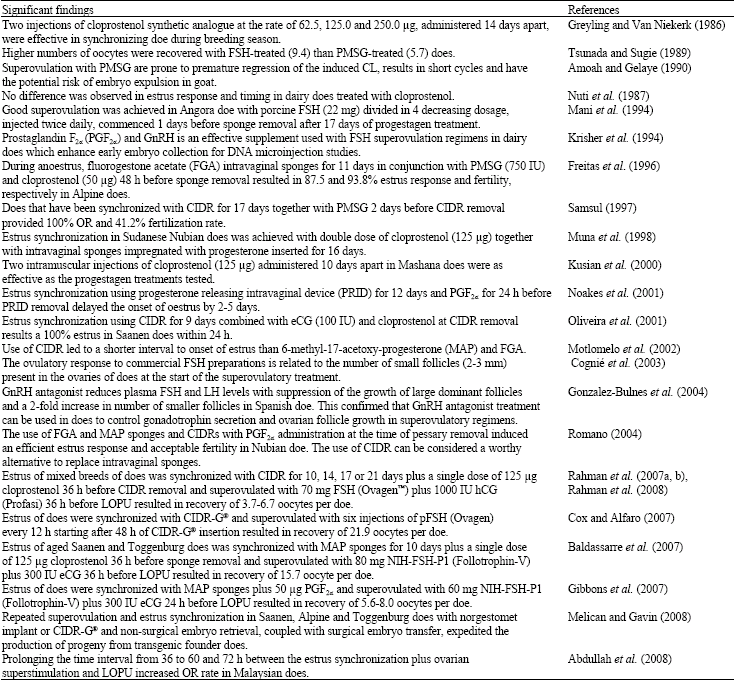 | |
Most superovulatory treatments are cumbersome and expensive and, over and above, accompanied by endocrine repercussions that take one or more subsequent cycles to subside (Holtz, 2005). Therefore, several attempts have been made to devise less labor-intensive treatment regimes without compromising oocyte or embryo yield. By applying a one shot-treatment regimen consisting of a single dose of FSH combined with a moderate dose of eCG (e.g., 60-80 mg FSH and 300IU eCG), Batt et al. (1993), Baldassarre et al. (2002, 2003), Baldassarre et al. (2007) and Gibbons et al. (2007) almost equaled the oocyte or embryo yield obtained with the traditional multiple injection regimen. The simplicity of this treatment is appealing. Of all the superovulation protocols in use to date, not a single one fulfils all expectations concerning predictability and reliability of the response. The variability in number of ovulations and yield of viable oocytes or embryos remains the main drawback (Holtz, 2005). Both intrinsic and extrinsic factors are responsible for the variability. Among the intrinsic factors, genetic (Nuti et al., 1987), age (Mahmood et al., 1991) and stage of the cycle at which the treatment applied (Wani et al., 1990) are important. A host of environmental factors such as season, nutrition, health state, AI (Holtz, 2005) and type of gonadotrophin administered (Gordon, 1997) are known to contribute to that variability. Therefore, vigorous research efforts are directed for the establishment of suitable superovulation regimes to augment the develoment of LOPU, ET and associated technologies based on them.
Time interval between the estrus synchronization plus superovulation treatment and LOPU may also influence the umber of oocytes recovered per doe (Abdullah et al., 2008). While other LOPU-IVP research groups (Baldassarre et al., 2002; Baldassarre et al., 2007) obtained optimum OR rates (13.4-15.7 oocytes per doe) after performing LOPU at 36 h of FSH plus hCG treatment, previous OR rates in our laboratory was always less than 7 oocytes per doe (Rahman et al., 2007b). In their experiment, Gibbons et al. (2007) used a lower time interval of 24 h between FSH plus eCG and LOPU and recovered lower OR rates (5.6-8.0 oocytes per doe). Therefore, we speculated that 36 h time interval between FSH plus hCG treatment and LOPU might not be optimum for proper stimulation of the ovarian follicles with our current protocol and, thus oocytes derived from these follicles had not acquired full meiotic competence and ooplasmic maturation. Keeping this in mind, we increased the time interval between FSH plus hCG treatment and the onset of LOPU from 36 h to 60 and 72 h and obtained 8.6 and 16.1 per doe, respectively, at 60 and 72 h interval (Abdullah et al., 2008). With slight modification of superstimulation protocol (decreasing hCG dose rate from 500 IU to 250 IU per doe), later we retrieved 14.9-17.6 oocytes per doe when LOPU performed 60 h post-FSH-hCG treatment (Rahman, 2008). These recovery rates are in agreement with Baldassarre et al. (2002), Baldassarre et al. (2003) and Baldassarre et al. (2007), who used slightly different protocol consisting of a single dose of FSH combined with a moderate dose of eCG (e.g., 80mg FSH and 300IU eCG). Using 60mg FSH and 300IU eCG in their superovulation protocol, (Gibbons et al., 2007) reported lower OR rate (5.6-8.0 oocytes per doe and 5.5-8.8 oocytes per ewe).
Dose rates of hormones for inducing superovulation might also have significant importance. Previous superovulation protocol in our laboratory was consisted of 70 mg FSH (Ovagen™; ICPbio Limited, New Zealand) and 1000 IU hCG (Ovidrel; Laboratories Serono, Switzerland) per doe (Rahman et al., 2007b). Later we have lowered the hormone doses from 70 to 35 mg FSH (Ovagen™) and 500 IU hCG (Ovidrel) per doe due to a shortage of hormones in the laboratory at that time (Abdullah et al., 2008). However, even with lower doses of hormones similar OR rate at 36 h and significantly higher OR rates at 60 and 72 h time intervals were obtained. Currently we further reduced the dose rate of hCG (Ovidrel) to 250 IU for 60 h time interval and OR rates increased to double (Rahman, 2008). It is not clear whether this increment of OR rates was the effect of time interval or hormonal dose rates or combined effects of time interval and hormonal dose rates. However, decreasing hormonal dose rates alone might not be responsible for increasing OR rates as both higher and lower dose rates of hormones at 36 h time interval provided similar OR rates in the previous studies in our laboratory. Timeline of significant finding in superovulation of does has been shown in Table 2.
CONCLUSIONS
The value of estrus synchronization is vital in goats as the duration of both estrous cycle and estrus is variable and estrus detection cannot be accomplished safely without a buck. Until now a number of studies have been performed to synchronize the estrous cycle of goats as well as to superovulate to increase the reproductive efficiency and genetic gain through the use of ARTs like Artificial Insemination (AI), Laproscopic Ovum Pick-Up (LOPU), In Vitro Production (IVP), Intracytoplasmic Sperm Injection (ICSI) or Somatic Cell Nuclear Transfer (SCNT). Through the application of findings from these studies reproductive performances of goat increased in some extent. Recently, estrus synchronization and superovulation received tremendous attention because of the rapid development of IVP and NT technologies. However, of all the estrus synchronization and superovulation protocols in use to date, not a single one fulfills all expectations concerning predictability and reliability of the response. The variability in number of ovulations and yield of viable oocytes or embryos remains the main drawback. Therefore, more studies are required to solve these problems.
ACKNOWLEDGMENTS
The authors wish to thank Islamic Development Bank (IDB) for providing an IDB Merit Scholarship to the first author. This work was supported by grants from MOSTI Special Project (Grant Number 01-02-03-0696) and IPPP (Grant Number Vote F-0179/2004D, 0145/2005D and P0170/2006C).
REFERENCES
- Abdullah, R.B., S.L. Liow, A.N.M.A. Rahman, W.K. Chan, W.E. Wan-Khadijah and S.C. Ng, 2008. Prolonging the interval from ovarian hyperstimulation to laparoscopic ovum pick-up improves oocyte yield, quality, and developmental competence in goats. Theriogenology, 70: 765-771.
CrossRefPubMedDirect Link - Attwood, B.M., 2007. An Introduction to Reproduction Management of Fibre and Meat Goats. 2nd Edn., Department of Primary Industries, Melbourne, Australia,.
Direct Link - Baldassarre, H., K.M. Rao, N. Neveu, E. Brochu, I. Begin, E. Behboodi and D.K. Hockley, 2007. Laparoscopic ovum pick-up followed by in vitro embryo production for the reproductive rescue of aged goats of high genetic value. Reprod. Fertil. Dev., 19: 612-616.
CrossRef - Baldassarre, H., B. Wang, N. Kafidi, M. Gauthier and N. Neveu et al., 2003. Production of transgenic goats by pronuclear microinjection of in vitro produced zygotes derived from oocytes recovered by laparoscopy. Theriogenology, 59: 831-839.
CrossRef - Baldassarre, H., B. Wang, N. Kafidi, C. Keefer, A. Lazaris and C.N. Karatzas 2002. Advances in the production and propagation of transgenic goats using laparoscopic ovum pick-up and in vitro embryo production technologies. Theriogenology, 57: 275-284.
CrossRef - Bartlewski, P.M., A.P. Beard and N.C. Rawlings, 2000. An ultrasound-aided study of temporal relationships between the patterns of LH/FSH secretion, development of ovulatory-sized antral follicles and formation of corpora lutea in ewes. Theriogenology, 54: 229-245.
CrossRef - Batt, P.A., I.D. Killeen and A.W. Cameron, 1993. Use of single or multiple injections of FSH in embryo collection programmes in goats. Reprod. Fertil. Dev., 5: 49-56.
Direct Link - Bobes, R.J., M. Perez-Martinez, Y. Gomez and M.C. Romano, 2003. Metabolism of progesterone to estrogens and androgens by individual follicles of the goat ovary. Small Rumin. Res., 47: 233-242.
CrossRefDirect Link - Bono, G., F. Cairoli, G. Tamanini and L. Arbrate, 1983. Progesterone, oestrogen, luteinising hormone, follicle stimulating hormone and prolactin concentrations in plasma during oestrous cycle in goat. Reprod. Nutr. Dev., 23: 217-222.
Direct Link - Cahill, L.P., J. Saumande, J.P. Ravault, M.R. Blanc and J. Thimonier et al., 1981. Hormonal and follicular relationships in ewes of high and low ovulation rates. J. Reprod. Fertil., 62: 141-150.
PubMed - Chemineau, P., D. Gauthier, J.C. Poirier and J. Saumande, 1982. Plasma levels of LH, FSH, prolactin, oestradiol-17β and progesterone during natural and induced oestrus in the dairy goats. Theriogenology, 17: 313-323.
CrossRef - Cognie, Y., G. Baril, N. Poulin and P. Mermillod, 2003. Current status of embryo technologies in sheep and goat. Theriogenology, 59: 171-188.
PubMedDirect Link - Cox, J.F. and V. Alfaro, 2007. In vitro fertilization and development of OPU derived goat and sheep oocytes. Reprod. Dom. Anim., 42: 83-87.
CrossRef - De Castro, T., E. Rubianes, A. Menchaca and A. Rivero, 1999. Ovarian dynamics, serum estradiol and progesterone concentrations during the interovulatory interval in goats. Theriogenology, 52: 399-411.
CrossRef - Freitas, V.J., G. Baril and J. Saumande, 1996. Induction and synchronization of estrus in goats: The relative efficacy of one versus two fluorogestone acetate-impregnated vaginal sponges. Theriogenology, 46: 1251-1256.
CrossRef - Freitas, V.J., G. Baril and J. Saumande, 1997. Estrus synchronization in dairy goats: Use of fluorogestone acetate vaginal sponges or norgestomet ear implants. Anim. Reprod. Sci., 46: 237-244.
CrossRef - Gibbons, A., F.P. Bonnet, M.I. Cueto, M. Catala and D.F. Salamone et al., 2007. Procedure for maximizing oocyte harvest for in vitro embryo production in small ruminants. Reprod. Domest. Anim., 42: 423-426.
CrossRef - Ginther, O.J. and K. Kot, 1994. Follicular dynamics during the ovulatory season in goats. Theriogenology, 42: 987-1001.
CrossRef - Ginther, O.J., K. Kot and M.C. Wiltbank, 1995. Associations between emergence of follicular waves and fluctuations in FSH concentrations during the estrous cycle in ewes. Theriogenology, 43: 689-703.
CrossRef - Gonzalez-Bulnes, A., J. Santiago-Moreno, R.M. Garcia-Garcia, C.J.H. Souza and A. Lopez-Sebastian et al., 2004. Effects of GnRH antagonist treatment on gonadotrophin secretion, follicular development and inhibin a secretion in goats. Theriogenology, 61: 977-985.
CrossRef - Greyling, J.P.C. and C.H. Van Niekerk, 1986. Synchronisation of oestrus in the Boer doe: Dose effect of prostaglandin in the double injection regimen. South Afr. J. Anim. Sci., 16: 146-150.
Direct Link - Herriman, I.D., D.J. Harwood, C.B. Mallinson and R.J. Heitzman, 1979. Plasma concentrations of ovarian hormones during the oestrous cycle of the sheep and cow. J. Endocrinol., 81: 61-64.
PubMed - Krisher, R.L., F.S. Gwasdaukas, R.L. Page, C.G. Russell and R.S. Canseco et al., 1994. Ovulation rate, zygote recovery and follicular populations in FSH-superovulated goats treated with PGF2a and or GnRH. Theriogenology, 41: 491-498.
CrossRef - Kusina, N.T., F. Tarwirei, H. Hamudikuwanda, G. Agumba and J. Mukwena, 2000. A comparison of the effects of progesterone sponges and ear implants, PGF2α and their combination on efficacy of estrus synchronization and fertility of mashona goat does. Theriogenology, 53: 1567-1580.
CrossRefPubMedDirect Link - Mahmood, S., G.L. Kaul and J.C. Biswas, 1991. Comparative efficacy of FSH-P and PMSG on superovulation in Pashmina goats. Theriogenology, 35: 1191-1196.
CrossRef - Mani, A.U., E.D. Watson and W.A.C. McKelvey, 1994. The effects of subnutrition before and after embryo transfer on pregnancy rate and embryo survival in does. Theriogenology, 41: 1673-1678.
CrossRef - Medan, M.S., G. Watanabe, K. Sasaki, N.P. Groom and S. Sharawy et al., 2005. Follicular and hormonal dynamics in during estrous cycle in goats. J. Reprod. Dev., 51: 455-463.
CrossRef - Medan, M.S., G. Watanabe, K. Sasaki, S. Sharawy and N.P. Groome et al., 2003. Ovarian dynamics and their associations with peripheral concentrations of gonadotrophins, ovarian steroids and inhibin during the estrous cycle in goats. Biol. Reprod., 69: 57-63.
Direct Link - Melican, D. and W.G. Gavin, 2008. Repeat superovulation, non-surgical embryo recovery, and surgical embryo transfer in transgenic dairy goats. Theriogenology, 69: 197-203.
CrossRef - Menchaca, A. and E. Rubianes, 2002. Relation between progesterone concentrations during the early luteal phase and follicular dynamics in goats. Theriogenology, 57: 1411-1419.
PubMed - Motlomelo, K.C., J.P.C. Greyling and L.M.G. Schwalbach, 2002. Synchronisation of oestrus in goats: The use of different progestagen treatments. Small Rumin. Res., 45: 45-49.
CrossRefDirect Link - Ahmed, M.M.M., S.E. Makawi and A.S. Jubara, 1998. Synchronization of estrus in Nubian goats. Small Rum. Res., 30: 113-120.
CrossRef - Nuti, L.C., B.S. Minhas, W.C. Baker, J.S. Capehart and P. Marrack, 1987. Superovulation and recovery of zygotes from Nubian and Alpine dairy goats. Theriogenology, 28: 481-488.
CrossRef - Pant, H.C., C.R.N. Hopkinson and R.J. Fitzpartick, 1977. Concentration of oestradiol, progesterone, luteinizing hormone and follicle stimulating hormone in the jugular plasma of ewes during the oestrous cycle. J. Endocrinol., 73: 247-255.
PubMed - Pierson, J.T., H. Baldassarre, C.L. Keefer and B.R. Downey, 2003. Influence of GnRH administration on timing of the LH surge and ovulation in dwarf goats. Theriogenology, 60: 397-406.
CrossRef - Pierson, R.A. and O.J. Ginther, 1987. Follicular populations during the estrous cycle of the mare. Anim. Reprod. Sci., 14: 219-231.
CrossRef - Pierson, R.A. and O.J. Ginther, 1987. Follicular populations during the oestrous cycle in heifers. I. Influence of day. Anim. Reprod. Sci., 14: 165-176.
CrossRef - Pierson, R.A. and O.J. Ginther, 1988. Ultrasonic imaging of the ovaries and uterus in cattle. Theriogenology, 29: 21-37.
CrossRefDirect Link - Rahman, A.N.M.A., 2006. Hormonal changes in the uterus during pregnancy-lessons from the ewe: A review. J. Agric. Rural Dev., 4: 1-8.
Direct Link - Rahman, A.N.M.A., R.B. Abdullah and W.E. Wan-Khadijah, 2007. Longer time intervals between chorionic gonadotrophin treatment and LOPU, but not the LOPU cycles, have positive effect on goat oocyte recovery. Proceedings of the 12th Biological Sciences Graduate Conference, December 17-19, 2007, Kuala Lumpur, Malaysia, pp: 97-97.
- Rahman, A.N.M.A., R.B. Abdullah and W.E. Wan Khadijah, 2007. Goat embryo development from in vitro matured oocytes of heterogeneous quality through intracytoplasmic sperm injection technique. Biotechnology, 6: 373-382.
CrossRefDirect Link - Rahman, A.N.M.A., R.B. Abdullah and W.E.W. Khadijah, 2008. A review of reproductive biotechnologies and their application in goat. Biotechnology, 7: 371-384.
CrossRefDirect Link - Ritar, A.J., S. Salamon, P.D. Ball and P.J. O'May, 1989. Ovulation and fertility in goats after intravaginal device-PMSG treatment. Small Rum. Res., 2: 323-331.
CrossRef - Sanga, G.K., R.K. Sharma and S.S. Guraya, 2002. Biology of corpus luteum in small ruminants. Small Rum. Res., 43: 53-64.
CrossRef - Savio, J.D., L. Keenan, M.P. Boland and J.F. Roche, 1988. Pattern of growth of dominant follicles during the estrous cycle in heifers. J. Reprod. Fertil., 83: 663-671.
PubMed - Sirois, J. and J.E. Fortune, 1988. Ovarian follicular dynamics during the estrous cycle in heifers monitored by real-time ultrasonography. Biol. Reprod., 39: 308-317.
Direct Link - Tsunada, Y. and T. Sugie, 1989. Superovulation in nonseasonable Japanese native goats, with special reference to the developmental progression of embryos. Theriogenology, 31: 991-996.
CrossRef - Romano, J.E., 2004. Synchronization of estrus using CIDR, FGA or MAP intravaginal pessaries during the breeding season in Nubian goats. Small Ruminant Res., 55: 15-19.
CrossRef
pitamber Reply
very excellent work
A N M Aminoor Rahman
Thank you very much Sir