
B.C. Li
Institute of Animal Husbandry and Veterinary, Jiangsu Academy of Agricultural Science, Nanjing, 210014, China
F. Yu
Institute of Animal Husbandry and Veterinary, Jiangsu Academy of Agricultural Science, Nanjing, 210014, China
Q. Xu
Institute of Animal Husbandry and Veterinary, Jiangsu Academy of Agricultural Science, Nanjing, 210014, China
L. G. Ni
Institute of Animal Husbandry and Veterinary, Jiangsu Academy of Agricultural Science, Nanjing, 210014, China
G. H. Chen
Institute of Animal Husbandry and Veterinary, Jiangsu Academy of Agricultural Science, Nanjing, 210014, China
X.M. Cheng
Not Available
H.H. Musa
Not Available
T.Z. Liu
Not Available
Journal of Biological Sciences
Year: 2007 | Volume: 7 | Issue: 8 | Page No.: 1331-1338







ABSTRACT
Chicken embryos from stage 15 to stage 45 were studied by means of serial section and light microscopy in order to investigate the relationship between the spermatogonium and the testicular development in early chicken embryos. The results showed that the glycogen in the PGCs (primordial germ cells) cytoplasm reduced gradually at the stage 22-28 (3.5th-5th hatching day). On the stage 29 (6th hatching day), the gonad of the embryo appeared the feature of testis and the glycogen in the PGCs cytoplasm reduced further. On the stage 31 (7th hatching day), the differentiation of ovary or testis was obvious and the glycogen in the PGCs cytoplasm later disappeared. On the stage 34 (8th hatching day), the testicular cord had began to differentiate. The each cord had been solid tubule with the spermatogonium. On the stage 35-37 (9th-11th hatching day), the number of testicular cord had been increased following embryonic development. The spermatogoniums were monolayer located in cord. At this stage, fewer sustentacular cells had been differentiated, while difficult to distinguish. On the stage 38-40 (12th-14th hatching day), the seminiferous tubules had been formed typically, the number of spermatogonium increased as well as sustentacular cells and the interstitial cells in seminiferous distributed in groups. On the stage 40-45 (16th-19th hatching day), the testis on the right was a little bigger than that of the left, the number of spermatogonium increased obviously and appeared like clusters of grape in the middle of seminiferous. Lumina in seminiferous tubule had formed and spermatogonium aligned with different layers.
PDF Abstract XML References Citation
How to cite this article
B.C. Li, F. Yu, Q. Xu, L. G. Ni, G. H. Chen, X.M. Cheng, H.H. Musa and T.Z. Liu, 2007. Relationship Between Spermatogonium and Testicular Development in Early Chicken Embryos. Journal of Biological Sciences, 7: 1331-1338.
DOI: 10.3923/jbs.2007.1331.1338
URL: https://scialert.net/abstract/?doi=jbs.2007.1331.1338
DOI: 10.3923/jbs.2007.1331.1338
URL: https://scialert.net/abstract/?doi=jbs.2007.1331.1338
INTRODUCTION
Since the development and structure of the vertebrate ovaries were studied by Balfour, the genesis and development of the human and animal’s gonad have been investigated endlessly. There were discussions of gonadal development in the monographs about animal embryonic development and had mainly obtained the definite conclusion about the process of the gonadal genesis and development (Paidom et al., 1979; Hao-Jian, 1984; Tie Sheng, 1987). The organizational structures of eugamic chicken’s generative organ had been researched by Su-Zhu et al. (1998), the results showed that there were differences of female and male’s gonad between the chicken and the mammal, seminiferous epithelium of seminiferous tubules in the chicken’s testes formed many cell trabeculums, but the mammal did not have this structure. The reports on the genesis and development changes of the chicken gonadal organizational structures, the appearance period and the morphology of the spermatogonial cells and the Sertoli’s cells were scattered. Chicken embryonic spermatogonial cells originated outside the gonad, their forefather Primordial Germ Cells (PGCs) arrived in developing gonadal ridge by complicated migrated process. There were few reports about PGCs’ intimate change and distribution after they arrived in spermary in male (Dubois and Croisille, 1970; Zaceanti et al., 1990). In present study, we observed the genesic and developmental process of the chicken embryonic testes daily and studied the genesic and developmental relationship between the spermatogonial cells and the testis systematically.
MATERIALS AND METHODS
Materials: This study was conducted from June 2005 to May 2006 in Laboratory of animal science of Yangzhou University, Yangzhou City, China. Materials used for this study were recessive leuco-feather chick embryos 50 h-19 days of incubation. Eight chick embryos of every stage were chosen. Embryos of 50 h-7 day were fixed in Rossman’s and embryos of 7-19 day were fixed in formaldehyde solution.
Methods: Dislodgment of blastoderms Use a forceps to knock at the blunt end of hatched egg. Abscise eggshell and shell membrane carefully to get a port of 3-4 cm in diameter. Then cut the blastoderm along borderline with eye scissors when egg yolk and blastoderm were exposed. Put blastoderms taken out of eggs by medicine spoon into a culture capsule containing warm isotonic Na chloride solution (38-39°C). One side which has yolk membrane is placed in supravergence. Slosh the culture capsule lightly to get egg yolk and golk membrane away from blastoderms. Next, pour out the most of isotonic Na chloride. Then poach blastoderms in fresh warm isotonic Na chloride. Repeat several times. Finally, put clean blastoderms into Rossman’s or formaldehyde solution to fix over night.
Dislodgment of embryos: The method of dislodgment is the same to blastoderms. Dislodge embryos carefully from blunt end of eggs with forceps. Put embryos into a culture capsule containing warm isotonic Na chloride. After snipping the saccus vitellinus and extraembryonic membranes, poach embryos for two times. Then fix embryos in Rossman’s or 4% formaldehyde solution.
Disposal of sections: Materials embed after fixed in paraffin. The serial sections were made 5-7 um thick. The sections of 50 h-7 days were stained by the PAS reaction or HE. The sections of 7-19 days were stained by HE Then the serial sections were observed by Olympus light microscopy (Olympus IX51, Japan) and record PGCs of every stage and the differentiation of genital ridge. After 10 days of incubation, the discrimination of paroecious can be observed easily based on the appearance and size of genitical gland.
Morphology identification of testis cells: Preparation of testis paraffin section and staining:Put poult cock testes removed amicula and albuginea in Bouind to fix 24-48 h at 4°C. Then after anhydration, embedment, section and HE staining, observe the sections by light microscopy and take photos. HE staining of testis cells suspension: Centrifuge testis cells suspension at 100 rpm for 8 min and then pour out clear supernatant liquid. Fix cell aggregate in 2.5% glutaraldehyde for 30 min and centrifuge them at 800 rpm for 5 min. Add some fixation fluid to praecipitatum to float cells. Then drop discus, H.E stain, observe and shoot.
RESULTS
The appearances of various kinds of cells were observed by Olympus light microscope. After PAS (periodic acid-Schiff reaction)-staining, PGCs were round (Fig. 1a), which cell nucleuses were large but not stained and cytoplasm became prunosus. After H.E staining, spermatogonial cells were round (Fig. 1b), which volumes were larger than sertoli cells. Nucleus of spermatogonial cells were deeper stained. The shape of sertoli cells was abnormity, such as round, metuliform and so on. Their volumes were smaller than spermatogonial cells. And the colour of H.E staining was lighter than spermatogonial cells. Cellula intersitialis generally distributed groupingly among contorted seminiferous tubules.
According to different embryonic incubation time, the results of development of testis in chick embryos and their relationship with spermatogenous cells were as follow:
At stage 15 (n = 6), the 24-27 somites (incubation time: 50-55 h). PGCs accumulated and intruded between the caudal of the embryo and the vitelline artery. Almost whole portions of Somatopleura and the coelomic epithelium covering the coelomic cavity became thicken. At this stage, PGCs were not found in coelomic epithelium and its neighboring area.
At stage 16 (n = 6), the 26-28 somites (incubation time: 53-56 h). We can find 3-5 layers of cells of the thickened portion of the coelomic epithelium limited in the 180 μm area from splanchnopleme to coelomic angle, while the region between the coelomic angle and mesonephros which corresponds to the site of gonad in later stage did not thicken, structured by single complanate or columnar cells. At this developmental stage, the number of PGCs increased, PGCs distributed between the caudal of the embryo and the vitelline artery, ranging in length from 0.8 to 1.9 mm in this territory, some PGCs were seen to be incorporated into the thickened coelomic epithelium and others were found in the mesenchyme nearby and in the blood vessel, in some cases PGCs were found between endothelial cells of the vessel. PGCs occasionally found in the splanchnopleure, no PGCs were found in the somatopleure (Fig. 2). It is noted that when the embryo developed to stage 16, PGCs were found in the thickened epithelium and the mesenchyme of the head or blood vessel of the caudal.
At stage 17 (n = 6), the 29-32 somites (incubation time: 52-64 h). Many PGCs concentrated in the site where perhaps gonad formation settled. At this stage, most thickened region of the coelomic epithelium translocated toward the mesonephros, though a small part was still present at the proximal portion of the splanchnopleura. Many PGCs were found in the thickened region of the coelomic epithelium and blood vessels.
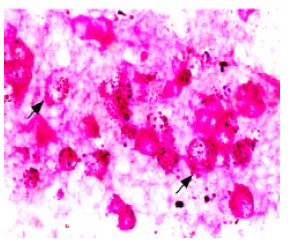 | |
| Fig. 1a: | Chicken embryonic PGCs stained by PAS, x1000; |
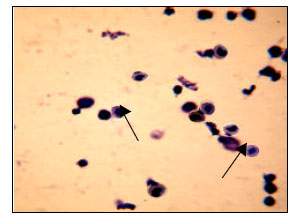 | |
| Fig. 1b: | Chicken spermatogonia stained by HE (the arrow’s direction is spermatogonia), x400 |
 | |
| Fig. 2: | The distribution of PGCs in the chicken epithelium of the embryo (8), stained by H.E., x400 |
At stage 18 (n = 6), the 33-36 somites (incubation time: 72 h). At this developmental stage, the dorsal aorta contacted with each other at the midline of embryos and left and right splanchnopleure start joined at their proximal portions to form the dorsal mesentery. The thickening of the coelomc epithelium was found in the region from the coelomic epithelium to the future gonadal site; however, some thickened epithelium was still present in the proximal of the splanchnopleure. Most PGCs were found to be involved in these thickened epithelia, some were found in the mesenchyme nearby. As the thickened portion of the epithelium moved toward the mesonephros, the localizing area of the PGCs was alternated. Genital gland presented highly narrow stripe stenosal from the observation of sagittal plane sections.
At stage 22 (n = 6), (incubation time: 82 h). The embryo began to form mesonephric duct, while in the inner edge of the mesonephric, the genital ridge has formed and began to separate to the mesonephros, especially at the anterior region. Many PGCs concentrated on the genital ridge, about 50-60 in each section, which indicated the PGCs and the thickened portion of the coelomic epithlium form the genital ridge together. Stained by PAS can find the cytoplasm easily reaction with the PAS, but the nucleus could not. The glycogen in the cytoplasm began to resolve in most PGCs, germinal ridge located in the conterminous region between the mesonephros and the hind-kidney, the shape and the size of the PGCs in genital ridge changed greatly, some were round oblong and some begin to divide. Genital ridge and the wall of mesonephric duct have no conspicuous margin and appeared to have the same portion of parenchyma (Fig. 3). Therefore, we presumed that some mesonephric cells participated in the development of genital ridge.
At stage 25 (n = 6), (incubation time: 5 days). The mesonephros development conspiciousness, primitive gonad closed to its inner side, separated obviously to the mesonephros. The primitive gonad was big and long, many PGCs located in it, the cytoplasmic glycogen was resolving, like flower. But still can identify the basic shape of the glycogen in the cytoplasm. The undifferentiated gonad obviously longer than that at 4 days, but still cannot identify the sex (Fig. 4).
At stage 29 (n = 6), (incubation time: 6 days). The genital ridge became more conspicuous, the neuropile closed to the mesonephros and the cortex of the ventral side can be distinguished. Some PGCs located in the neuropile of the genital gonad, some began to migration toward the cortex, cytoplasmic glycogen resolved and distributed disorder, which indicated the gonad began to differentiation. Stained by HE, there was one large cell like PGCs in the epithelium of the genital gonad and has obvious gap under it, the genital gonad separated conspicuously into two parts. Both of the embryonic genital gonads showed ovarian characteristic, which indicated the sex began to differentiation at this stage (Fig. 5).
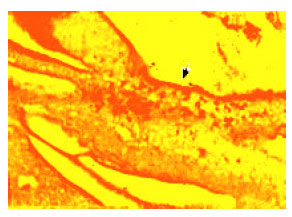 | |
| Fig. 3: | Many PGCs accumulated in gonad and lateral part of mesonephros posterior segment (8), stained by PAS, x100 |
 | |
| Fig. 4: | PGCs regularly arranged in the distal part of gonad-in the germinal epithelium, cytoplasmic glycogen of PGCs was resolving (8), stained by PAS, x400 |
At stage 31 (n = 6), (incubation time: 7 days). By PAS-staining was cannot find PGCs, gonad on the left like thread, bias located in the inner wall of the mesonephros, gonad on the right lightly short than the left, showed rectangle. Observed the section, the gonad obvious separated into two layers, the exterior contained blood vessel and net like cells, showed ovarian characteristic. Observed another section, the epitheliac cell had two layers, like the early tunica albuginea of the testis development progress of the mammalia, closed to epithelium was one non-classic rhomboidal layer cells, showed testiculate characteristic. There was high stained cell mass cord called in the parenchyma, the testicle cord separated by the early tunica albuginea that made it separated from the epithelium (Fig. 6).
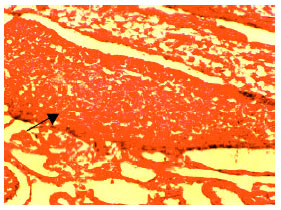 | |
| Fig. 5: | The whole slice of ovary contained medulla and cortex, while the testis had not (8), stained by HE, x100 |
 | |
| Fig. 6: | The testicular cord and albuginea in early stage (8), stained by HE, x400 |
At stage 34 (n = 6), (incubation time: 8 days). The outside of testicle were only arranged with an order lamella of cells, which appeared cardinal red after HE staining. There was no germinal epithelium like ovary. Inside layer detached clearly from outside was tunica albuginea which comprised by 2-3 layers of oblate cells. The parenchyma part was looser, in which distributed a lot of germ cells and appeared the original seminiferous tubule (testicle cable) surrounded with very thin basement membrane. The inner part distributed a lot of great circular cells- spermatogonial cells with round and dyed deeply nuclei and lightly dyed periplast. Spermatogonial cells arranged in the periphery of testicular cord with a single layer. A few cells started to differentiate. Except that spermatogonial cells were great volume, there were no obvious morphology characters to distinguish spermatogonial cells, mesenchymal cells and sertoli cells. (Fig. 7).
At stage 35 (n = 6), (incubation time: 9 days). Two sides of the sex gland of male embryo were same in width. The initial albuginea could be discerned dimly and had two layers of pavement cells. The testicular cord presented the shape of infarctate round, in which, spermatogonial cells distributed with the radiate form and the spermatogonial cells in testicular cord arranged in an orderly round.
At stage 36 (n = 6), (incubation time: 10 days). Ovary tissue and testis tissue could be distinguished obviously from their morphology and the structure. The whole testis was like a clog, the whole slice showed that its bilateralis were thin, the center was wide and edges were regular. There was no obvious discrimination between cortical substances with medullary substance. The amount of testicular cord increased and still were infarctate form. Spermatogonial cells lined a layer in the ambitus of testicular cord and outside covered by basilar membrane. Minority of ovalis or conical cells presented along the basilar membrane and formed contorted seminiferous tubules cord. A few amount of mesenchymal cells distributed between these cords. Few sertoli cells mainly distributed along with the membrane. There was no obvious difference between sertoli cells with other cells in the morphous. Contorted seminiferous tubules cord was still not typical.
At stage 37 (n = 6), (incubation time: 11 days). The morphology of the testis was similar to that on 10th day. The amounts of seminiferous tubules increased while the lumens remained solid center as before. Spermatogenous cells arranged singly in the lumens, only a few lumped or catenated, which arrayed regularly as a wrap in the seminiferous tubules. Some spermatogenous cells began to stick basal membrane tightly and some did not. A few Sertoli cells with little volume, unsteady round or pyramidal shape started to appear. There were small amounts of Sertoli cells in the seminiferous tubules, which appeared like spermatogenous cell. Albuginea presented unistrate flat cell. There displayed redundant seminiferous tubules in the spermary. At the same time, spermatogenous cells were large, irregular and some of them stayed at the status of cleavage. Seminiferous tubules showed circle and large ellipse shape (Fig. 8).
At stage 38 (n = 6), (incubation time: 12 days). One head of testicle began to broaden. In the internal, there was an evident feature of seminiferous tubules and the cross-section began to scale-up, but it also remained solid center, as well as on 11th day. Spermiogoniums’ irregular array was botryoidalis in the center of seminiferous tubule of testicular cord, where were also small amounts of Sertoli cells with blurred morphology. The amounts of mesenchymocytes increased, but remained a few and scattered. A few of them were distributed in the seminiferous tubules.
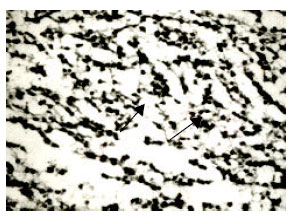 | |
| Fig. 7: | The testicular cord (8), stained by HE, x400 |
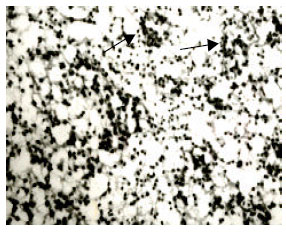 | |
| Fig. 8: | The testicular cord (8), stained by HE, x400 |
At stage 40 (n = 6), (incubation time: 14 days). Typical seminiferous tubules were evident and still remained solid center. Spermatogenous cells scattered distributed at the verge and centre of seminiferous tubules. There were no cytoplasmic bridges between spermatogenous cells. The number of spermatogenous cells increased slightly, their volume smaller than spermatogenous cell at the stage 37, which maybe caused by cell division. The amounts of spermatogenous cell groups with botryoidalis or catenat array increased slightly. Number of Sertoli cells in the seminiferous tubules and mesenchymal cells between the seminiferous tubules increased continually and some of them distributed in group.
At stage 42 (n = 6), (incubation time: 16 days). The volume of testis continued to increase and became like soybean valvula in the end. The morphology of seminiferous tubules did not alter and the tubules began to form. Most spermatogenous cells were distributed in the center of seminiferous tubules. Some of them stick to the basal membrane, some alone and the amount was increasing with the shape somewhat larger than mesenchymal cells, botryoidalis or like chain. Distributed in the seminiferous tubules, the amounts of Sertoli cells increased somewhat, the diastem of them were great. Between the seminiferous tubules, much more mesenchymal cells presented. There were also connective tissues and blood vessels between Sertoli cells (Fig. 9).
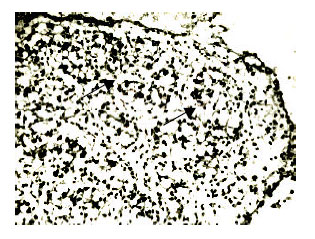 | |
| Fig. 9: | The testicular cord (8), stained by HE, x400 |
At stage 43 (n = 6), (incubation time: 17 days). There was little difference between the volume of two testis, the right was little bigger than the left and the slice of them was sole-like. The amount of seminiferous tubules increased obviously, spermatogenous cells were botryoidalis distributed in the center of seminiferous tubules. Sertoli cells distributed along with tubal wall of seminiferous tubule, the shape of which was irregularand like a cone. There were a large number of Sertoli cells and some connective tissues between the seminiferous tubules.
At stage 44 (n = 6), (incubation time: 18 days). Testes paraffin sections from the cocks of 18 day embryonic age were observed by light microscope. The main component of testes from 18 day embryonic age was mesenchyme and developmental contorted seminiferous tubules. Mesenchyme was mainly made of mesenchymal cells. Sertoli cells and gonocyte which adhered tightly to basal membrane were found in contorted seminiferous tubules. Round or ellipse gonocytes were larger and had the larger nucleus. The nuclei were stained superficially. Sertoli cell nuclei were smaller and inordinate. But cell nucleus were stained darkly (Fig. 10).
At stage 45 (n = 6), (incubation time: 19 days). Seminiferous tubules appeared closely and clearly. spermatogenous cells big and almost round, which scattered or strung out in the center of seminiferous tubules. There were a long strip of cells distributed between the seminiferous tubules, should be some pristine connective tissues. Sertoli cells were nearly same with those on 16-17 days, just the amount increased.
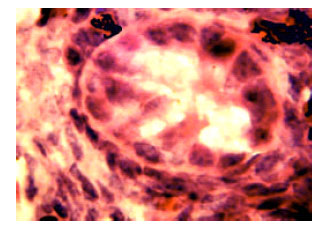 | |
| Fig. 10: | Morphologic character of 18 day chicken embryo testis organ after hatching. (Long arrow is gonocytes and short arrow is sertoli cells) x400 |
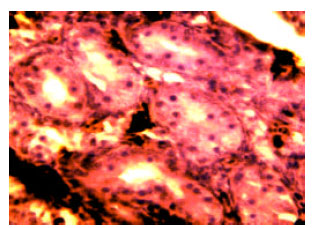 | |
| Fig. 11: | Morphologic character of 6 dat chicken testis organ. (Arrow is spermatogonia) x200 |
At this time, some lumens could be seen dimly in the seminiferous tubule but were not typical. There were 2 or 3 layers of spermatogenous cells distributed as cocenric circle in the tubal wall of seminiferous tubule. The amount of mesenchymal cells was increased and the number of group was increased too, though, as a whole, they were scattered sparsely.
The age of 6 days. Paraffin section of testes from 6 days chicken were observed by light microscope. At this time, spermatogenous cells in testes began to increase. There were about 3-5 spermatogenous cells in each transaction, which volumes were smaller than gonocytes. Cell nuclei were round and ellipse (Fig. 11). The morphologic characteristics were coincidence with the appearance of HE staining.
DISCUSSIONS
Didymus-genesis: There were two theories about didymus-genesis at present. First version considered that dydimus was originated from mesonephros blastema. Another argued that didymus was constituted of spermatogonium and thickening somatic cavity epithelium. Rodemer et al. (1986) thought that mesoderm gradually developed to form mesonephros from 16 pair somites stage to 23 pair soaties stages. Then day 6th-7th, gonad differentiated and mesonephros began to participate into didymus-genesis. But, in the present experiment, we found cavity epithelium thickened from somatic cavity corn to future gonad region at 3rd and most of PGCs existed among thickening somatic cavity epithelium. Following thickening somatic cavity epithelium moved in direction to mesoephros, PGCs located region also changed. All these demonstrated that didymus was sourced from thickening somatic cavity epithelium and supported second version.
Origin, development and differentiation of spermatogonium: Ukeshima et al. (1987) pointed out that PGCs arrived at small blood vessel near the germ ridge by blood cycling on 2rd-3rd hatching day and then PGCs intruded into thickened somatic cavity epithelium following permeating somatic epithelium of lamina visceralis when epithelium of future gonad region, which first formed gonadial morphous. In our experiments, based on migration and ecesis position of PGCs, we estimated that chick spermatogonium was not originated from gonad but soused from area pellucida of early blast, firstly existing in form of primordial germ cells and then migrated into developing gonad by embryo cycling system and PGCs began to profile after sex differentiation, meanwhile glycogen particle in PGCs plasma decomposed which suggested germ cells differentiation would initiate. On 7th hatching day, glycogen particle in PGCs plasma disappeared, gonad appeared testicual characterization in the male. On 8th hatching day, convoluted seminiferous tubule cord commenced to form, demonstrating solid center morphous and germ cells situated among them. Afterward, germ cells sequencely mitosed and cells number increased unceasing. A part of them renewed into stem cells stored in seminiferous tubule. Others entered growth phase and became primary spermatocyte (2n).
Spermatogonium migration and gonad: Spermatogonium originated from outside the gonad but finally arrived at gonad. During the process, for what reason made germ cells migrate in direction of gonad, there were two explanations that were chemotaxis effect and mechanical effect. Dantschakaff (1931) deemed that the mechanical effects that located PGCs at gonad probably were smaller antilinear of visceral mesoderm blood capillary under gonad proximal wall, which made blood cycle deceleration, retarded PGCs movement and caused PGCs initiative leave blood vessel into gonad region. This mechanical process followed by slow blood cycle and low blood pressure. Goldin et al. (1990) reported that PGCs migration was caused by a long-distance chemotaxis effect. Mouse germ ridge in vitro culture results showed that germ ridge had attraction to PGCs and the effect was probably relative to tissue growth factor TGFβ1. PGCs maybe had TGFβ1 receptors response to a gradient signal, then reorganized cell skeleton to polarize and migrate along the signal concentration gradient. When anti-TGFβ1 antibody was added into conditioned medium of germ cells, even very low concentration (1/1500), attraction effect of germ ridge could be blocked completely. Someone estimated that germ ridge could secrete TGFβ1 to stimulate peripheral cells externalizating extracellular matrix molecules. Furthermore, it was confirmed that TGFβ could stimulate fibronectin (FN), other extracellular matrix and their receptors secretions.
ACKNOWLEDGMENT
We are grateful to the National Natural Science Foundation of China (Project No. 30671509).
REFERENCES
- Golden, I., C. Wylie and J. Heasman, 1990. Genital ridges exert long-range effects on mouse primordial germ cell numbers and direction of migration in culture. Development, 103: 357-363.
Direct Link - Zaceanti, F., M. Yallisneri and Quaglia, 1990. Early aspects of sex differentiation in the gonads of chick embryos. Differe, 43: 71-80.
Direct Link