
Arshad Ali
Department of Plant Protection, Faculty of Agricultural Sciences, Aligarh Muslim University, Aligarh, India
Parvez Qamar Rizvi
Department of Plant Protection, Faculty of Agricultural Sciences, Aligarh Muslim University, Aligarh, India
Journal of Biological Sciences
Year: 2007 | Volume: 7 | Issue: 8 | Page No.: 1478-1483







ABSTRACT
The performance of Coccinella septempunctata was investigated on five-aphid species viz., Lipaphis erysimi, Aphis craccivora, Hyadaphis coriandri, Rhopalosiphum nymphae and Macrosiphum rosae under controlled conditions (25±1°C, 70±5% RH and 12 h L: 12 h D). The overall developmental period of C. septempunctata was found significantly longer on L. erysimi and shorter on M. rosae. The grubs, however, required significantly longer developmental period on H. coriandri and shorter on M. rosae. Nonetheless, the adult longevity was higher on L. erysimi and shorter on M. rosae. Between the sexes, the female required longer period for its development than male with respect to all aphid species. With regard to predation, the grubs consumed maximum H. coriandri, whereas, adult preferred L. erysimi. The last larval stage (grub 4), irrespective of aphid species, devoured more aphids than the other grub stages. The predation by female of all aphid species was more as compared to male. The overall predation by C. septempunctata was significantly higher on L. erysimi than other species. A linear correlation curve showed the dependency of predation on the developmental period of C. septempunctata.
PDF Abstract XML References Citation
How to cite this article
Arshad Ali and Parvez Qamar Rizvi, 2007. Development and Predatory Performance of Coccinella septempunctata L. (Coleoptera: Coccinellidae) on Different Aphid Species. Journal of Biological Sciences, 7: 1478-1483.
DOI: 10.3923/jbs.2007.1478.1483
URL: https://scialert.net/abstract/?doi=jbs.2007.1478.1483
DOI: 10.3923/jbs.2007.1478.1483
URL: https://scialert.net/abstract/?doi=jbs.2007.1478.1483
INTRODUCTION
Aphids are a diverse group of plant feeding insects belong to family Aphididae of order Hemiptera, predominantly found in temperate climate zones such as North America, Europe, Central and Eastern Asia (Nelson and Rosenheim, 2006). Approximately 4000 species of aphids have been described feeding over 250 agricultural and horticultural crops throughout the world. The eggs of aphids develop parthenogenetically within the mother (female body) and nymphs born after few days, hence, population density increases rapidly (Blackman and Eastop, 2000). In the early stage of infestation, adult have no wings but as they become crowded, winged forms appeared in subsequent generations for dispersal from one plant to another (Braendle et al., 2005). They are found primarily on the growing points of the host plants, including tips, flowers and developing pods and cover whole plant at high density (Blackman and Eastop, 2000). They cause damage directly by sucking cell sap, secrete honeydew resulting in development of sooty mould on leaves and shoots and indirectly as vectors of certain plant viruses (Kennedy et al., 1962; DiFonzo et al., 1997; Raboudi et al., 2002). Many insecticides have been recommended for its control but these are known to cause mortality of the beneficial organisms, residue problem, hazards to man and animals besides polluting the environment. On several occasions, insecticidal applications have accentuated the aphid population and quite often resulted in outbreaks (Orlandao et al., 1970). Therefore, use of natural enemies in biological control is a good pest management tactic to minimize the population level of invasive pests (Delfosse, 2005).
Ladybirds are generalized predator that feed on a diverse range of foods. Aphids are the principal food of ladybirds, whereas coccids, mites, honeydew, pollen, nectar and mildew are recorded as secondary foods (Bianchi et al., 2004; Deligeorgidis et al., 2005). Among ladybeetle group, seven spotted ladybird, Coccinella septempunctata is one of the potential predators of aphids in India (Pervez and Omkar, 2005). Both larvae (grubs) and adults of C. septempunctata feed on aphids. The development and potential feeding of coccinellids vary with their choice food and change with the environment condition. Therefore, present study was initiated to determine the development and predatory performance of C. septempunctata on five aphid species (Lipaphis erysimi, Aphis craccivora, Hyadaphis coriandri, Rhopalosiphum nymphae and Macrosiphum rosae) under controlled conditions. The information, so generated, will provide a preliminary step in exploitation of C. septempunctata in biological control of above mentioned aphids of economic significance.
MATERIALS AND METHODS
Aphid cultures: To maintain the culture of different aphid species, various crops viz., Indian mustard (Brassica juncea L.), common bean (Phaseolus vulgaris L.), coriander (Coriandrum sativum L.) and ornamental plant, verbena (Verbena laciniata) were grown in the micro-plots sized 3x4 meter (each replicated thrice) in winter season of year 2005 and 2006 and five year old rose (Rosa indica) plants were maintained at experimental fields of the Department of Plant Protection, Faculty of Agricultural Sciences, Aligarh Muslim University, Aligarh, India. The infestation of aphid appeared naturally in the last week of December on indian mustard, coriander and rose plants, whereas, on common bean and verbena it appeared in the second week of January. They multiply parthenogenetically and their density increased rapidly on the growing points of plants. The aphid species collected from the respective crops were got identified from the Laboratory of Aphidology in the Department of Zoology, University of Kalyani, West Bengal. The aphid species, which attacked on indian mustard, common bean, coriander, verbena and rose plants were identified as: Lipaphis erysimi, Aphis craccivora, Hyadaphis coriandri, Rhopalosiphum nymphae and Macrosiphum rosae, respectively. For natural control of these aphid species, various ladybird species were also attributed with the aphid colonies on respective host plants. Of all the ladybird beetles, Coccinella septempunctata was predominantly predating all aphid species.
Ladybird cultures: The grubs and pupae of Coccinella septempunctata were collected from naturally infested field crops and kept in BOD incubator for adult emergence. The condition maintained in BOD incubator was 25±1°C temperature coupled with 70±5% relative humidity and the photoperiod of 12 h L: 12 h D.
Freshly emerged adult beetles were reared on respective aphid species in pairs in petridishes (90 mm diameter and 10 mm height). A blotting paper was spread over its inner surface for egg laying. The eggs laid by females were counted and transferred in other petridishes, with the help of soft camel hair brush, for hatching. After hatching, a total of twenty newly hatched (zero day old) grubs of predator species were reared individually in plastic vials (measuring 4.0 cm in diameter and 6.0 cm in height) on respective aphid species. Each experiment was replicated thrice. For newly born grubs, 30-40 nymphs of respective aphid species with the twig of respective plants were provided as food. The number of aphids was subsequently increased, reaching maximum up to 100 nymphs daily with advancement in the age of grubs. This procedure was followed till pupation. Side by side, the exuvae found in each of vials was removed when the grub entered into next instar. Total number of aphid consumed by each grub, dead and unconsumed aphids were counted daily and were replaced by fresh aphids. After the emergence of adult (male and female), they were again provided with a minimum of 100 aphid nymphs daily, till their death. In this way, daily consumption by grub, male and female of C. septempunctata on different aphid species was recorded. Sexing of adult was made as per suggestion of Sathe and Bhosale (2001).
Statistical analysis: Development and predation at different life stages of C. septempunctata was recorded with respect to aphid species. To determine statistical difference, ANOVA was computed and the mean values were compared using Duncan’s multiple range test (p≤0.05). Correlation between developmental period and predation of C. septempunctata on different aphid species was also calculated.
RESULTS
Development: The developmental period of all immature stages of C. septempunctata fed with H. coriandri was significantly longer than those fed with L. erysimi, R. nymphae, A. craccivora and M. rosae (Fig. 1). However, the longevity of the adult (male and female) was significantly higher on L. erysimi and lower on M. rosae. The female required longer period for their development as compared to male with respect to all aphid species (Fig. 1).
The total developmental period of immature stages was statistically different among the aphid species. Both, total grub and total pupal period of C. septempunctata showed significantly high value (17.00±0.533 and 13.30±0.144 days) on H. coriandri against the low (15.70±0.186 and 9.40±0.153 days) on M. rosae, respectively. However, the overall developmental period was significantly longer (62.75±1.430 days/progeny) on L. erysimi and shorter (47.55±0.678 days/progeny) on M. rosae (Table 1).
Duncan’s multiple range test on different developmental stages (total grub, total pupal and average adult) of C. septempunctata revealed a significant difference (p≤0.05) with respect different aphid species.
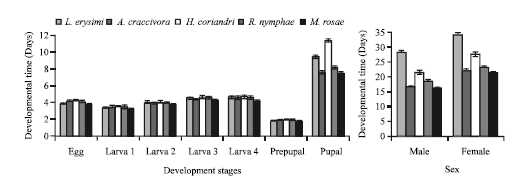 | |
| Fig. 1: | Developmental time (days±SE) of Coccinella septempunctata on different aphid species |
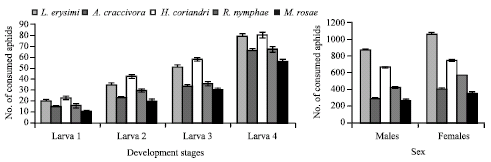 | |
| Fig. 2: | Predation (number of aphids±SE) of Coccinella septempunctata on different aphid species |
| Table 1: | Development and predation of C. septempunctata on different aphid species |
 | |
| Given values are developmental period in days±SE and number of aphid consumed±SE; Same alphabets showing non-significance and different alphabets indicate significance in accordance to Duncan’s multiple range test | |
Predation: The predatory response of different developmental stages of C. septempunctata exhibited significant difference with respect to all aphid species. The grub stages devoured voraciously H. coriandri followed by L. erysimi, A. craccivora, R. nymphae and M. rosae. The last larval stage (grub 4) consumed more aphids than the other grub stages. The adults (male and female) ate significantly higher L. erysimi than other aphid species. Nonetheless, the consumption of female was higher in contrast to male with respect to all aphid species (Fig. 2).
It was observed that, the overall feeding efficiency of C. septempunctata was significantly higher on L. erysimi (1142.75±18.706 aphids/progeny) followed by H. coriandri (901.65±15.280 aphids/progeny), R. nymphae (638.05±12.262 aphids/progeny), A. craccivora (486.30±10.367 aphids/progeny) and M. rosae (436.80±10.399 aphids/progeny) (Table 1). Duncan’s multiple range test also exhibited that the predation of C. septempunctata was significantly different (p≤0.05) with respect to all aphid species.
When the correlation was made between the predation and the developmental period of C. septempunctata, a linear curve was obtained with respect to all aphid species. Figure 3 clearly showed that the predation of C. septempunctata depends on the developmental period; it increased with the advancement in age and registered the highest R2-value (0.9994) on L. erysimi and the lowest (0.9942) on M. rosae.
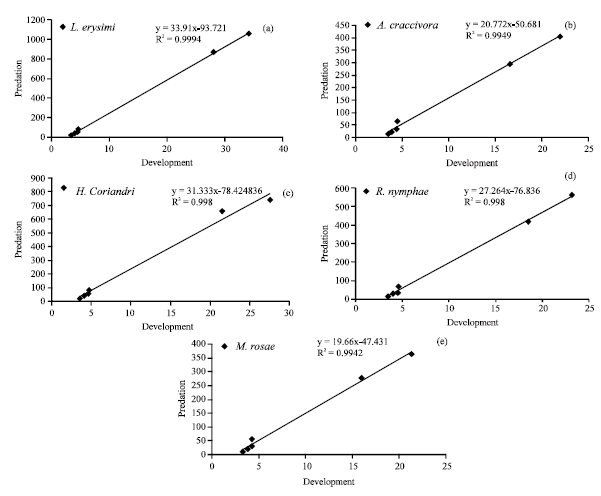 | |
| Fig. 3: | Correlation between the development and predation of C. septempunctata on different aphid species; (a) L. erysimi, (b) A. craccivora, (c) H. coriandri, (d) R. nymphae and (e) M. rosae |
DISCUSSION
The observations revealed that the developmental period of C. septempunctata varied significantly with respect to aphid species. The variation could be attributed on account of difference in size/biomass of different aphid species (Bhadauria et al., 2001). In present studies, it was also found that the female gave more attention to survive on all aphid species in comparison to male. Atlihan and Kaydan (2002) and Omkar and James (2004) also held the same opinion.
In our experiments, grubs of C. septempunctata tended to eat maximum H. coriandri than other species, probably due to small size of the prey (Alikhan and Yousuf, 1986; Singh et al., 1994). The adults (male and female) devoured maximum L. erysimi in comparison to other aphid species. This observation is in consonance with the findings of Hodek (1973) Kring and Gilstrap (1984) and Lee and Kang (2004).
Advancement in the age of C. septempunctata clearly showed the increase in the consumption of prey and has been well supported by the findings of other workers (Elliott et al., 1994; Babu, 1999). Besides this, host specificity (Elliott et al., 2000; Aslan and Uygun, 2005) and plant characteristics (Hodek and Honek, 1996) do influence on the predation potential, which have also been observed in the present investigation.
Coccinella septempunctata adult devoured more aphids than grubs, such investigations are in consonance with the judgment of Haque and Islam, 1982 and Singh et al., 1994. However, between sexes, female consumed more aphids than the male (Singh and Singh, 1994; Ba M’Hameed and Chemseddine, 2001; Brown, 2004; Omkar and Mishra, 2005).
It could be accomplished from the present findings that Lipaphis erysimi gave more response to C. septempunctata, under laboratory conditions, in terms of development and predatory preference as compared to other aphid species (A. craccivora, H. coriandri, R. nymphae and M. rosae) and hence, offer preliminary information to introduce C. septempunctata in the biological control programme of aphids of economic significance.
ACKNOWLEDGMENTS
The authors are thankful to Professor Samiran Chakraborty, Department of Zoology, University of Kalyani, West Bengal, for identifying the aphid species and also grateful to the authorities of the Department of Plant Protection, Faculty of Agricultural Sciences, to support our research.
REFERENCES
- Atlihan, R. and M.B. Kaydan, 2002. Development, survival and reproduction of three coccinellids feeding on Hyalopterus pruni (Geoffer) (Homoptera: Aphididae). Turk. J. Agric. For., 26: 119-124.
Direct Link - Ba-M`Hameed, T. and M. Chemseddine, 2001. Assessment of temperature effects on the development and fecundity of Pullus mediterraneus Fabr. (Coleoptera: Coccinellidae) and consumption of Saissetia oleae Olivier (Homoptera: Coccidae) eggs. J. Applied Entomol., 125: 527-531.
Direct Link - Bianchi, J.J., A. Felix and W. Werf, 2004. Model evaluation of the function of prey in non-crop habitats for biological control by ladybeetles in agricultural landscapes. Ecol. Modelling, 171: 177-193.
Direct Link - Braendle, C., I. Friebe, M.C. Caillaud and D.L. Stern, 2005. Genetic variation for an aphid wing polyphenism is genetically linked to a naturally occurring wing polymorphism. Proc. R. Soc. Lond., 272: 657-664.
Direct Link - Brown, M.W., 2004. Role of aphid predator guild in controlling spirca aphid population on apple in West Virginia, USA. Biol. Control, 29: 189-198.
Direct Link - Deligeorgidis, P.N., C.G. Ipsilandis, M. Vaiopoulou, G. Kaltsoudas and G. Sidiropoulos, 2005. Predatory effect of Coccinella septempunctata on Thrips tabaci and Trialeurodes vaporariorum. J. Applied Entomol., 129: 246-249.
CrossRefDirect Link - DiFonzo, C.D., D.W. Ragsdale, E.B. Radcliffe, N.C. Gudmestad and G.A. Secor, 1997. Seasonal abundance of aphid vectors of potato virus Y in the Red River Valley of Minnesota and North Dakota. J. Econ. Entomol., 90: 824-831.
Direct Link - Elliott, N.C., B.W. French, J.G.J. Michels and D.K. Reed, 1994. Influence of four aphid prey species on development, survival and adult size of Cycloneda ancoralis (Germar) (Coleoptera: Coccinellidae). Southwest. Entomol., 19: 57-61.
Direct Link - Elliott, N.C., R.W. Kieckhefer and D.A. Beck, 2000. Adult coccinellid activity and predation on aphids in spring cereals. Biol. Control, 17: 218-226.
Direct Link - Kring, T.J. and F.E. Gilstrap, 1984. Efficacy of the natural enemies of grain sorghum aphids (Homoptera: Aphididae). J. Kans. Entomol., 76: 57-62.
Direct Link - Lee, J.H. and T.J. Kang, 2004. Functional response of Hormonia oxyridis (Pallas) (Coleoptera: Coccinellidae) to Aphis gossypii Glover (Homoptera: Aphididae) in the Laboratory. Biol. Control, 31: 306-310.
Direct Link - Nelson, E.H. and J.A. Rosenheim, 2006. Encounters between aphids and their predators: The relative frequencies of disturbance and consumption. Entomol. Exp. Applic., 118: 211-219.
Direct Link - Omkar and B.E. James, 2004. Influence of prey species on immature survival, development, predation and reproduction of Coccinella transversalis Fabricius (Col., Coccinellidae). J. Applied Entomol., 128: 150-157.
Direct Link - Omkar and G. Mishra, 2005. Preference-performance of a generalist predatory ladybird: A Laboratory study. Biol. Control, 34: 187-195.
Direct Link - Pervez, A. and Omkar, 2005. Functional responses of coccinellid predators: An illustration of a logistic approach. J. Insect Sci., 5: 5-5.
PubMedDirect Link - Raboudi, F., A. Ben Moussa, H. Makni, M. Marrakchi and M. Makni, 2002. Serological detection of plant viruses in their aphid vectors and host plants in Tunisia. EPPO Bull., 32: 495-498.
Direct Link - Singh, V.S., R.P. Yadav and R. Singh, 1994. Post-embryonic development, survival rate and predation potential of Coccinella septempunctata Linn. In: relation of mustard aphid (Lipaphis erysimi Kalt.). J. Entomol. Res., 18: 5-10.
Direct Link