
Hussein M. Al-Wadi
Department of Biological Sciences, College of Sciences,
King Khalid University, Abha, 9004, Saudi Arabia
Gamal M.A. Lashin
Department of Biological Sciences, College of Sciences,
King Khalid University, Abha, 9004, Saudi Arabia
Journal of Biological Sciences
Year: 2007 | Volume: 7 | Issue: 4 | Page No.: 626-631







ABSTRACT
This study aims to explore the relationship between cytology and palynology relationship between the species under consideration. Pollen morphology and cytology of three species (Solanum forsskalii, S. macracanthum and S. glabrata var. sepicula) of genus Solanum of the family Solanaceae from Saudi Arabia has been investigated by using Light Microscope (LM) and electron microscope (SEM and TEM). Pollen grains are generally radially symmetrical, isopolar, tricolporates, zonoaperturates, prolate and with non perforate tectum. Tecta scabrate often punctuate. The dimensions of the pollen of the three species vary according to polar and equatorial axis. On the other hand according to pollen shape, two distinct pollen types are recognized, prolate-subprolate for Solanum forsskalii and S. macracanthum and prolate-spheroidal for S. glabrata var. sepicula. In addition, on the basis of exine columella, three types are recognized, viz., S. glabrata var. sepicula (regularly columellate), S. forsskalii (less and irregular columellate) and Solanum macracanthum (columella indistinct). It is obviously that solanaceae is considered an europalynous family. Chromosomal characters of the three species provides that Solanum forsskalii and S. glabrata var. sepicula are cytologically stable and higher pollen fertility (80-90%) while Solanum macracanthum shows irregular meiosis and less pollen fertility (35%). The Pollen fertility and cross-pollination between the studdied species were also discussed. These data could furnish adequate information that support the relation characters between the species.
PDF Abstract XML References Citation
How to cite this article
Hussein M. Al-Wadi and Gamal M.A. Lashin, 2007. Palynological and Cytological Characters of Three Species of Genus Solanum (Family: Solanaceae) from Saudi Arabia. Journal of Biological Sciences, 7: 626-631.
DOI: 10.3923/jbs.2007.626.631
URL: https://scialert.net/abstract/?doi=jbs.2007.626.631
DOI: 10.3923/jbs.2007.626.631
URL: https://scialert.net/abstract/?doi=jbs.2007.626.631
INTRODUCTION
Solanaceae is considered as an important and large family between angiosperm and distributed throughout tropical and temperate regions of the world. Its center of diversity is western and southern South America and only ten genera are native to new and old world, while Solanum, Physalis and Lycium are widely distributed across both the eastern and western hemisphere (D’Arcy, 1991). This family is composed of 98 genera and about 2300 species belonging to 14 tribes grouped in three subfamilies (Hunziker, 1979; D’Arcy, 1991). Judd et al. (1999) reported that the number of the genera in the Solanaceae sense lato is about 147 genera occurring worldwide of which 7 genera and 33 species occur in Saudi Arabia (Collenette, 1998). Several studies have been carried out on the generic and sub-generic level (Sharma, 1974; Mabberley, 1997). Solanum is an economic important genus of annuals, perennials, sub-shrubs, shrubs and climbers. Most of its species are poisonous and medically important. They often have attractive fruit and flowers and bear edible fruit or other parts, such as the common foods tomato, potato, eggplant and pepper. Synonyms and common name: Nightshade, as well as referring to the related genus Atropa, is also used for many members of the genus Solanum.
Pollen characters have received attention in taxonomic and pollen morphology but still very little is known about the ultra-structure of the pollen wall stratification and internal structure can hardly be studied by light microscope (Zavada, 1990). Therefore the scanning and transmission electron microscopy become necessary in examining these characters. Many studies were reported on pollen morphology of plant species (Punt and Nienhuis, 1977; Hesse, 1981; El-Ghazaly, 1990; Harley et al., 2000). So far, no previous ultra-structural studies of pollen grains of family Solanaceae have been reported in Saudi Arabia except the study of Al-Wadi (2000), who studied the pollen morphology and ultra-structure of Withania sommnifera. In Arabic region, El-Ghazaly (1993, 1999) studied some species of Solanaceae from Sudan and Qatar. Many species of this genus were subjected to cytological studies in the world (Anis et al., 1994). Haroun and Al-Wadi (1999) have studied meiotic division and breeding of three wild species of Solanum in Saudi Arabia. Palynological and cytological analysis with a combination of different parameter is believed to serve in a better understanding and evaluation of the relationships between taxa of the genus Solanum. However the present study aims to investigate palynological and cytological characters of three species of Solanum widely distributed in south-western part of Saudi Arabia.
MATERIALS AND METHODS
The studied materials were obtained from the herbarium specimens deposited at the Department of Biological Sciences (1992-2006), Collage of Sciences, King Khalid University, Abha, Saudi Arabia (Table 1).
Processing of pollen grains for light microscope (LM): Polliniferous materials were placed in glacial acetic acid for three minutes, acetolysed according to the method of Erdtman (1960) and Moore et al. (1991) and then mounted in glycerin jelly.
| Table 1: | Sites of collection of the examined taxa of genus Solanum. The taxa were identified according to Migahid (1978) and Collenette (1985, 1998) |
 | |
The investigation and photographing were done by a CH 2 Olympus light microscope.
Processing of pollen grains for scanning electron microscope (SEM): The dried polliniferous material was attached to stubs with double-faced selotape. The stubs were gold-coated in B sputter coater for one minute and examined (Moore et al., 1991). The representative pollen grains were photographed at various magnifications in a Jeol-T. 100 SEM operated at an accelerating voltage of 15 kv.
Processing of pollen grains for transmission electron microscope (TEM): The polliniferous materials were fixed in 2.5% glutaraldehyde in 0.05M cacoldylate buffer at pH 7.4 for 24 h. and post fixed in 1% OsO 4 in the same buffer for 2 h (Cresti et al., 1985). The pollen then dehydrated in graded series of ethanol and embedded in Spurr’ s resin (Spurr, 1969). Ultra thin sections were cut using a diamond knife on Ultra microtome, stained with uranyl acetate followed by lead citrate (Reynolds, 1963). The stained grids were examined and photographed with a Jeol-T. 100 B TEM.
RESULTS
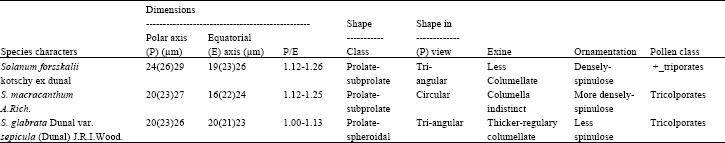
The palynological characters are illustrated in Table 2 and Fig. 1-3. Pollen grains are generally radially symmetrical, isopolar, tricolporates, zonoaperturates and prolate with non-perforate tecta. Tecta are scabrate or often punctuate.
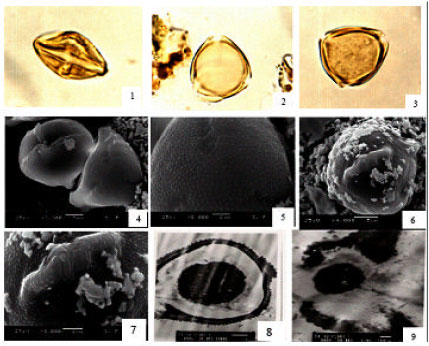 | |
| Fig. 1: | Solanum forsskalii 1: LM equatorial view, showing tricolporate pollen.x1000. 2: LM, polar view showing thick exine and micro-ornamentation. x1000.3: LM polar view showing pores structure. x1000. 4: SEM, Equatotrial view and polar view showing triporates pollen and tricolpate furrow. 5: SEM, polar view, showing surfaced furrow. 6,7: SEM, equatorial view showing inaperturate lalongate pore. 8: TEM, LS, in pollen grain, showing spinulose tecta,. x4000. 9: TEM, LS, in pollen grains showing circular lalongate pore and fastigium-like cavity formed by ectoapertural margins. x5000 |
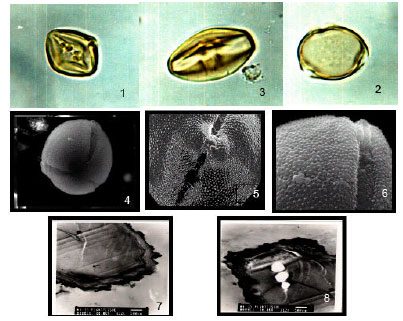 | |
| Fig. 2: | Solanum forsskalii 1: LM, LS., equatorial view pollen grains 2: LM. polar view, showing protrusion of ectoapretures and microechnates exine. x1000. showing large colpi.x1000. 3: LM, equatorial view showing tricolporate pollen grains and thick exine. x1000. 4: SEM, polar view of pollen grain showing tricolpates furrows x3500. 5: SEM, equatorial view, note ectoaperture protrusion and lalongate pore.x7000. 6: SEM, equatorial view showing spinulose exine and granular colpus. x10000. 7-8: TEM, LS, in pollen grain, showing irregular exine |
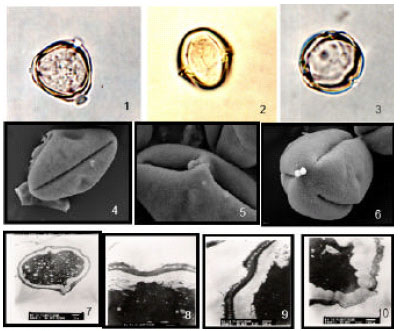 | |
| Fig. 3: | Solanum glabrata var. sepicula, 1: LM, polar view, showing tricolporate pollen. x1000. 2: LM, equatorial view showing thick exine and ornamentation. x1000. 3: LM, polar view showing pores structure. x1000. 4: SEM, equatorial view showing indistinct endoapreture and straight colpus. x 3500. 5: SEM, equatorial view, showing bridge of ectoaperture and microechinate exine x5500. 6: SEM, polar view showing small amb and straight tricolpus. x4500. 7: TEM, T.S., in pollen grain, showing spinulose tecta and thick intine. x2500. 8: TEM, L.S., in pollen grains showing ectoexine without columella in pore. x12000. 9: TEM, apreatures area of pollen grains without columella, few spinolose on ectoexine. x12000. 10: TEM, LS, showing thin exine and intine in apertural area. x12000 J. Biol. Sci., 7 (4): 626-631, 2007 628 |
| Table 2: | Dimensions, shape classes, shape and exine structures of the pollen of Solanum species as observed by LM, SEM and TEM |
 | |
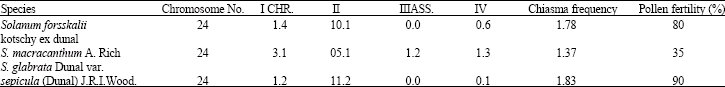
| Table 3: | Meiotic chromosome number, association, chiasma frequency and pollen fertility for the three species of Solanum |
 | |
| CHR. ASS.= Chromosome association | |
POLLEN GRAINS DESCRIPTION
Solanum forsskalii Kotschy ex Dunal: [Fig. 1(1-9)] Sy. S. albicule: Pollen grain is rhomboidal with broad ends in equatorial view and triangular in amb. Colpi with tapering ends equatorially constricted and tectate, faintly perforate. Columellae more or less distinct regularly distributed. Pore lalongate, 4.5 μm wide. Exine, 1.5 μm thick. Sexine is thicker than nexine and microscabrates. Ornamentation homobrochate Fig. 1-3. By SEM, pore lalongates, protruded, colpal membrane not sunken (surfaced) and densely granular. Exine, densely spinulose-non-perforate [Fig. 1(4-6)]. By TEM, pore distinct and circular in shape. Note fastigium-like cavity formed by the slight protrusion of the ectoapertural margins and an intact/continuous endexine. Ectexine tectate-spinulose elements on the tectum are densely and pointed. The columella is less distinct and irregular (Fig. 1 (7-9).
Comment: A common and endemic plant in the flora of Saudi Arabia (Migahid, 1978; Collenette, 1985; 1998). The pollen grains are distinguished by their comparatively long polar axis (24 (26)29 μm) and triangular lobed. The exine is densely spinulose-the intine oily cytoplasmic.
Solanum macracanthum A. Rich. [Fig. 2 1-8)]: Pollen grain is angular-rhomboidal with tapering ends in equatorial view and rounded-circular in amb. Colpi with tapering ends are equatorially constricted and tecta indistinct perforate. Columellae regular distributed and pore rounded. Pore lalongate, 3.4 μm wide. Exine, 1.3 μm thick, sexine is thicker than nexine and microscabrates. Ornamentations are homobrochate, [(Fig. 2 (1-3)]. By SEM, pore lalongate and colpal membrane is sunken and densely granular. Exine is regularly, densely and spinulose [(Fig. 2 (4-6)]. By TEM, apertures: indistinct, ectexine heterogenous, endoexine is lamellated and columella indistinct, [(Fig. 2 (7-8)].
Comment: A common and endemic plant in the flora of Saudi Arabia (Migahid, 1978; Collenette, 1985; 1998). The pollen grains are distinguished by their comparatively long polar axis (20(23)27 μm) and rounded ends and colpi margins in equatorial plane fuse to form abridge-like structure over the pore. The exine is densely spinulose.
S. glabrata Dunal var. sepicula (Dunal) J.R.I.Wood. [(Fig. 3, (1-10)].
Sy. S. sepicula: Pollen grain is angular-rhomboidal with broad ends in equatorial view and triangular in amb. Colpi with tapering ends equatorially constricted. Pore lalongate, 3.6 μm wide. Exine, 1.22 μm thick, sexine is thicker than nexine and microscabrates. Ornamentations are homobrochate [Fig. 3 (1-3)]. By SEM, apertures colpi are some regularly equatorial constricted, pore bridged lalongates. Exine, is scattered spinulose and less near the colpal membrane (Fig. 3 (4-6)]. By TEM, apertures, without columella, ectexine very thin and endexine is thick. Exine, tectate-columellate and very few scattered spinulose (punctate-microspinulose tecta) [(Fig. 3 (7-10)].
Comment: A common and endemic plant in the flora of Saudi Arabia (Migahid, 1978; Collenette, 1985; 1998). The pollen grains are distinguished by their comparatively long polar axis (20(23)26 μm) and triangular lobed. The exine is densely spinulose-the intine oily cytoplasmic.
Cytological characters: The meiotic parameters have been done in Table 3.
The study of meiotic chromosome of the taxa under investigation recorded, 24 as diploid (x = 12) this number is common for most Solanum species in the area (Haroun and Al-Wadi, 1999). Behavior of chromosome during meiosis is normal at diaknesis and metaphase I stages. Few univalent and multivalents were observed for S. forsskalii and S. glabrata var. sepicula with high ratio of bivalent and few abnormal cells is laggard. The pollen fertility of these two species is 80 and 90%, respectively Table 3. Behavior of S. macracanthum chromosome shows some meiotic disorders in form of abnormal cells and irregular pairing. Univalent and multivalent chromosome were also observed and found high compared to that recorded for the other two taxa (Table 3).
DISCUSSION
The present investigation concentrates on the ultra-structures, comparative structure and meiosis of the pollen grains of three taxa of genus Solanum. Palynological data indicated that all pollen are isopolar, symmetrical trizoncolporates or trizonoporates. On the basis of pollen shape class (P/E ratio), two types are regonized, viz., Solanum macracanthum (prolate-subprolate) and S. forsskalii and S. glabrata var. sepicula (prolate-spheroidal). On the basis of ectexine columella, three types are regonized, viz., S. glabrata var. sepicula (regularly columellate), S. forsskalii (less and irregular columellate) and Solanum macracanthum (columella indistinct). Now the scientists believe that the palynological studies can provide more accurate basis for the identification of a plant species. The pollen morphology is useful to identify various species and taxa in their respective families. It is used to identify hybrid plants and to solve the taxonomic problems of medicinal and other economically important plants. Palynology has been very helpful in indication of probable lines of evolutionary trends (Diaz and Lifante, 1991). The basic chromosome number in Solanum is diploid at x = 12, with 2n = 24 (Grant, 1982) like that of most of the other genera belonginig to the subfamily Solanoideae (Hunziker, 1979). Cytological investigation of the species provides that Solanum forsskalii and S. glabrata var. sepicula are stable cytologically, while Solanum macracanthum shows irregular meiosis. Davis and Heywood (1967) reported the same concept for some species of Solanum. According to correlation coefficient between Solanum macracanthum and Solanum forsskalii suggest: that Solanum forsskalii is probably a parent of Solanum macracanthum. The normal chromosome behavior for Solanum forsskalii and S. glabrata var. sepicula could interpret their high pollen fertility (80-90%). Pollen fertility studies investigate plant communities to reveal their genetic variation and the indication of hybridization between different species these results agree with Gonzales et al. (1991) who studied the pollen fertility of 35 species of genus Solanum which was 68-99%. On the other hand palynological investigations reveal strong relationships between Solanum forsskalii and S. glabrata var. sepicula rather than with Solanum macracanthum. The species with a meiotically stable and potentially fertile, apparently without problems in selection for fertilization in crossing and viable seed production programs (Coelho and Battistin, 1998). These results could predict that there is a high potential of cross-pollination in area of the studded species with some related species especially to Solanum macracanthum (Haroun and Al-Wadi, 1999). These data could furnish adequate information that supports the relation characters between the species. The data derived from palynological and cytological characters of the examined species could also contribute to the taxonomy of the family Solanaceae family. These studies will be very helpful for compiling such work and identifying the flora according to their pollen morphology, fertility and economic importance for wider circulation.
KEY TO THE SPECIES
1-+_triporates pollen grains in SEM ……YYYSolanum forsskalii
-tricolporates pollen grains in SEM ……YYYY.……2
2-densly columelate exine……YYYYY. Solanum glabrata var. sepicula
3-columella indistinct……YYYYYY...…....Solanum macracanthum
ACKNOWLEDGMENTS
The authors thank M.H. Aseeri Department of Biological Sciences, College of Sciences, King Khalid University, Abha, for his help to collect the herbarium materials and photographs by light microscope. In addition, we thank Dr. Z. Baka Department of Biological Sciences, College of Sciences, King Khalid University, Abha, for his proof of the manuscript.
REFERENCES
- Coelho, L.G.M. and A. Battistin, 1998. Meiotic behavior of Adesmia DC. (Leguminosae-Fabiodeae) species native to Rio Grande do Sul, Brazil. Genet. Mol. Biol., 21: 403-406.
Direct Link - El Ghazali, G., 1993. A study on the pollen flora of Sudan. Rev. Palaeobot. Palynol., 76: 99-345.
Direct Link - Reynolds, E.S., 1963. The use of lead citrate at high pH as an electron-opaque stain in electron microscopy. J. Cell Biol., 17: 208-212.
CrossRefPubMedDirect Link