
Noura El-Ahmady El-Naggar
Department of Bioprocess Development, Genetic Engineering and Biotechnology Research Institute, City of Scientific Research and Technological Applications, Alexandria, Egypt
Mervat H. Hussein
Department of Botany, Faculty of Science, Mansoura University, Mansoura, Egypt
Sami A. Shaaban-Dessuuki
Department of Botany, Faculty of Science, Mansoura University, Mansoura, Egypt
Shimaa R. Dalal
Department of Botany, Faculty of Science, Mansoura University, Mansoura, Egypt
Journal of Applied Sciences
Year: 2016 | Volume: 16 | Issue: 9 | Page No.: 429-437







ABSTRACT
Background: Due to high nutritional value, natural pigments and antioxidant activity, Chlorella vulgaris is used as a health food, feed supplement, as well as in the pharmaceutical and cosmetics industry. It contains polysaccharides, proteins, lipids, antioxidants, vitamins, carotenoids, immunostimulator compounds and minerals. A statistical approach has been employed for which a Plackett-Burman design is used for identifying significant variables influencing polysaccharides production by Chlorella vulgaris. Materials and Methods: Ten different independent variables including mannitol, sodium acetate, potassium citrate, NaNO3, K2HPO4, MgSO4, CaCl2.2H2O, Na2CO3, inoculum size and inoculum age were screened by Plackett-Burman experiment and High Performance Liquid Chromatography (HPLC) was used to characterize the compounds in Chlorella vulgaris polysaccharides. Results: Among the variables screened, mannitol, sodium acetate and inoculum age had significant effects on polysaccharides production. The HPLC chromatograms indicated that Chlorella vulgaris extract contain fructose, glucose, maltose and lactose with concentration of 13.259, 283.027, 26.387 and 25.489 mg g–1 dry weight, respectively. Conclusion: The results indicate that this alga contains high amount of glucose which can use in different applications.
PDF Abstract XML References Citation
Received: May 24, 2016;
Accepted: July 15, 2016;
Published: April 29, 2019
How to cite this article
Noura El-Ahmady El-Naggar, Mervat H. Hussein, Sami A. Shaaban-Dessuuki and Shimaa R. Dalal, 2016. Rapid Screening of Various Factors Influencing Polysaccharides Production by Microchlorophyte Alga, Chlorella vulgaris, by Application of Plackett-Burman Design: Characterization of Polysaccharides Using High Performance Liquid Chromatography. Journal of Applied Sciences, 16: 429-437.
DOI: 10.3923/jas.2016.429.437
URL: https://scialert.net/abstract/?doi=jas.2016.429.437
DOI: 10.3923/jas.2016.429.437
URL: https://scialert.net/abstract/?doi=jas.2016.429.437
INTRODUCTION
Microalgae are considered as some of the most economically promising organism in our world1. They have drown increased attention from researchers in recent years because they provide lipids for biofuel production, as well as numerous high-value bioproducts such as omega-3 fatty acids2, health food, carotenoids and animal feed3. Chlorella is the most cultivated eukaryotic green microalga, as it is widely used as a health food and feed supplement, as well as in the pharmaceutical and cosmetics industry4. Chlorella vulgaris contains 45% protein (w/w, dry basis), 20% fat, 20% carbohydrates, 5% fiber and 10% minerals and vitamins. The growth of algae is a function of many factors, including nutrients, pH, salinity, temperature and light5.
Polysaccharides are biopolymers of simple sugars (monosaccharides) linked together by glycosidic bonds and they have numerous commercial applications in products such as stabilisers, thickeners, emulsifiers, food, feed and beverages etc.6,7. Algal polysaccharides posses biological activities as antitumor, antivirus, antihyperlipidemia and anticoagulant have been found to be associated with polysaccharides in some species of seaweed which have been purified and developed as a new generation of drugs8,9. Studies are widely conducted on isolation, purification and structural analysis of polysaccharides10, 11. Several reports have indicated that chemical components and molecular weight of seaweed polysaccharides are among the major important factors responsible for biological activities12.
A statistical approach has been employed in the present study for which a Plackett-Burman design is used for identifying significant variables influencing polysaccharides production by freshwater algae Chlorella vulgaris. Also high performance liquid chromatography was used to characterize the compounds in Chlorella vulgaris polysaccharides.
MATERIALS AND METHODS
Algal strain isolation, identification and culture conditions: Fresh water Chlorella vulgaris was isolated from water sample from river Nile. Culture purification was according to Andersen13 and the alga was identified according to Philipose14. Chlorella vulgaris was grown in axenic cultures at 25±2°C under continuous illumination 3600 lux in 500 mL conical flasks, containing 200 mL BG11 medium15 for 12 days incubation period.
Extraction of polysaccharides: The hot water method of Wang et al.16 at 70°C for 4 h was followed.
Determination of polysaccharides: The polysaccharide was determined by the phenol-sulfuric acid reaction using glucose as standard17,18.
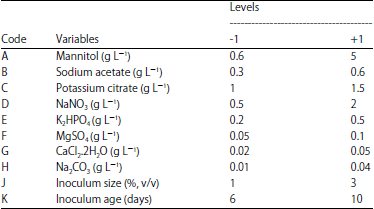
Screening of main factors influences polysaccharides production by Plackett-Burman design: In order to determine the major factors affecting polysaccharides production, different variables were evaluated using Plackett-Burman statistical experimental design which is a 2 factorial design that identifies the critical variables required for elevated polysaccharides production and is very useful for screening the most important factors with respect to their main effects19,20. Plackett-Burman statistical design is a well-established and suitable for complicated systems with multiple variables to screen out and select most significant environmental and nutritional variables. Compared with other medium design strategies, the Plackett-Burman design is simple, fast method for screening large number of variables in one experiment to see which gives the best results and is often used to evaluate the important variables affecting culture requirements for polysaccharides production. The total number of experiments to be carried out according to Plackett-Burman is n+1 where, n is the number of variables21. A total of 10 independent (assigned) and one unassigned variables (commonly referred as dummy variable) were screened in Plackett-Burman experimental design. Dummy variables (D) are used to estimate experimental errors in data analysis. Table 1 shows the 11 different independent variables including mannitol, sodium acetate, potassium citrate, NaNO3, K2HPO4, MgSO4, CaCl2.2H2O, Na2CO3, inoculum size and inoculum age which chosen to be screened by Plackett-Burman experiment. Each variable is represented at 2 levels high and low denoted by (+) and (-), respectively. The experiment was conducted in 12 runs to study the effect of the selected variables on the production of polysaccharides.
| Table 1: | Experimental independent variables at 2 levels used for the production of polysaccharides by Chlorella vulgaris using Plackett-Burman design |
 | |
All trials were performed in duplicate and the average of polysaccharides production were treated as responses. Plackett-Burman experimental design is based on the first order model:
| (1) |
where, Y is the response or dependent variable (polysaccharides production), it will always be the variable we aim to predict, β0 is the model intercept and βi is the linear coefficient and Xi is the level of the independent variable, it is the variable that will help us explain polysaccharides production.
This model does not describe interaction among factors and it is used to screen and evaluate the important factors that influence the response.
Statistical analysis: Minitab 14.0 (Minitab Inc., Pennsylvania, USA) was used for the experimental designs. Design Expert® 7.0 software version 7 (Stat-ease Inc., USA) for windows was used for the statistical analysis for the obtained experimental data. The p-values were used as a tool to check the significance of the interaction effects, which in turn may indicate the patterns of the interactions among the variables22. The quality of fit of regression model was expressed via the correlation coefficient (R), the coefficient of determination (R2) and the adjusted R2 and its statistical significance was determined by an F-test.
Hydrolysis of the algal polysaccharide: Algal polysaccharides were hydrolyzed in 2 mol L–1 trifluoroacetic acid for 10 h at 100°C in a sealed glass tube under nitrogen flow. The residual acid was removed under a stream of N2 in a 40-45°C water-bath after adding methanol and taking to dryness 3 times and then distilled water (1 mL) was added to dissolve the residue.
High Performance Liquid Chromatography (HPLC)- refractive index detector: Monosaccharides analysis in the extracts of the decomposed alga was performed with HPLC (Agilent 1100 HPLC system) using an Hypersil ASP-2 column (4.6×250 mm) with a mobile phase of acetonitrile-water (80:20) a flow rate of 1 mL min–1. The temperature of column was set at 40°C. The injection volume of mixed monosaccharide standards and samples hydrolyzates was 10 μL of the final extract.
Standards and quantification of monosaccharides: Identification of the monosaccharides in the hydrolyzates of samples were carried out by comparing their retention times obtained for the authentic standards for six monosaccharides (xylose, fructose, glucose, sucrose, maltose and lactose) in mobile phase which analyzed individually and mixed in concentration of 10 mg mL–1 under the same HPLC conditions.
RESULTS AND DISCUSSION
Statistical screening of various factors influencing extracellular polysaccharides production by Chlorella vulgaris using Plackett-Burman design: The influence of 11 medium factors and conditions namely mannitol, sodium acetate, potassium citrate, NaNO3, K2HPO4, MgSO4, CaCl2.2H2O, Na2CO3, inoculum size and inoculum age was investigated in 12 runs using Plackett-Burman design. The Plackett-Burman design for the 12 runs and the corresponding response for polysaccharides production are shown in Table 2. The polysaccharides production varied markedly with the variables tested in range of 2.303-84.040 in the 12 trials, this variation reflects the importance of medium optimization to attain higher polysaccharides production. The maximum polysaccharides production (84.040 mg g–1 cell dry weight) was achieved in the run number 8, when minimal levels of mannitol, NaNO3, MgSO4, CaCl2.2H2O, Na2CO3 and maximal levels of sodium acetate, potassium citrate, K2HPO4, inoculum size and inoculum age were used while the lowest polysaccharides production (2.303 mg g–1 cell dry weight) was observed in the run number 7 when NaNO3, K2HPO4, MgSO4, Na2CO3, inoculum size were adjusted to the highest levels and mannitol, sodium acetate, potassium citrate, CaCl2.2H2O and inoculum age were adjusted to the lowest levels.
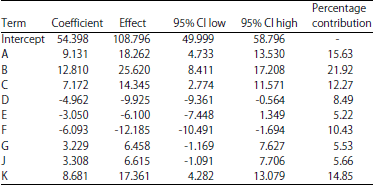
Statistical analysis of Plackett-Burman design for polysaccharides production by Chlorella vulgaris: Plackett-Burman design was used to determine which variables significantly affect polysaccharides production by Chlorella vulgaris. The relationship between a set of independent variables and the response (Y) is determined by a mathematical model called multiple-regression model. Statistical analysis of polysaccharides production was performed which is represented in Table 3 and 4. Table 3 and Fig. 1 shows the main effect of each variable on the polysaccharides production which estimated based on the difference between the average measurement calculated at the higher (+1) and lower (-1) levels of the constituent.
| Table 2: | Twelve trial Plackett-Burman experimental design for evaluation of independent variables with coded values along with the observed polysaccharides production |
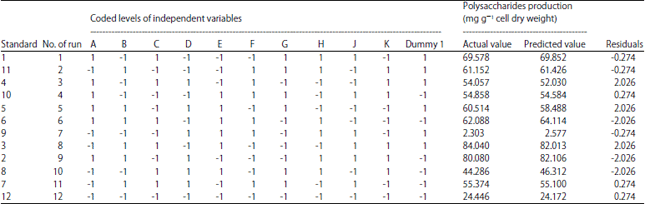 | |
| “-1": Correspond to the minimum value and "+1": Correspond to the maximum value of the input parameter range | |
| Table 3: | Regression coefficients, estimated effect and percentage of contribution for polysaccharides production by Chlorella vulgaris using Plackett-Burman design |
 | |
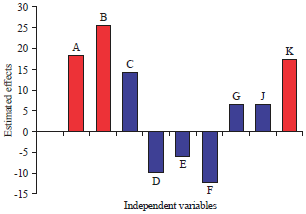 | |
| Fig. 1: | Effect of independent variables on polysaccharides production by Chlorella vulgaris using Plackett-Burman design (Red color represent the most significant positive independent variables affecting enzyme production) |
A large main effect either positive or negative indicates that a factor has a large impact on polysaccharides production, while an effect close to zero means that a factor has little or no effect.
| Table 4: | Regression statistics and analysis of variance (ANOVA) for the experimental results of Plackett-Burman design used for polysaccharides production by Chlorella vulgaris |
 | |
*Significant values, SS: Sum of squares, MS: Mean square, F: Fishers’s function, p-value: Level of significance, PRESS: Predicted residual sum of squares, CV%: Coefficient of variation% and df: Degree of freedom | |
The percentages contributions of the variables are given in Table 3. The results revealed that mannitol, sodium acetate and inoculum age are the most contributing components with 15.63, 21.92 and 14.85%, respectively.
The effects of the examined variables on polysaccharides production using the Plackett-Burman design were calculated and summarized in Table 3. The data revealed that, Na2CO3 (H) is insignificant variables with lower effect (-2.3) and lower percent of contribution (1.930). Lower percentage of contribution indicated higher p-value (0.414). Thus instead of starting with the maximum model effects, backward regression at alpha 0.15 was applied to remove/or eliminate the small effects of Na2CO3 (H). Then, the model fitted for the test of significance. Actually, the removal procedure ends when the model was at significant level.
Pareto chart allows detecting the order and significant effects of variables affecting polysaccharides production in Plackett-Burman experimental design. It displays the absolute values of the effects and draws a reference line on the chart. Any effect that extends past this reference line is potentially important. Pareto chart in design expert version 7.0 reproduce the relation between t-value (effect) vs., ranks. Pareto chart (Fig. 2) showed that sodium acetate (B) was the most significant variable affecting polysaccharides production followed by mannitol (A) and inoculum age (K).
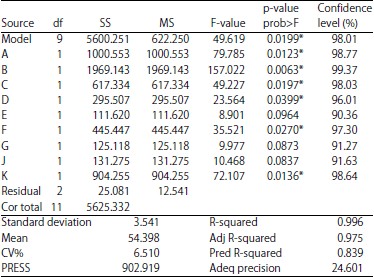
The analysis of variance (ANOVA) of the experimental design was calculated and the sum of square, mean square, F-value, t-value, p-value and confidence level are given in Table 4. The significance of the model was calculated by the p-value. The p-value is the probability which serves as a tool for checking the significance of each of the parameter. The model F-value of 49.62 implies that the model is significant. There is only a 1.99% chance that a "Model F-value" this large could occur due to noise. Values of "Prob>F" (p-value) less than 0.05 indicate model terms are significant. In this case A, B, C, D, F and K are significant model terms. Values greater than 0.1 indicate the model terms are not significant. If there are many insignificant model terms.
The data revealed that, 9 variables (mannitol, sodium acetate, potassium citrate, NaNO3, MgSO4 and inoculum age) were found to significantly affect polysaccharides production while the remaining five variables (K2HPO4, CaCl2.2H2O, Na2CO3 and inoculum size) have not significant influence on the polysaccharides production (Table 4). In this connection the analysis showed that, sodium acetate (B) with a probability value of 0.0063 was determined to be the most significant factor affecting polysaccharides production by Chlorella vulgaris at 99.37% confidence followed by mannitol (A) (p-value = 0.0123), inoculum age (K) (p-value = 98.64), the lower probability values indicate the more significant factors affecting polysaccharides production. Also, it was clear that the three variables exerted a positive effect on polysaccharides production which means that the increase in the concentrations of mannitol, sodium acetate and inoculum age could exert positive effect on polysaccharides production.
The R2 values provide a measure of how much variability in the observed response values can be explained by the experimental factors. The R2 value is always between 0 and 1. When R2 is closer to the 1, the stronger the model is and the better it predicts the response23. The value of the determination coefficient (R2) was found to be 0.996 indicates that 99.6% of the variability in the polysaccharides production could be explained by the model independent variables and only 0.4% of the total variations are not explained by the independent variables.
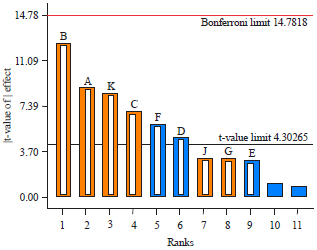 | |
| Fig. 2: | Pareto chart illustrates the order and significance of the variables affecting polysaccharides production by Chlorella vulgaris using Plackett-Burman design (blue colors represent negative effects and orange color represent positive effects) |
The adjusted R2 (0.9755) is also very high that indicates that the model is very significant24. The "Pred R-squared" of 0.8395 is in reasonable agreement with the "Adj R-squared" of 0.9755. This indicated a good adjustment between the observed and predicted values. "Adeq precision" measures the signal to noise ratio. A ratio greater than 4 is desirable. The ratio of 24.601 indicates an adequate signal.
The coefficient of variation percentage (CV%) is a measure of residual variation of the data relative to the size of the mean. Usually, the higher the value of CV, the lower is the reliability of experiment. Here a lower value of CV (6.510%) indicates a greater reliability of the experimental performance. The predicted residual sum of squares (PRESS) is a measure of how well the model fits each point in the design. The smaller the PRESS statistic, the better the model fits the data points. The value of PRESS is 902.919. The model shows standard deviation and mean value of 3.541 and 54.398, respectively (Table 4).
To approach the neighborhood of the optimum polysaccharides production, the coefficients of regression equation were calculated using design expert version 0.7 and the data (Table 4) was fitted to a first order polynomial equation. By neglecting the insignificant terms, the following regression equation in terms of coded factors was obtained:
(2) |
where, Y is the response (polysaccharides production) and A, B, C, D, F and K are mannitol, sodium acetate, potassium citrate, NaNO3, MgSO4 and inoculum age, respectively. The coefficient of each variable represents the effect extent of these variables on polysaccharides production.
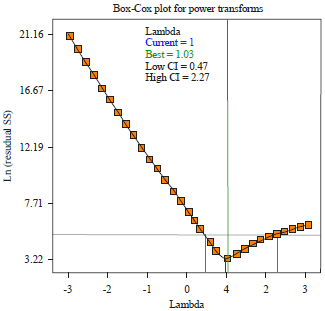 | |
| Fig. 3: | Box-Cox plot of model transformations |
Model adequacy checking: Box-Cox plot provides guideline for selecting the correct power law transformation to further improve the model. As observed from Fig. 3, the blue line indicates the current transformation (Lambda = 1) and the green line indicates the best lambda value (Lambda = 1), while the red lines indicate the minimum and maximum 95% confidence interval values (0.47 and 2.27, respectively). Therefore, the model needs no transformation, as current value of confidence interval (λ = 1) is very close to model design value (best = 1.03) and the model is in the optimal zone since the blue line falls within the red lines. So that the model is well fit to the experimental data obtained and well satisfies the assumptions of the analysis of variance.
The normal probability plot of the residuals is an important diagnostic tool to detect and explain the systematic departures from the normality22. Figure 4 shows a plot of normal probability of the experimental results. The normal probability plot is a graphical technique for assessing whether or not a data set is approximately normally distributed. The residual was plotted against a theoretical normal distribution of the model in such a way that the points should form an approximate straight line for polysaccharides production. Departures from this straight line indicate departures from normality. A linear pattern demonstrated that the errors are normally distributed and are independent of each other. The normal probability plot of the residuals shows the points close to a diagonal line; therefore, the residuals appear to be approximately normally distributed. This indicates that the model was well fitted with the experimental results.
Many algal organisms are capable of using either metabolism process (autotrophic or heterotrophic) for growth, meaning that they are able to photosynthesize as well as ingest prey or organic materials25.
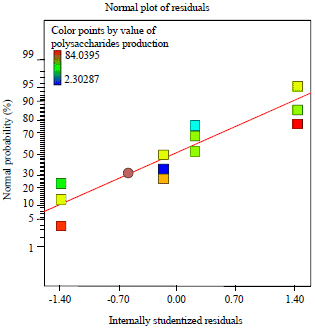 | |
| Fig. 4: | Normal probability plot of residuals to check for normality of residuals for polysaccharides production by Chlorella vulgaris determined by the first order polynomial equation |
Although, most microalgae are photoautotrophs, some microalgae can use organic carbon substances as the sources of energy and carbon for cell growth. Mixotrophy is growth in which organic carbon is assimilated in the light simultaneously with carbon dioxide fixation. The ability of microalgae to use organic carbon as an energy source is important because it can minimize the inhibitory effects of seasonal and diurnal light limitation on growth in outdoor cultures. A considerable number of algae, for example Chlorella26 can grow mixotrophically and heterotrophically in the presence of organic matter such as carbohydrates and acetate. Mixotrophic culture of C. vulgaris integrates some advantages of heterotrophic and phototrophic configurations27, while overcoming the difficulties associated with these two types of cultures28. The benefits are associated with a significant increase in growth rate and productivity due to high organic carbon incorporation with in cell, particularly in the formation of metabolites such as lipids, polysaccharides and proteins29. This is particularly important, as increases of up to 5 times have been reported compared with autotrophic cultures of microalgae30. Another advantage of mixotrophic cultures is the use of light, which induces and regulates the production of high value macromolecules such as pigments31 and fatty acids32.
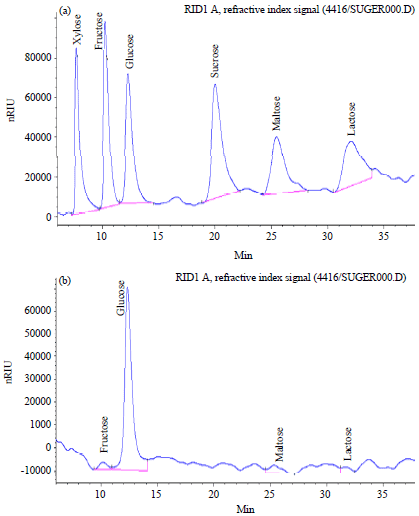 | |
| Fig. 5(a-b): | (a) HPLC chromatograms of standard monosaccharides (Xylose, fructose, glucose, sucrose, maltose and lactose), and (b) HPLC chromatograms of the extract of Chlorella vulgaris, x-axis: Retention times (min), y-axis: Observed peak area (nRIU: Intensity of refraction index signals) |
It is true that acetate can be used as a carbon source via acetyl CoA, which can then be changed to pyruvate and further, oxidised in the metabolic pathway. Sodium acetate could promote both growth and lipid accumulation in many microalgae such as Phaeodactylum tricornutum33, Brachiomonas submarina34 and Chlorella vulgaris35, while Scenedesmus sp. AARL G02236 seemed unaffected by sodium acetate supplement.
Yee37was found that sodium acetate and sugar alcohol mannitol were potential carbon sources for mixotrophic culture growth enhancement of M. griffithii. Supplementation of culture medium with these carbon sources resulted in approximately 1-1.5 fold increase in final cell density compared to control in a small scale culture. In a larger scale mixotrophic culture with aeration, 0.05% mannitol resulted in a decent 1-1.5 fold increase in final cell density, approximately 2 fold increase in growth rate. In terms of biomass, mannitol resulted in approximately 0.5 fold increase compared to control. Supplementation of culture medium with 0.05% mannitol resulted in a final dry biomass weight of 497.4 mg L–1 which is higher than for Chlorella sp. The Y-8 supplemented with 0.1% fructose, which produced approximately 300 mg L–1 dry biomass38. A slight decrease in total oil content was observed in cells grown in mannitol (11.87%) compared to control (13.8%)38.
It was also found that the age of a parent culture used as a source of inoculum may exert a very pronounced effect on the rates of photosynthesis and respiration in cells of the daughter-cultures for several days, the age of inoculum is important to achieve optimum growth of algae. Inoculum age of 10 days was found to be the most suitable conditions for maximum production of polysaccharides production by Chlorella vulgaris may be because cells are in the logarithmic or early-exponential growth phase and the cells are more active under the used growth conditions.
High Performance Liquid Chromatography (HPLC): The HPLC chromatograms (Fig. 5) indicated that Chlorella vulgaris extract contain fructose, glucose, maltose and lactose with concentration of 13.259, 283.027, 26.387 and 25.489 mg g–1 dry weight, respectively (Table 5).
| Table 5: | Composition of the polysaccharides isolated from extract of Chlorella vulgaris (mg g–1 sample) |
 | |
The results indicate that this alga contains high amount of glucose which can use in different applications such as bioethanol production. Polysaccharides in general and sulphated exopolysaccharides in particular are released by many species of microalgae for example, Chlorella stigmatophora the main component is glucose and xylose39 and contain sulphated polysaccharide type, Chlorella spp., contain soluble polysaccharide40. The polysaccharides from microalgae serve as antiviral agents, health foods, antioxidants, they have anti-inflammatory properties and have a role in the immunomodulatory system and they may also be used as lubricants for bone joints or even as drag-reducing substances for ships41.
CONCLUSION
In this study, polysaccharides production by microalgae Chlorella vulgaris was investigated. A Plackett-Burman design is used for identifying significant variables influencing polysaccharides production process by microalgae Chlorella vulgaris. The results showed that among the variables screened, mannitol, sodium acetate and inoculum age had significant effects on polysaccharides production. Also high performance liquid chromatography was used to characterize the compounds in Chlorella vulgaris polysaccharides which indicated that Chlorella vulgaris extract contain fructose, glucose, maltose and lactose with concentration of 13.259, 283.027, 26.387 and 25.489 mg g–1 dry weight, respectively.
ACKNOWLEDGMENT
The authors gratefully acknowledge the Department of Botany, Faculty of Science, Mansoura University, Mansoura, Egypt, for their financial support.
REFERENCES
- Pittman, J.K., A.P. Dean and O. Osundeko, 2011. The potential of sustainable algal biofuel production using wastewater resources. Bioresour. Technol., 102: 17-25.
CrossRefDirect Link - Larsdotter, K., 2006. Wastewater treatment with microalgae-a literature review. Vatten, 62: 31-38.
Direct Link - Sharma, R., G.P. Singh and V.K. Sharma, 2012. Effects of culture conditions on growth and biochemical profile of Chlorella vulgaris. J. Plant Pathol. Microbiol., Vol. 3.
CrossRefDirect Link - Belasco, W., 1997. Algae burgers for a hungry world? The rise and fall of chlorella cuisine. Technol. Cult., 38: 608-634.
Direct Link - Tseng, C.K., 2001. Algal biotechnology industries and research activities in China. J. Applied Phycol., 13: 375-380.
CrossRefDirect Link - Bixler, H.J. and H. Porse, 2010. A decade of change in the seaweed hydrocolloids industry. J. Applied Phycol., 23: 321-335.
CrossRefDirect Link - Schaeffer, D.J. and V.S. Krylov, 2000. Anti-HIV activity of extracts and compounds from algae and cyanobacteria. Ecotoxicol. Environ. Safety, 45: 208-227.
CrossRefPubMedDirect Link - Trento, F., F. Cattaneo, R. Pescador, R. Porta and L. Ferro, 2001. Antithrombin activity of an algal polysaccharide. Thrombosis Res., 102: 457-465.
CrossRefDirect Link - Girod, S., P. Baldet-Dupy, H. Maillols and J.M. Devoisselle, 2002. On-line direct determination of the second virial coefficient of a natural polysaccharide using size-exclusion chromatography and multi-angle laser light scattering. J. Chromatogr. A, 943: 147-152.
CrossRefDirect Link - Maksimova, I.V., L.B. Bratkovskaia and S.E. Plekhanov, 2004. [Extracellular carbohydrates and polysaccharides of the algae Chlorella pyrenoidosa Chick S-39]. Izv Akad. Nauk. Ser. Biol., 2: 217-224, (In Russian).
PubMedDirect Link - Wang, Y., M. Zhang, D. Ruan, A.S. Shashkov, M. Kilcoyne, A.V. Savage and L. Zhang, 2004. Chemical components and molecular mass of six polysaccharides isolated from the sclerotium of Poria cocos. Carbohydr. Res., 339: 327-334.
CrossRefDirect Link - Rippka, R., J. Deruelles, J.B. Waterbury, M. Herdman and R.Y. Stanier, 1979. Generic assignments, strain histories and properties of pure cultures of cyanobacteria. J. Gen. Microbiol., 111: 1-61.
CrossRefDirect Link - Wang, H., Q. Wang, S.C. Wang, Z.T. Wang and J.F. Shen, 2006. [Extraction, isolation and structure identification of polysaccharide in root of Salvia miltiorrhiza]. China J. Chin. Mater. Med., 31: 1075-1077.
PubMedDirect Link - DuBois, M., K.A. Gilles, J.K. Hamilton, P.A. Rebers and F. Smith, 1956. Colorimetric method for determination of sugars and related substances. Anal. Chem., 28: 350-356.
CrossRefDirect Link - Cuesta, G., N. Suarez, M.I. Bessio, F. Ferreira and H. Massaldi, 2003. Quantitative determination of pneumococcal capsular polysaccharide serotype 14 using a modification of phenol-sulfuric acid method. J. Microbiol. Meth., 52: 69-73.
CrossRefDirect Link - Krishnan, S., S.G. Prapulla, D. Rajalakshmi, M.C. Misra and N.G. Karanth, 1998. Screening and selection of media components for lactic acid production using Plackett-Burman design. Bioprocess Eng., 19: 61-65.
CrossRef - El-Naggar, N.E.A., 2015. Extracellular production of the oncolytic enzyme, L-asparaginase, by newly isolated Streptomyces sp. strain NEAE-95 as potential microbial cell factories: Optimization of culture conditions using response surface methodology. Curr. Pharm. Biotechnol., 16: 162-178.
Direct Link - Plackett, R.L. and J.P. Burman, 1946. The design of optimum multifactorial experiments. Biometrika, 33: 305-325.
CrossRefDirect Link - Kaushik, R., S. Saran, J. Isar and R.K. Saxena, 2006. Statistical optimization of medium components and growth conditions by response surface methodology to enhance lipase production by Aspergillus carneus. J. Mol. Catal. B: Enzym., 40: 121-126.
CrossRefDirect Link - Zhang, X.W., Y.M. Zhang and F. Chen, 1999. Application of mathematical models to the determination optimal glucose concentration and light intensity for mixotrophic culture of Spirulina platensis. Process Biochem., 34: 477-481.
CrossRefDirect Link - Endo, H., H. Sansawa and K. Nakajima, 1977. Studies on Chlorella regularis, heterotrophic fast-growing strain II. Mixotrophic growth in relation to light intensity and acetate concentration. Plant Cell Physiol., 18: 199-205.
Direct Link - Lee, Y.K., 2001. Microalgal mass culture systems and methods: Their limitation and potential. J. Applied Phycol., 13: 307-315.
CrossRefDirect Link - Lee, Y.K., S.Y. Ding, C.H. Hoe and C.S. Low, 1996. Mixotrophic growth of Chlorella sorokiniana in outdoor enclosed photobioreactor. J. Applied Phycol., 8: 163-169.
CrossRefDirect Link - Syrett, P.J., S.M. Bocks and M.J. Merrett, 1964. The assimilation of acetate by Chlorella vulgaris. J. Exp. Bot., 15: 35-47.
CrossRefDirect Link - Li, X., H. Xu and Q. Wu, 2007. Large-scale biodiesel production from microalga Chlorella protothecoides through heterotrophic cultivation in bioreactors. Biotechnol. Bioeng., 98: 764-771.
CrossRefDirect Link - Wang, H., R. Fu and G. Pei, 2012. A study on lipid production of the mixotrophic microalgae Phaeodactylum tricornutum on various carbon sources. Afr. J. Microbiol. Res., 6: 1041-1047.
Direct Link - Yang, X., X. Zeng, Y. Ji and Q. Liu, 2003. Effects of sodium nitrate and sodium acetate concentrations on the growth and fatty acid composition of Brachiomonas submarina. J. Ocean Univ. Qingdao, 2: 75-78.
Direct Link - Heredia-Arroyo, T., W. Wei, R. Ruan and B. Hu, 2011. Mixotrophic cultivation of Chlorella vulgaris and its potential application for the oil accumulation from non-sugar materials. Biomass Bioenergy, 35: 2245-2253.
CrossRefDirect Link - Dittamart, D., C. Pumas, J. Pekkoh and Y. Peerapornpisal, 2014. Effects of organic carbon source and light-dark period on growth and lipid accumulation of Scenedesmus sp. AARL G022. Maejo Int. J. Sci. Technol., 8: 198-206.
Direct Link - Yee, W., 2015. Feasibility of various carbon sources and plant materials in enhancing the growth and biomass productivity of the freshwater microalgae Monoraphidium griffithii NS16. Bioresour. Technol., 196: 1-8.
CrossRefPubMedDirect Link - Lin, T.S. and J.Y. Wu, 2015. Effect of carbon sources on growth and lipid accumulation of newly isolated microalgae cultured under mixotrophic condition. Bioresour. Technol., 184: 100-107.
CrossRefDirect Link - Guzman, S., A. Gato, M. Lamela, M. Freire-Garabal and J.M. Calleja, 2003. Anti-inflammatory and immunomodulatory activities of polysaccharide from Chlorella stigmatophora and Phaeodactylum tricornutum. Phytother. Res., 17: 665-670.
CrossRefPubMedDirect Link - Sun, Y. and C. Wang, 2009. The optimal growth conditions for the biomass production of Isochrysis galbana and the effects that phosphorus, Zn2+, CO2 and light intensity have on the biochemical composition of Isochrysis galbana and the activity of extracellular CA. Biotechnol. Bioprocess Eng., 14: 225-231.
CrossRefDirect Link - Raposo, M.F.D.J., R.M.S.C. de Morais and A.M.M.B. de Morais, 2013. Bioactivity and applications of sulphated polysaccharides from marine microalgae. Mar. Drugs, 11: 233-252.
CrossRefDirect Link