
A.N.M.A. Rahman
Animal Biotechnology-Embryo Laboratory (ABEL), Institute of Biological Sciences, University of Malaya, Lembah Pantai, 50603 Kuala Lumpur, Malaysia
R.B. Abdullah
Animal Biotechnology-Embryo Laboratory (ABEL), Institute of Biological Sciences, University of Malaya, Lembah Pantai, 50603 Kuala Lumpur, Malaysia
W.E. Wan-Khadijah
Animal Biotechnology-Embryo Laboratory (ABEL), Institute of Biological Sciences, University of Malaya, Lembah Pantai, 50603 Kuala Lumpur, Malaysia
Journal of Biological Sciences
Year: 2008 | Volume: 8 | Issue: 7 | Page No.: 1115-1128







ABSTRACT
Embryo in vitro Production (IVP) and associated Assisted Reproductive Technologies (ARTs) in goat, e.g., estrus synchronization and superovulation, Laparoscopic Ovum Pick-up (LOPU), in vitro Maturation (IVM), in vitro Fertilization (IVF), Intracytoplasmic Sperm Injection (ICSI) and in vitro Culture (IVC) attained significant attention in the recent years. However, for the success in any IVP protocols in goat sound knowledge of physiology of gametogenesis, fertilization and early embryogenesis in vivo is crucial because in vivo information form the basis and guide for any in vitro experiment. Unlike human, laboratory animals, cattle and sheep, fewer studies have been conducted in gametogenesis, fertilization and early embryo development in goat. Data for sheep and cattle are mostly used as a basis for goat IVP studies. Therefore, the current study is intended to review gametogenesis, sperm oocyte interaction, fertilization and early embryogenetic process in mammals with special reference to goat.
PDF Abstract XML References Citation
How to cite this article
A.N.M.A. Rahman, R.B. Abdullah and W.E. Wan-Khadijah, 2008. Gametogenesis, Fertilization and Early Embryogenesis in Mammals with Special Reference to Goat: A Review. Journal of Biological Sciences, 8: 1115-1128.
DOI: 10.3923/jbs.2008.1115.1128
URL: https://scialert.net/abstract/?doi=jbs.2008.1115.1128
DOI: 10.3923/jbs.2008.1115.1128
URL: https://scialert.net/abstract/?doi=jbs.2008.1115.1128
INTRODUCTION
In vitro Embryo Production (IVEP) or in vitro Production (IVP) technique is currently in the central focus among all the Assisted Reproductive Technologies (ARTs) in human as well as in animals. Therefore, the physiology of normal fertilization should be considered as a must for the development of any IVEP or IVP protocols especially for in vitro Fertilization (IVF) or Intracytoplasmic Sperm Injection (ICSI) techniques. Gametogenesis, fertilization and early embryogenesis or pre-implantation embryo development are crucial periods for normal development of an embryo afterwards. Until now researches in mammalian gametogenesis, fertilization and early embryogenesis mainly based on the laboratory animals and human (Tulsiani et al., 1997; Kupker et al., 1998; Elder and Dale, 2000). Compared with laboratory and farm animals like cattle, sheep and pig, studies in goat are scarce (Hafez and Hafez, 2000a; Miyano and Hirao, 2003). Therefore, the current paper will briefly review some aspects of gametogenesis, fertilization and early embryogenesis in mammals with special reference to goat.
GAMETOGENESIS
Genetically and functionally competent gametes are a prerequisite for normal fertilization and early embryo development. The first phase in the sexual reproduction of an organism is gametogenesis, a process of formation of gametes from the germ cells in the testes and ovaries. This process is termed as spermatogenesis in the male and oogenesis in the female. It is the fundamental biological process in both the sexes and the key event of gametogenesis is the halving of the number of chromosomes to produce haploid germ cells (sperm and oocyte) through meiosis. Thus, in goat, where the chromosome number of somatic cells is 60, each sperm and each oocyte has only 30 chromosomes. However, until this point spermatogenesis and oogenesis resume their similarity. After this, in the male, each primary spermatocyte divides meiotically and produces four spermatids, each destined to become a functional sperm. In the female, on the other hand, of the four cells produced from each primary oocyte only one finally becomes a functional oocyte. A schematic diagram of gametogenesis in mammals has been illustrated in Fig. 1.
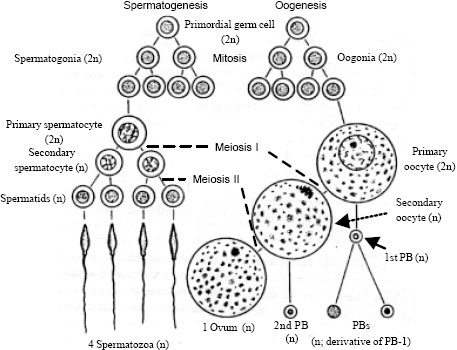 | |
| Fig. 1: | Schematic diagram of gametogenesis in mammals. Modified from Hickman et al. (1986) |
In mammals, the male and female gametes originate from the embryonic yolk sac. The gametes initially colonize in the primitive gonadal ridge at the early stage of pregnancy by migration through the developing mesentery of the embryo, where germ cells associate with somatic cells; it continues with their multiplication, growth and maturation. Finally, migrate to the pelvic and inguinal regions to form the ovary or testis, respectively and ends at fertilization. Mammalian spermatogenesis and oogenesis are briefly described below:
Spermatogenesis: Competent sperm are required for the successful contribution of the paternal genotype to embryo development. The process of spermatogenesis results in the formation of the haploid male gamete required for fertilization of an oocyte. Spermatogenesis is a continual and complex process that involves three major steps: (a) proliferation- multiplication of spermatogonia by the process of mitosis (spermatocytogenesis); (b) meiosis (spermiogenesis)- meiotic divisions whereby the chromosome number is reduced from diploid to haploid and (c) differentiation- transformation of the round spermatid into the complex structure of the spermatozoon (reviewed in Barth and Oko, 1989; De Kretser et al., 1998; Elder and Dale, 2000). Spermatogenesis in buck and other male mammals begins at puberty through the proliferation of interphase germ cells, continues throughout adult life and takes place inside the seminiferous tubules of the testes. The age of puberty for male goat or buck ranged between 4 and 6 months (Jainudeen et al., 2000). The walls of the seminiferous tubules contain the differentiating sex cells arranged in a stratified layer, 5 to 8 cells deep. The outermost layers contain spermatogonia, which have increased in number by spemtocytogenesis. The spemtocytogenesis begins with mitosis of the diploid A spematogonia in the basal compartment of the Sertoli cells. The A spematogonia differentiate into B spermatogonia which enter their final mitotic division prior to their entrance into the pre-leptotene phase of first meiosis (meiosis 1). Meiosis occurs in the adluminal compartment of the Sertoli cell and results in the formation of primary spermatocytes. Condensation of chromosomes occurs during the leptotene phase followed by the zygotene phase. The long pachytene phase involves the crossing over of chromosomal material and is most susceptible to damage. Meiosis progresses through the diplotene phase, diakinesis, Metaphase I (MI) and anaphase, finally resulting in secondary spermatocytes (2c, 1n). Each secondary spermatocyte then enters the second meiotic division (meiosis 2), without the intervention of a resting period, resulting in spermatids with a haploid number of chromosomes and DNA content (1c, 1n), required for fertilization. The round-shaped spermatids proceed through spermiogenesis and metamorphically changed into highly specialized motile cells. Spermiogenesis consists of the Golgi phase, the acrosomal cap phase, the acrosomal phase and the complex maturation stage involves development of the sperm tail (reviewed by Barth and Oko, 1989). Deviations from the process of normal spermatogenesis result in abnormal morphology or dysfunction of the sperm cell. The progression from spermatogonia to mature spermatozoa in mammals require approximately 60 to 70 days, with at least three mitotic and two meiotic divisions during spermatogenesis (Kupker et al., 1998). Finally, the newly formed sperm are released into the lumen of the seminiferous tubules. These sperm are immotile and still immature in terms of fertilizing capability. Therefore, to acquire motility as well as maturation they need to pass down the epididymis to the ejaculatory duct, which is known as epididymal maturation (Elder and Dale, 2000). During epididymal maturation, the sperm shed the cytoplasmic droplet, undergo modifications in the protein, carbohydrate and glycoprotein composition of the plasma membrane and acquire a net negative charge (Yanagimachi, 1994; Harrison, 1996). Sperm motility and capacitation are suppressed during storage in the cauda epididymis, which is characterized by low pH, low Ca2+, low Na+ and increased K+ (Dalvit et al., 1995; Jones and Murdoch, 1996). Disruptions in the epididymal environment may result in abnormalities of sperm function. Immediately after ejaculation, the sperm are incapable of fertilizing the oocyte and, therefore, to acquire fertilization potential, they need to undergo functional changes (or capacitation), which occur inside the female reproductive tract.
Although in comparison with the oocytes, the spermatozoa are very small, however, they are essentially very long and compact cells with a few highly specialized cytoplasmic structures, the flagellum for motility and the acrosome, which is instrumental in sperm-oocyte binding and fusion. Morphologically and functionally, a goat spermatozoon is composed of four regions: (a) the head containing nucleus and the acrosome, (b) the neck containing centrioles, (c) the middle piece containing mitochondria and d) the tail piece or flagellum. A goat spermatozoon is approximately 60 μm long of which head and tail piece are 7.69 and 52 μm long, respectively (Gravance et al., 1995). Frozen-thawed goat spermatozoa from proven quality Jermasia buck (Jermasia is a synthetic breed developed by the University of Malaya) have been shown in Fig. 2a, b.
Oogenesis: The maternal contribution to the development of the embryo is determined during formation and maturation of the female gamete, the oocyte. The ability of the oocyte to achieve sperm-oocyte fusion is acquired early in oogenesis or the process of oocyte formation. The oogenetic products synthesized during oocyte growth must also be sufficient to support embryonic development from fertilization until the activation of the embryonic genome (Olszanska and Borgul, 1993). Ultimately, the nuclear and ooplasmic maturity of the oocyte influences the success of fertilization and embryo development. In mammals, oogenesis commences during early fetal development, stops at birth and continues during puberty throughout the reproductive life of the female. After continuation of meiosis, the oogenesis process until completion is very fast. Oogenesis in mammals includes seven steps: (a) generation of primordial germ cells (PGCs), (b) migration of PGCs to the prospective gonads, (c) colonization of the gonads by PGCs, (d) differentiation of PGCs to oogonia, (e) proliferation of oogonia, (f) initiation of meiosis and (g) arrest at the diplotene stage of first meiotic prophase or prophase 1 (reviewed in Van den Hurk and Zhao, 2005).
Oogonia are the early germ cells in the ovary, which increase in number by mitosis. Oogonial multiplication begins during early fetal development and ends months to years later in the sexually mature adult (Picton et al., 1998). Once mitosis ceases, the oogonia then grow in size and enter the prophase of the first meiotic division at approximately day 55 of gestation in the ewe (McNatty et al., 1995) and are then referred to as primary oocytes (Wassarman and Albertini, 1994). Each oogonium or primary oocyte contains the diploid number of chromosome. The primary oocyte which is transformed from each oogonium is a cell which becomes enclosed in a follicle, known as primordial follicle. In goat, sheep and cow, large population (approximately 100,000) of primordial or pre-antral follicles with meiotically incompetent oocytes are present in the ovaries (Miyano, 2003; Miyano and Hirao, 2003; Zhou and Zhang, 2006). Most of them are lost at various stages of development owing to atresia and only a very minority of oocytes becomes available for ovulation. In contrast, Ariyaratna and Gunawardana (1997) found in their histological study that one pair of ovaries of Batu goat (a Sri Lankan goat breed) aged between 18 to 36 months contained 35,092 primordial follicles (which are 90% of total ovarian follicle population), 10.67 normal and 20.42 atretic vesicular (antral) follicles (1-6 mm diameter). At birth, all oocytes from growing and dominant follicles are arrested at the diplotene stage of prophase 1 (Van den Hurk and Zhao, 2005). This dictyate stage is characterized by the enclosure of the chromosomes within the large nucleus, also known as the Germinal Vesicle (GV) (Elder and Dale, 2000). The oocytes remain in the arrested state until a few hours before ovulation. Surprisingly, the oocytes may stay at this arrested stage for a longer period of time depending on the species, waiting for the signal to resume growth and subsequent development occurs at puberty. The age of puberty for female goat or doe is ranged between 5 and 7 months (Jainudeen et al., 2000). The reason for storing the oocytes in this remarkable frozen meiotic state is unknown (Johnson and Everitt, 1980).
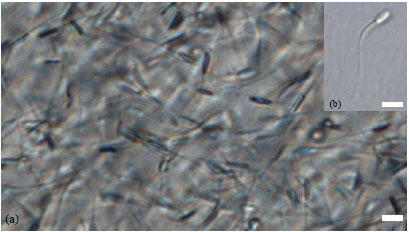 | |
| Fig. 2: | Microphotographs of goat sperm. (a) Frozen-thawed goat sperm from Jermasia buck of proven quality, (b) a single motile normal goat sperm before being immobilized for ICSI procedure. Scale bar represents 10 μm |
Oocyte growth and development: The growth and development of an oocyte occurs inside an ovarian follicle and oocyte undergoes a progressive series of morphological modifications as it grows and proceeds through the different stages of development (Eppig et al., 1994). Although data are lacking for doe, Ariyaratna and Gunawardana (1997) indicated from their study that follicular morphology and activity are similar in does and ewes. In the ewe, primordial, primary and secondary follicles, respectively, appear in the fetal ovary at days 75, 100 and 120 (McNatty et al., 1995). Once a primordial follicle oocyte is activated to grow, it embarks on a complex journey that involves numerous molecular and morphological changes to both the oocyte and the follicle. The modifications are carefully orchestrated and require sensitive communication between the oocyte and surrounding Granulosa Cells or GCs (Fair, 2003). These structural rearrangements facilitate the increasing energy and nucleic acid synthesis requirements of the developing oocyte and are a prerequisite to the oocyte achieving meiotic competence and embryo developmental potential. The first sign of morphological change when the oocyte begins to grow is turning of the flat GCs to cuboidal which is known as primary follicle. After completion of the morphological change, the GCs proliferate actively, which cause the follicles to develop and increase in size. Through a series of mitotic division of GCs, unilaminar primary follicles are converted to multilaminar secondary follicles, followed by the antral or tertiary follicles (Miyano and Hirao, 2003). The antral follicle is a highly complex unit consisting of several layers of GC surrounding a fluid-filled cavity or ‘antrum` in which the oocyte surrounded by somatic cells is bathed. In the doe, antrum formation began when the GCs are about six cell layers in thickness and the Zona Pellucida (ZP) is visible at this stage (Ariyaratna and Gunawardana, 1997). The fluid found in the ‘antrum` is known as Follicular Fluid (FF). During this growth phase there is a major increase in ooplasmic organelles. The follicle provides a microenvironment for oocyte growth, development and is responsible for the production of hormones. The walls of mature preovulatory follicles consist of membrana granulosa (mural granulosa), theca interna and theca externa. The GCs are cells of epithelial origin essential for the growth and survival of the oocyte. The GCs consist of the corona radiata; Cumulus Cells (CCs), membrana granulosa and antral granulosa cells. The CCs surround the oocyte, which nourish the oocyte, are involved in oocyte growth, maturation (Buccione et al., 1990) and participate in the formation of the ZP (made of a translucent acellular layer of glycoprotein). In addition, these cells have also been implicated in the modulation or generation of oocyte maturation inhibitors (Tsafriri et al., 1982; Eppig and Downs, 1984). The CCs in close contact with the oocyte are known as corona radiata. They are in close contact with the oocyte through ooplasmic extensions or processes across the ZP (De Loos et al., 1991), which are known as gap junctions. The heterologous gap junctions provide the basis for extensive network of intracellular communication among GCs. Normally CCs or corona cells surrounding the goat oocyte shed ≥30 h after ovulation (Harper, 1982).
Oocyte maturation: As mentioned earlier that oocytes are arrested at the diplotene stage of the prophase 1 at birth, they resume meiosis after a long quiescent phase at puberty which involve sequential sub-cellular and molecular transformations by various components of the follicle. During postnatal life, starting from puberty, ovarian follicles continue to grow, mature and either ovulate or regress. Follicles are recruited continuously until the original store is exhausted. Reinitiation of meiosis in the fully-grown oocyte is the first sign of oocyte maturation, which involves condensation of interphase chromatin, breakdown of nuclear membrane (germinal vesicle breakdown: GVBD), spindle formation and chromosome segregation. In vivo, resumption of meiosis is initiated by a preovulatory LH surge and only occurs in fully grown, meiotically competent oocytes from dominant follicles. Small oocytes in primordial and primary follicles have no ability to resume meiosis. Oocytes acquire the competence to resume meiosis when their size exceeds 80% of their final diameter; they then become gradually competent to progress to metaphase 2 (MII) as the diameter increases to over 90% of the maximum (Miyano and Hirao, 2003). The diameter of mature goat oocytes excluding ZP (ooplasm) ranged from 119-146 μm (De Smedt et al., 1994; Crozet et al., 2000). Diameter of mature oocyte in different animals and human is presented in Table 1. During this follicle and oocyte growth phase, oocytes not only acquire competency to resume meiosis, but also acquire ooplasmic maturity, also known as oocyte capacitation, both of which are required to ensure normal fertilization and embryo development (Gosden et al., 1997; Hyttel et al., 1997). From in vitro studies, it is found that goat oocytes acquired the ability to initiate meiotic resumption in early antral follicles of 0.5 to 0.8 mm in diameter and to reach MI in follicles of 1.0 to 1.8 mm in diameter (De Smedt et al., 1994). Although 86% of goat oocytes from follicles larger than 2 mm progress to MII (De Smedt et al., 1992), only a small proportion of them can support embryonic development (Crozet et al., 1993). In cattle, oocytes originating from follicles larger than 6 mm in diameter yield a significantly higher percentage of blastocyst than the smaller follicles (Lonergan et al., 1994). In an in vitro study, a significantly higher oocyte maturation, morula and blastocyst development rates were achieved with goat oocytes originating from larger than 5 mm follicles compared with medium (>3-5) and smaller (2-3) follicles (Crozet et al., 1995). To reach the maturity, goat oocytes (ooplasm) grow from 29.6 μm in primordial follicles to 119 to 146 μm (De Smedt et al., 1994; Crozet et al., 2000) in antral follicles (>2 mm in diameter). The ZP of a goat oocyte from antral follicle bigger than 2 mm is about 3.2 μm thick (Ariyaratna and Gunawardana, 1997). The growth of goat oocyte in relation to follicular growth in the ovary as described by Ariyaratna and Gunawardana (1997) has been presented in Table 2.
Before and at the time of LH surge, the oocyte is surrounded by a compact CCs investment. The oocyte undergoes a series of changes in its nucleus, ooplasm and organization of the plasma membrane (oolemma) during the period between the LH surge and ovulation, which is known as oocyte maturation. Completion of the meiosis 1 takes place when oocytes have undergone extensive growth in cellular interaction with GCs and theca cells. The oocyte undergoes asymmetric cytokinesis and extrudes the first polar body (PB-1) containing a haploid chromosome complement (Kupker et al., 1998). Immediately after the meiosis 1 is completed, the meiosis 2 is initiated and the oocytes are again arrested at MII stage until fertilization, when an activation stimulus provided by sperm penetration triggers the completion of the meiotic cycle and initiates embryonic development. Ooplasmic maturation is required to acquire to the conditions to block polyspermy in case of fertilization, to decondense penetrated spermatozoon and to form pronucleus (PN). It includes redistribution of cell organelles, migration of mitochondria to perinuclear position and accumulation of granules along the oolemma (Van den Hurk and Zhao, 2005). The endpoint of this, in vivo, is the ovulation and release from the follicle of a MII oocyte with potential to support normal embryonic development (Elder and Dale, 2000). However, the doe ovulates two to three mature oocytes in each estrous cycle (Jainudeen et al., 2000). Compared with human or mouse, goat oocyte is dark in opacity (Betteridge, 2003) due to very dense ooplasm consisted of concentrated lipid materials (Keskintepe et al., 1997). The ooplasmic opacity in different animals and human has been presented in Table 1. A developmentally competent immature goat Cumulus-Oocyte Complex (COC) and mature goat oocyte have been illustrated, respectively, in Fig. 3a, b.
| Table 1: | Mature oocyte size and opacity in different animals and human |
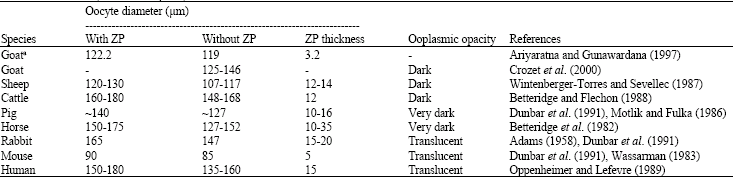 | |
| Oocyte diameter without ZP (ooplasmic diameter) was calculated by deducting average ZP thickness from oocyte diameter with ZP. aBelongs to Batu breed (a Srilankan goat breed) | |
| Table 2: | Growth of goat (Batu breed) oocyte (Mean ± SD) in relation to follicular growth in the ovary |
 | |
| In each column, values with different superscript letters are different (p<0.05) | |
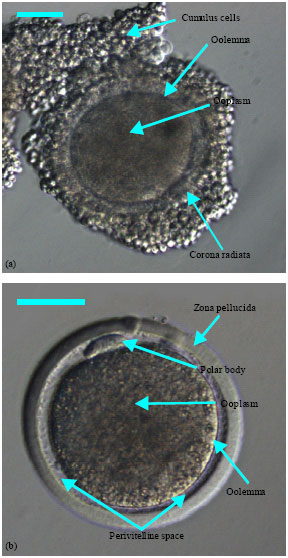 | |
| Fig. 3: | Developmentally competent (a) immature and (b) mature goat oocytes recovered through Laparoscopic Ovum Pick-Up (LOPU) technique from estrus synchronized and superstimulated doe. Scale bar represents 50 μm |
ACTIVATION OF SPERM AND SPERM-OOCYTE INTERACTION
Fertilization encompasses a series of different steps which have to be performed in a well orchestrated way to create a new individual. These include sperm capacitation, sperm binding and penetration of the ZP, traversing the perivitelline space (PVS), binding and fusion with oolemma, activation of the oocytes and decondensation of the sperm head to form the Male PN (MPN). In mammals, fertilization process is internal and the male gametes (spermatozoa) must be introduced into the female reproductive tract at coitus or artificially. As already mentioned, when the ejaculated spermatozoa are deposited into the female reproductive tract at that time they have no fertilizing capability. Therefore, the spermatozoa need to gain this ability, or in other word, need to be activated or made in ready state first before they can interact with oocyte to ensure fertilization success. The known physiological events occurring during sperm activation and sperm-oocyte interaction have been reviewed mainly based on mice models (Bedford, 1982; Yanagimachi, 1994; Tulsiani et al., 1997; Bedford, 1998).
Activation of spermatozoa: In mammalian species, activation of spermatozoa follows several physiological and structural changes upon exposure to environmental signals in the female reproductive tract before they interact with the oocytes towards a successful fertilization. These include various alterations in the plasma membrane and intracellular components and changes in motility pattern and metabolism of the spermatozoa. Two main processes, namely capacitation and acrosome reaction, must occur to activate the spermatozoa before the sperm interact with the oocyte.
Capacitation: The spermatozoa must reside a minimum period in the female reproductive tract before gaining the ability to fertilize oocytes (Austin, 1951; Chang, 1951). It is speculated that during this time, glycoproteins from the sperm surface are removed, thus exposing receptor sites that can respond to oocyte signals and lead to acrosome reaction. This process allows the spermatozoa to bind to the ZP, which subsequently leads to the Acrosome Reaction (AR), penetration of the ZP and fertilization of the oocyte. In another word, the spermatozoa gains the capacity to fertilize oocyte and, therefore, is termed as capacitation (reviewed in Markert, 1983; Elder and Dale, 2000). This phenomenon is first noted by Chang (1951) and Austin (1951) and the term ‘capacitation` was referred by Austin (1951). The time required for sperm capacitation varies among different species of animals and ranges from less than one hour in male mouse to 6 h in man (Elder and Dale, 2000). The changes that occur during capacitation in the sperm plasma membrane include modification of ion channels, increased adenylate cyclase and cyclic adenosine monophosphate (cAMP), changes in surface glycoprotein moieties that lead to changes in lectin binding patterns and also enzymatic modification of surface proteins such as sugar transferases. Metabolic changes include increased glycolytic activity and oxygen consumption, hyperactivation associated with activation of adenylate cyclase system and loss of Zn+2 ions that lead to increased nuclear stability (Yanagimachi, 1994; Tulsiani et al., 1997). A number of enzymes and factors that present in the female reproductive tract have been implicated to affect sperm capacitation, such as arylsulphatase, fucosidase and taurine. However, till to date, the exact mechanism on how these enzymes and factors capacitate sperm remains unknown (reviewed in Elder and Dale, 2000).
During capacitation, sperm motility is apparently regulated by changes in the intracellular concentration of calcium ions through a calcium channel (named CatSper) that is only expressed in the tail of mature sperm (Ren et al., 2001). It is thought that the capacitation phenomenon is necessary for spermatozoon to fuse with the oolemma, but not for subsequent steps of fertilization such as DNA decondensation and PN formation. This is supported by in vitro experiments in Xenopus laevis, an amphibian species (Brun, 1974), mouse (Kimura and Yanagimachi, 1995) and human (Palermo et al., 1992) in which injection of intact sperm into the ooplasm resulted in development of normal offspring. However, discrepancies also exists, for example, the capacitation step appears to be critical in cattle (Goto et al., 1990; Sutovsky et al., 1997), goat (Keskintepe et al., 1997; Wang et al., 2003) and in other farm animals where ICSI is problematic (Catt and Rhodes, 1995). The failure of ICSI in some farm animals may be due to the failure of removal of the perinuclear theca, a cytoskeletal capsule present between the sperm membrane and the nuclear envelope (Sutovsky et al., 1997). This structure could block the access of cytoplasmic factors involved in decondensation of the sperm nucleus. This is supported by results where artificially capacitated bull spermatozoa resulted in PN formation and production of a normal calf after ICSI (Goto et al., 1990). Capacitation may be attained in vivo or in vitro. Different in vitro conditions are used to capacitate spermatozoa from various species (Yanagimachi, 1994). Capacitation is temperature-dependant and only occurs at 37° to 39°C.
Acrosome reaction: The first interaction between the spermatozoon and the oocyte is at the ZP, an extracellular coat that surrounds all mammalian oocytes (Wassarman et al., 2001). The sperm nucleus is covered by membrane bound secretory vesicle known as the acrosome which is a large, Golgi-derived lysosome-like organelle that overlies the nucleus in the apical region of the sperm head (Kaji and Kudo, 2004). The acrosome or its surrounding membranes contains a large array of hydrolytic enzymes including hyaluronidase, acrosin, serine protease, proacroson, phosphatase, arylsulphatase, colagenase, phospholipase C and β-galactosidase (reviewed in Elder and Dale, 2000). The spermatozoon interacts with the ZP by the plasma membrane overlying the acrosome. The specific binding of the spermatozoon to the ZP induces in the acrosome a calcium-mediated signal transduction process that leads to AR. The AR brings exposure of the inner acrosomal membrane to the outside by breaking the organelle and involves fusion of the outer acrosomal membrane with the overlying plasma membrane allowing the acrosomal content to be released (Wassarman et al., 2001). This process is necessary for the spermatozoon to acquire fusibility with the oolemma (Kaji and Kudo, 2004). The AR is accompanied by modifications in the sperm plasma membrane, which exposes receptors for ZP binding and possibly factors exposed on the equatorial segment in preparation for sperm-oocyte fusion (Kupker et al., 1998). The acrosomal enzymes react with the extracellular matrix (or ZP) of oocyte and digest the ZP at the point of spermatozoon contact. After losing its acrosomal content the spermatozoon retains only the inner acrosomal membrane that makes direct contact with the ZP during penetration. Following fusion, the sperm plasma membrane remains in the ooplasm that indicate the point of fusion.
In mammals, Austin and Bishop (Austin and Bishop, 1958) first described the AR. Although, AR is now considered as a separate process (Bavister, 2002), which triggers after capacitation, however, discrepancies exists as some authors believe that it was a part of the capacitation process (Austin and Bishop, 1958) and others considered AR as the final phase of capacitation (Yanagimachi, 1969). The AR is considered as the final prerequisite step in the sperm activation process before it gets the ability to fuse with the oocyte. It is obligatory that this reaction only occur in the presence of a rise in the intracellular calcium (Ca2+) level (Florman and Babcock, 1991; Yanagimachi, 1994; Ben-Yosef and Shalgi, 1998; Wassarman, 1999). This increase of intracellular calcium can be achieved in vitro by exposing sperm to Ca2+ ionophores or phosphodiesterase inhibitors (Kupker et al., 1998). An artificially high pH of 9.0-9.5 could also induce AR (Elder and Dale, 2000). Although, it is not clear whether the AR is initiated whilst the sperm is interacting with the cumulus mass, however, it is speculated that the reaction may be started in the CCs as a major component of the cumulus matrix is hyaluronic acid and the acrosome contains hyaluronidase. The AR is relatively rapid once the correct trigger signals have been received and may take 2 to 15 min in vitro (Elder and Dale, 2000).
Sperm-oocyte interaction: As mentioned earlier, sperm-oocyte interaction is the earliest step towards the fertilization process, which involves the contact between the acrosome of the sperm and the ZP of the oocyte. The acrosome is membrane-bound and contains lytic agents such as proteases, sulphatases and glycosidases. On the other hand, the ZP of the oocyte is made up of protein and carbohydrates in the form of glycoprotein units that are probably stabilized by disulphide bonds. The principal types of carbohydrate found are fucose and glycoprotein units, which are synthesized by the oocyte itself (reviewed in Elder and Dale, 2000).
After completion of AR, the sperm penetrate the ZP. Following penetration, the acrosome-reacted sperm passes through PVS and interacts with the sperm plasma membrane of the oocyte (oolemma) (Schultz and Kopf, 1995). This initial interaction involves specific recognition, binding and then fusion between the sperm plasma membrane in the region of the equatorial segment and the oolemma (Yanagimachi, 1994). The fusion site on the equatorial segment may be excluded from the acrosome reaction and the sperm plasma membrane is still intact on the apical part of the postacrosomal sheath and the equatorial segment. The sperm tail is still beating outside the ZP after its head and mid-piece have entered the PVS and the equatorial segment has made an initial contact with the oolemma. The sperm head fuses with microvilli on the oolemma (Kupker et al., 1998). The attachment of the sperm head to the ZP inevitably alters the permeability of the sperm plasma membrane, causing a transient change in the concentration of several intracellular ions (Tulsiani et al., 1997). Once fusion has occurred, the microvilli retract and draw most or the entire spermatozoon into the ooplasm. As it is incorporated into oocyte, the spermatozoon becomes immotile. Subsequently the entire spermatozoon including the tail is drawn into the ooplasm (Kupker et al., 1998). This initiates syngamy and further development of the zygote (Tulsiani et al., 1997). The exact mechanism of sperm penetration of the ZP and the complementary molecules that initiate penetration of the ZP by the sperm are not yet known. However, it is generally believed that the process involves receptor-ligand interaction between sperm-surface proteins and ZP glycoproteins. In rodents, these consist of sperm membrane galactosyltransferase and N-acetylglucosamine residues of the ZP. The receptor-ligand interaction induces the AR leading to exocytosis and release of acrosomal enzymes from the acrosomal cap. The specific sperm protein(s) responsible for sperm-oocyte interaction are yet to be fully characterized. However, four ZP proteins with variable roles in sperm-oocyte interaction have been isolated in mammals. In the mouse, ZP3 plays the primary role, while ZP1 is the primary sperm receptor in pig, rabbit and non-human primates (Dunbar et al., 1998). It is not known yet whether such a characterization is available for the goat. It is thought that the penetration of the ZP by the sperm is mediated by lytic acrosomal enzymes (Tulsiani et al., 1997; Bedford, 1998) or by the mechanical action of the sperm itself (Tulsiani et al., 1997; Bedford, 1998). The latter author argues that available evidence is in favor of mechanical instead of lytic penetration.
FERTILIZATION AND EARLY EMBRYOGENESIS
Fertilization in mammals is one of the most carefully regulated cell-cell interactions in the animal body, involving two morphologically disparate cells that must recognize, bind and fuse with each other in a very specific way. It can be defined as the process of union of two germ cells, namely spermatozoon and oocyte, whereby the somatic chromosome number is restored and the development of a new and unique individual exhibiting characteristic of the same species is initiated (Wassarman, 1999). By transferring genetic information from one generation to the next, it ensures the immortality of an individual and by creating variation it allows evolutionary forces to operate (Elder and Dale, 2000).
As already mentioned earlier, before penetrating into the oocyte, various biochemical changes occur in the spermatozoon including capacitation, AR and sperm oocyte interaction. Following ZP penetration, a series of events take place leading to syngamy and the production of the zygote. These steps have been summarized previously (Bedford, 1982; Yanagimachi, 1994). In the following three sub-sections oocyte activation, PN formation and cleavage as well as early embryonic development or embryogenesis in mammals with special reference to goat is briefly discussed.
Activation of oocyte: Oocyte activation is the restoration of metabolic activity in the quiescent oocyte, in other word; it is the process of releasing the oocyte from the second meiotic arrest when the spermatozoon fertilizes it. It is a cell signaling event that results in events including the cortical granule reaction or Cortical Reaction (CR), decondensation of the sperm nucleus, maternal RNA recruitment, resumption of meiosis as evidenced by the extrusion of second polar body (PB-2) and later events such as PN formation and initiation of DNA synthesis and cleavage (Yanagimachi, 1994). The very early cellular event observed in all activated mammalian oocytes is an intracellular rise in Ca2+ concentration. In human, this increase occurs within 1 to 3 min of sperm-oocyte fusion and takes the form of a wave originating at the point of spermatozoon entry (Reviewed in Ben-Yosef and Shalgi, 2001). The site of Ca2+ release and sequestration is thought to be the endoplasmic reticulum, where inositol 1, 4, 5-triphosphate (IP3) receptors are present (Kline and Kline, 1992). The first Ca2+ transient is followed by a series of shorter Ca2+ transients of high amplitude (Ca2+ oscillations). In IVF studies, Ca2+ oscillations were observed in all mammalian species including goat (Jellerette et al., 2006), although their frequency is species specific (Ben-Yosef and Shalgi, 2001). As fertilization progresses, the amplitude and frequency of the Ca2+ transients decrease, while the duration increases until an absolute cessation of Ca2+ oscillations during entry into interphase and PN formation, several hours after sperm entry (Jones et al., 1995). Calcium oscillations require a continuous Ca2+ influx to refill endoplasmic reticulum stores (Miyazaki, 1995). It was suggested that although a single rise is sufficient to produce activation, oscillations might be required for additional developmental events (Ozil, 1990).
There are two postulated hypotheses of oocyte activation mechanism, namely ‘receptor hypothesis` and sperm factor hypothesis, which describe the contribution of the spermatozoa to successful oocyte activation (Kimura et al., 1998; Fissore et al., 1998 ). In the receptor hypothesis, it is the spermatozoon interacting with the oolemma that results in oocyte activation. In this model, an oocyte surface receptor is coupled to a G-protein (Miyazaki et al., 1993) or a tyrosine-kinase-mediated (Ben-Yosef and Shalgi, 1998) signaling pathway and when activated by a sperm, activates phospholipase C that results in the formation of IP3 and hence intracellular Ca2+ is released. The second hypothesis, which is called the sperm factor hypothesis, suggests that the release of intracellular Ca2+ is triggered by diffusible messenger(s) in sperm cytoplasm which enters the ooplasm following sperm-oocyte fusion. This hypothesis gained support with the development of ICSI technique where initial Ca2+ rise, Ca2+ oscillations and full oocyte activation occur after injecting sperm or sperm extract into the ooplasm (Fissore et al., 1998).
Formation of pronuclei: Soon after oocyte activation through sperm entry the PB-2 extrudes (Payne et al., 1997). The PB-2 is generally extrudes immediately adjacent to the PB-1. The mammalian sperm nucleus is packed with distinct protamines. When sperm enters the ooplasm, the sperm nuclear envelope breaks down, the protamines are lost, the sperm head enlarges in the ooplasm and chromosome or nuclear decondensation takes place a few hours by reduction of disulphide bonds between protamines by the action of glutathione (Kupker et al., 1998). Protamines are replaced by the histones generated by the oocyte. Formation of MPN takes place simultaneously with disappearance of the nuclear membrane, decondensation of the chromosomes and reformation of the pronuclear membrane that is supported by action of growth factors. This coincides with decondensation of maternal chromatin and formation of the female PN (FPN). Usually, but not always, the MPN appears near the site of sperm entry whereas the FPN forms close to the PB-2 (Payne et al., 1997). At this stage (pronuclear development), DNA synthesis and RNA transcription begin. The MPN and FPN then merge (syngamy) to form the zygote. This marks the end of the fertilization process and the beginning of embryonic development.
Cleavage and early embryogenesis: After the zygote stage, embryo enters into several mitotic divisions. The zygote or one-cell stage embryo is comparatively large, having a low nuclear to ooplasmic ratio. DNA replication and cell divisions occur without an increase in cell mass to attain a ratio similar to somatic cells. This process is referred to as cleavage (Hafez and Hafez, 2000b). Once the diploid constitution of the species has been restored by fertilization then the cleavage commences. Cleavage of the zygote occurs by vertical division through the main axis of the oocyte from the animal to vegetal pole (animal pole refers to the site of PB extrusion and vegetal pole refers to the site of yolk reserve). The cleavage furrow often goes through the region where the PN resided at the initiation of syngamy. The resulting daughter cells are known as blastomeres. The plane of second division is also vertical and passes through the main axis but at a right angle to the initial plane of cleavage, resulting in 4 blastomeres. The third cleavage division occurs approximately at a right angle to the second, resulting in 8 blastomeres. This doubling sequence is followed on through the remainder of early cleavage (Hafez and Hafez, 2000b). Cleavage divisions are always mitotic and each blastomere receives full assortment of chromosomes. Although the blastomeres undergo mitosis like adult somatic cells and their number increases, however, unlike somatic cells they do not retain the size of the parent cell. The blastomeres resulting from each division are always approximately half the size of the parent blastomere. Thus, the size of a pre-implantation embryo remains the same even though the number of blastomeres increases. The embryo undergoes polarization and differences arise between the blastomeres as cleavage stage progresses. This may be due to the unequal distribution of ooplasmic components already laid down in the oocyte during oogenesis, or due to new embryonic gene transcription that results in changes to blastomeres. This way, each blastomere sets off on its own particular program of development to give rise to specific cell lines, for example, epithelium, muscle, nerve or other connective tissue. During cleavage stage, individual blastomeres can be clearly detected when viewed under the microscope. It is important to note that the precise cleavage of zygotes and blastomeres into two equally sized daughter cells relies upon the position of the spindle and the functional activity of cytoskeletal elements. Slight variations in blastomere sizes within the same embryo are probably unimportant, but major differences may indicate defects in underlying cellular processes (Hardarson et al., 2001). Although oocyte size (diameter of ooplasm) was measured in most of the species, there was paucity of information on blastomeres size. A search of available literature revealed one paper where morphometric measurements of human ICSI-derived cleaved embryos were performed up to 4-cell stage. Using computer-controlled morphometric analysis of blastomere size, it was found that average diameter of blastomeres of 2-, 3-and 4-cell stage embryos were 80.1, 68.7 and 64.9 μm, respectively (Hnida et al., 2004). The diameter of ooplasm of a fully grown human oocyte or 1-cell embryo is 135 to 160 μm (Oppenheimer and Lefevre, 1989) which is about the size of a mature goat oocyte (119-146 μm) (Crozet et al., 2000) (Table 1). It was found from their study that blastomere size significantly affected by degree of fragmentation and multinuclearity and that computer-assisted, multilevel analysis of blastomere size may function as a biomarker for embryo quality (Hnida et al., 2004). The chronology of embryo development from 2-cell stage to blastocyst hatching in different species of farm animals including goat, rabbit, mouse and human has been presented in Table 3. In goat, initial and principal activations of embryonic genome (cell stage) are reported to be occurring at 2-cell and 8-cell stage, respectively (Kelk et al., 1994). After the embryo reaches to the morula stage it is very difficult to discern the boundaries between blastomeres as the embryo undergoes compaction in which the blastomeres flatten against each other to form intercellular junctions between them. Depending upon the species, there is a great deal of variation of the beginning of embryo compaction stage. In goat, compaction of the embryo begins at 8-cell stage (Sakkas et al., 1989). The data for other farm animals, rabbit, mouse and human are depicted in Table 3. As the morula progresses to further embryonic development, some morphological change occurs and the morula progresses to blastocyst stage. The blastocyst is characterized by the formation of an outer layer of trophectoderm cells, Inner Cell Mass (ICM) and a fluid-filled cavity, known as blastocoel. In goat, the blastocoel appears at 120 h post-fertilization (Sakkas et al., 1989). Data for other farm animals, rabbit, mouse and human have been depicted in Table 3.
| Table 3: | Chronology of pre-implantation embryo development in goat and other farm animals, rabbit, mouse and human |
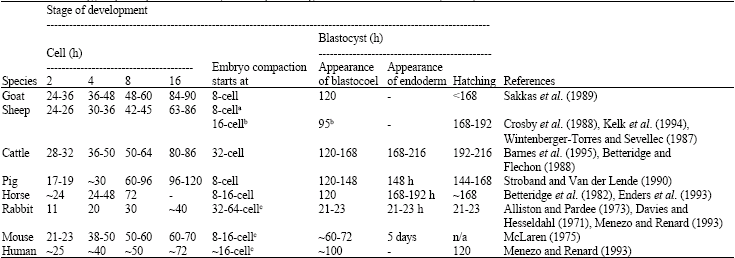 | |
| N.B: Data mainly derived from in vivo studies; h = hours; n/a = not applicable; aKelk et al. (1994); bCrosby et al. (1988); cYang (1991) | |
The embryo is confined within the ZP (analogous to eggshell in bird, reptiles or monotremes) throughout the early stages of development. This keeps the blastomeres together prior to compaction and prevents embryos from fusing together. During early cleavage, there is little increase in metabolic rate, however, a sharp rise occurs between the morula and blastocyst stage. The goat embryo stays in the oviduct for 48 to 72 h from the time of ovulation (Moore, 1977). The trophectoderm cells of the embryo secrete proteolytic enzymes that digest a passage through the ZP and allow the blastocyst to hatch. The hatching of goat blastocyst occurs around 168 h (7 days) after fertilization (Sakkas et al., 1989). The times required to blastocyst hatching in case of other domestic animals, rabbit, mouse and human are depicted in Table 3. The exposed cells of the goat blastocyst then make firm physical contact with the uterine wall, implantation stage begins, embryo becomes elongated and the final implantation in the uterine endometrium occurs at day 16 post-fertilization (Gurdon et al., 1995).
CONCLUSIONS
Currently, goat producing developed countries focusing their researches on ARTs for rapid increment of goat numbers as well as genetic improvements. Besides, goats are found to be more appropriate animal for the production of valuable recombinant pharmaceutical or biomedical protein through using latest ARTs like transgenesis, NT or ICSI. Although, using modern ARTs genetic and overall improvement in goat have been achieved in some extent, however, it is still a long way to go. To get success in any in vitro studies in any species of animals it crucial to know the normal fertilization process that occurs in vivo. Although, previously a number of in vivo fertilization studies have been performed in goat, however, compare to cattle, sheep and laboratory animals it is still not enough. Most of the description of goat gametogenesis, sperm-oocyte interaction, fertilization and early embryogenesis are mainly based on the data derived from sheep and in some extent cattle. Although, sheep and cattle are also ruminants, however, it is to be remembered that they are different species and their physiology is different from that of goat. Therefore, there is still a need for study some basic in vivo aspects of gametogenesis, fertilization and pre-implantation embryogentic process in goat which will not only fill up gaps of information but also help in the successful deigning of suitable ARTs for rapid improvement of goat industry.
ACKNOWLEDGMENTS
The authors wish to thank Islamic Development Bank (IDB) for providing an IDB Merit Scholarship to the first author. This work was supported by grants from MOSTI Special Project (Grant No. 01-02-03-0696) and IPPP (Grant No. Vote F-0179/2004D, 0145/2005D and P0170/2006C).
REFERENCES
- Adams, C.E., 1958. Egg development in the rabbit: the influence of post-coital ligation of the uterine tube and of ovariectomy. J. Endocrinol., 16: 283-293.
PubMed - Alliston, C.W. and N.R. Pardee, 1973. Variability of embryonic development in the rabbit at 19 to 168 hours after mating. Lab. Anim. Sci., 23: 665-670.
PubMed - Ariyaratna, H.B.S. and V.K. Gunawardana, 1997. Morphology and morphometry of ovarian follicles in the goat. Small Rum. Res., 26: 123-129.
CrossRef - Austin, C.R. and M.W.H. Bishop, 1958. Capacitation of mammalian spermatozoa. Nature., 181: 851-851.
PubMed - Barnes, F.L., A. Crombie, D.K. Gardner, A. Kausche, O. Lacham-Kaplan and A.M. Suikkari et al., 1995. Blastocyst development and birth after in-vitro maturation of human primary oocytes, intracytoplasmic sperm injection and assisted hatching. Hum. Reprod., 10: 3243-3247.
PubMedDirect Link - Barth, A.D. and R.J. Oko, 1989. Abnomal Morphology of Bovine Spermatozoa. 1st Edn., Iowa State Univ. Press, Ames, Iowa, USA. ISBN: 0813801125.
Direct Link - Bavister, B.D., 2002. Early history of In vitro fertilization. Reproduction, 124: 181-196.
Direct Link - Bedford, J.M., 1998. Mammalian fertilisation misread?: Sperm penetration of the eutherian zona pellucida is unlikely to be a lytic event. Biol. Reprod., 59: 1275-1287.
Direct Link - Ben-Yosef, D. and R. Shalgi, 1998. Early ionic events in activation of the mammalian eggs. Rev. Reprod., 3: 96-103.
Direct Link - Ben-Yosef, D. and R. Shalgi, 2001. Oocyte activation: lessons from human infertility. Trends Mol. Med., 7: 163-169.
CrossRef - Betteridge, K.J., 1995. Phylogeny, ontogeny and embryo transfer. Theriogenology, 44: 1061-1098.
CrossRef - Betteridge, K.J., 2003. A history of farm animal embryo transfer and some associated techniques. Anim. Reprod. Sci., 79: 203-244.
CrossRefDirect Link - Betteridge, K.J., M.D. Eaglesome, D. Mitchell, P.F. Flood and R. Beriault, 1982. Development of horse embryos up to twenty-two days after ovulation: Observations on fresh specimens. J. Anat., 135: 191-209.
PubMedDirect Link - Betteridge, K.J. and J. Flechon, 1988. Anatomy and physiology of pre-attachment bovine embryos. Theriogenology, 29: 155-187.
CrossRef - Buccione, R., A.C. Schroeder and J.J. Eppig, 1990. Interactions between somatic cells and germ cells throughtout mammalian oogenesis. Biol. Reprod., 43: 543-547.
PubMedDirect Link - Catt, J.W. and S.L. Rhodes, 1995. Comparative intracytoplasmic sperm injection (ICSI) in human and domestic species. Reprod. Fertil. Dev., 7: 161-166.
CrossRef - Chang, M.C., 1951. Fertilizing capacity of spermatozoa deposited into the fallopian tubes. Nature, 168: 697-698.
CrossRefPubMedDirect Link - Crosby, I.M., F. Gandolfi and R.M. Moor, 1988. Control of protein synthesis during early cleavage of sheep embryos. J. Reprod. Fertil., 82: 769-775.
PubMed - Crozet, N., M. Ahmed-Ali and M.P. Dubos, 1995. Developmental competence of goat oocytes from follicles of different size categories following maturation, fertilization and culture in vitro. J. Reprod. Fertil., 103: 293-298.
Direct Link - Crozet, N., M. Dahirel and L. Gall, 2000. Meiotic competence of in vitro grown goat oocytes. J. Reprod. Fert., 118: 367-373.
PubMedDirect Link - Crozet, N., V. De Smedt, M. Ahmed-Ali and C. Sevelec, 1993. Normal development following in vitro oocyte maturation and fertilization in the goat. Theriogenology, 39: 206-206.
CrossRef - Dalvit, G.C., M.H. Mingaya, M.G. Chaves and M.T. Beconi, 1995. Energy requirement of bovine spermatozoa for in vitro capacitation. Theriogenology, 44: 1051-1058.
CrossRef - De Kretser, D.M., K.L. Loveland, A. Meinhardt, D. Simorangkir and N. Wreford, 1998. Spermatogenesis. Hum. Reprod., 13: 1-8.
CrossRef - De Loos, F., P. Kastrop, P. Van Maurik, T.H. Van Beneden and T.A.M. Kruip, 1991. Heterologous cell contacts and metabolic coupling in bovine cumulus oocyte complexes. Mol. Reprod. Dev., 28: 255-259.
PubMed - De Smedt, V., N. Crozet and L. Gall, 1994. Morphological and functional changes accompanying the acquisition of meiotic competence in ovarian goat oocyte. J. Exp. Zool., 269: 128-139.
PubMed - Enders, A.C., S. Schlafke, K.C. Lantz and I.K.M. Liu, 1993. Endoderm cells of the equine yolk sac from Day 7 until formation of the definitive yolk sac placenta. Equine Vet. J., 15: 3-9.
CrossRef - Eppig, J.J. and S.M. Downs, 1984. Chemical signals that regulate mammalian oocyte maturation. Biol. Reprod., 30: 1-11.
Direct Link - Fair, T., 2003. Follicular oocyte growth and acquisition of developmental competence. Anim. Reprod. Sci., 78: 203-216.
CrossRef - Fissore, U.A., A.C. Gordo and H. Wu, 1998. Activation of development in mammals: Is there a role for a sperm cytosolic factor? Theriogenology, 49: 43-52.
PubMed - Gosden, R., J. Krapez and D. Briggs, 1997. Growth and development of the mammalian oocyte. BioEssays, 19: 875-882.
PubMed - Goto, K., Y. Kinoshita, Y. Takuma and K. Ogawa, 1990. Fertilisation of bovine oocytes by the injection of immobilised, killed spermatozoa. Vet. Rec., 127: 517-520.
PubMed - Gravance, C.G., K.M. Lewis and P.J. Cassey, 1995. Computer automated sperm-head morphometry analysis (ASMA) of goat spermatozoa. Theriogenology, 44: 989-1002.
CrossRef - Gurdon, J.B., A. Mitchell and D. Mahony, 1995. Direct and continuous assessment by cells of their position in a morphogen gradient. Nature, 376: 520-521.
CrossRef - Hardarson, T., C. Hanson, A. Sjögren and K. Lundin, 2001. Human embryos with unevenly sized blastomeres have lower pregnancy and implantation rates: Indications for aneuploidy and multinucleation. Hum. Reprod., 16: 313-318.
PubMed - Harrison, R.A.P., 1996. Capacitation mechanisms and the role of capacitation as seen in eutherian mammals. Reprod. Fertil. Dev., 8: 581-594.
CrossRef - Hnida, C., E. Engenheiro and S. Ziebe, 2004. Computer-controlled, multilevel, morphometric analysis of blastomere size as biomarker of fragmentation and multinuclearity in human embryos. Hum. Reprod., 19: 288-293.
Direct Link - Hyttel, P., T. Fair, H. Callesen and T. Greve, 1997. Oocyte growth, capacitation and final maturation in cattle. Theriogenology, 47: 23-32.
CrossRefDirect Link - Jellerette, T., D. Melican, R.E. Butler, S. Nims, C. Ziomek, R. Fissore et al., 2006. Characterisation of calcium oscillation pattern in caprine oocytes induced by IVF or an activation techniques used in nuclear transfer. Theriogenology, 65: 1575-1586.
CrossRef - Jones, K.T., J. Caroll, J.A. Merriman, D.G. Whittingham and T. Kono, 1995. Repetitive sperm-induced Ca2+ transients in mouse oocytes are cell cycle dependant. Development, 121: 3259-3266.
PubMed - Jones, R.C. and R.N. Murdoch, 1996. Regulation of the motility and metabolism of spermatozoa for storage in the epididymis of eutherian and marsupial animals. Reprod. Fertil. Dev., 8: 553-568.
CrossRef - Kaji, K. and A. Kudo, 2004. The mechanism of sperm-oocyte fusion in mammals. Reproduction, 127: 423-429.
target='_blank' class='btn btn-sm btn-outline-primary mr-3 mt-3'>Direct Link - Keskintepe, L., P. Morton, S.E. Smith, M.J. Tucker, A.A. Simplicio and B.G. Brackett, 1997. Caprine blastocyst formation following intracytoplasmic sperm injection and defined culture. Zygote, 5: 261-265.
PubMed - Kimura, Y. and R. Yanagimachi, 1995. Intracytoplasmic sperm injection in the mouse. Biol. Reprod., 52: 709-720.
Direct Link - Kimura, Y., R. Yanagimachi, S. Kuretake, H. Bortkiewicz, A.C.F. Perry and H. Yanagimachi, 1998. Analysis of mouse oocyte activation suggests the involvement of sperm perinuclear material. Biol. Reprod., 58: 1407-1415.
Direct Link - Kline, D. and J.T. Kline, 1992. Repetitive calcium transients and the role of calcium in exocytosis and cell cycle activation in the mouse eggs. Dev. Biol., 149: 80-89.
CrossRef - Kupker, W., K. Diedrich and R.G. Edwards, 1998. Principles of mammalian fertilization. Hum. Reprod., 13: 20-31.
Direct Link - Lonergan, P., P. Monagham, D. Rizos, M.P. Boland and L. Gordon, 1994. Effect of follicle size on bovine oocyte quality and developmental competence following maturation, fertilization and culture in vitro. Mol. Reprod. Dev., 37: 48-53.
CrossRefPubMedDirect Link - Markert, C.L., 1983. Fertilisation of mammalian eggs by sperm injection. J. Exp. Zool., 228: 195-201.
PubMed - McNatty, K.P., P. Smith, N.L. Hudson, D.A. Heath, D.J. Tisdall, O. Ws and R. Braw-Tal, 1995. Development of the sheep ovary during fetal and early neonatal life and the effect of fecundity genes. J. Reprod. Fertil., 49: 123-135.
PubMed - Miyano, T., 2003. Bringing up small oocytes to eggs in pigs and cows. Theriogenology, 59: 61-72.
CrossRef - Miyano, T. and Y. Hirao, 2003. In vitro growth of oocytes from domestic species. J. Mammal. Ova Res., 20: 78-85.
Direct Link - Miyazaki, S., 1995. Inositol triphosphate receptor mediated spatiotemporal calicum signalling. Current Opin. Cell Biol., 7: 190-196.
CrossRef - Miyazaki, S., M. Shirakawa, K. Nakada and Y. Honda, 1993. Essential role of the inositol 1,4,5 trisphasphate receptor/Ca2+ release channel in Ca2+ waves and Ca2+ oscillations at fertilization of mammalian eggs. Dev. Biol., 158: 62-78.
CrossRef - Olszanska, B. and A. Borgul, 1993. Maternal RNA content in oocytes of several marnmalian and avian species. J. Exp. Zool., 265: 317-320.
PubMed - Ozil, J.P., 1990. The parthenogenetic development of rabbit oocytes after repetitive pulsatile electrical stimulation. Development, 109: 117-127.
PubMed - Payne, D., S.P. Flaherty, M.F. Barry and C.D. Matthews, 1997. Preleminary observation on polar body extrusion and pronuclear formation in the human oocytes using time-lapse video cinematography. Hum. Reprod., 12: 532-541.
PubMed - Ren, D., B. Navarro, G. Perez, A.C. Jackson, S. Hsu and Q. Shi et al., 2001. A sperm ion channel required for sperm motility and male fertility. Nature, 413: 603-609.
CrossRefDirect Link - Sakkas, D., P.A. Batt and A.W. Cameron, 1989. Development of preimplantation goat (Capra hircus) embryos in vivo and in vitro. Reprod. Fertil., 87: 359-365.
PubMed - Schultz, R.M. and G.S. Kopf, 1995. Molecular basis of mammalian egg activation. Current Topics Dev. Biol., 30: 21-61.
CrossRef - Stroband, H.W.J. and T. Van der Lende, 1990. Embryonic and uterine development during early pregnancy in pigs. J. Reprod. Fertil. Suppl., 40: 261-277.
PubMed - Sutovsky, P., R. Oko, L. Hewitson and G. Schatten, 1997. The removal of sperm perinuclear theca and its association with the bovine oocyte surface during fertilization. Dev. Biol., 188: 75-84.
CrossRef - Tsafriri, A., N. Dekel and S. Bar-Ami, 1982. The role of oocyte maturation inhibition in follicular regulation of oocyte maturation. J. Reprod. Fertil., 64: 541-551.
PubMed - Tulsiani, D.R.P., H. Yoshida-Komiya and Y. Araki, 1997. Mammalian fertilization: A carbohydrate-mediated event. Biol. Reprod., 57: 487-494.
Direct Link - Van den Hurk, R. and J. Zhao, 2005. Formation of mammalian oocytes and their growth, differentiation and maturation within ovarian follicles. Theriogenology, 63: 1717-1751.
CrossRef - Wang, B., H. Baldassarre, J. Pierson, F. Cote, K.M. Rao and C.N. Karatzas, 2003. The in vivo and in vitro development of goat embryos produced by intracytoplasmic sperm injection using tail-cut spermatozoa. Zygote, 11: 219-227.
PubMed - Wassarman, P.M., 1999. Mammalian fertilization: Molecular aspects of gamete adhesion, exocytosis and fusion. Cell, 96: 175-183.
PubMed - Wassarman, P.M., L. Jovine and E.S. Litscher, 2001. A profile of fertilization in mammals. Nat. Cell Biol., 3: E59-E64.
Direct Link - Yanagimachi, R., 1969. In vitrocapacitation of hamster spermatozoa by follicular fluid. J. Reprod. Fertil., 18: 275-286.
PubMed - Zhou, H.M. and Y. Zhang, 2006. In vitro growth and maturation of caprine oocytes. Reprod. Domest. Anim., 41: 444-447.
PubMed
Simoen Dip Reply
This review article appears to be valuable for providing an overview of gametogenesis, fertilization, and early embryogenesis in mammals, with a special focus on goats. This review could be a useful resource for researchers and practitioners working in the field of animal reproduction.
Editor
Thank you for your positive feedback on the review article "Gametogenesis, Fertilization and Early Embryogenesis in Mammals with Special Reference to Goat: A Review." We are glad to hear that the article was useful and informative for you. We hope that this review will contribute to the advancement of knowledge and research in the field of animal reproduction, particularly with respect to goats. Please feel free to share any suggestions or feedback that you may have.