
Kek Heng Chua
Department of Molecular Medicine,Faculty of Medicine, University of Malaya, 50603 Kuala Lumpur, Malaysia
Hwa Chia Chai
Department of Molecular Medicine,Faculty of Medicine, University of Malaya, 50603 Kuala Lumpur, Malaysia
S.D. Puthucheary
Department of Medical Microbiology, Faculty of Medicine, University of Malaya, 50603 Kuala Lumpur, Malaysia
Journal of Applied Sciences
Year: 2008 | Volume: 8 | Issue: 21 | Page No.: 3999-4003







ABSTRACT
This study reportsa sequential biopanning against serum obtained from a patient infected with B. pseudomallei using a phage display peptide library. A peptide bearing the consensus sequence NKNSFDAWLQSF was obtained after 3 rounds of biopanning and it partly resembles the sequence 35FDAWLAAQEF44 of B. pseudomallei site-specific recombinase, which is a phage integrase family protein. Overall, the mimotope obtained in this study could be a potential candidate for the development of a cost effective diagnostic test or vaccine in the future.
PDF Abstract XML References Citation
How to cite this article
Kek Heng Chua, Hwa Chia Chai and S.D. Puthucheary, 2008. Identification of Burkholderia pseudomallei Mimotope using Phage Display Approach. Journal of Applied Sciences, 8: 3999-4003.
DOI: 10.3923/jas.2008.3999.4003
URL: https://scialert.net/abstract/?doi=jas.2008.3999.4003
DOI: 10.3923/jas.2008.3999.4003
URL: https://scialert.net/abstract/?doi=jas.2008.3999.4003
INTRODUCTION
Burkholderia pseudomallei is a soil saprophyte, which is geographically restricted in distribution to tropical and subtropical Australia and Southeast Asia. This organism is widely disseminated in soil, streams, ponds and rice paddies. Surveys conducted on the soil and surface water of Sabah and West Malaysia for B. pseudomallei showed that the organism is widely distributed in Malaysia. The highest isolation rate of 14.6% was in surface water from wet rice fields (Babjee and Aidah, 1994). In addition, the most striking finding in the soil sampling was the high isolation rate of 27.8% of B. pseudomallei from cleared fields. These surveys suggest that nutritional requirements do not limit the geographical distribution of B. pseudomallei (Babjee and Aidah, 1994). B. pseudomallei causes melioidosis which exhibits several symptoms, including formation of skin abscesses, sepsis and septic shock, abscess formation in several internal organs and acute pulmonary disease (Forbes et al., 1998). Infection with B. pseudomallei may result in a subclinical infection and remain undetected for a number of years. Moreover, both the chronic and subclinical forms generally can remain undiagnosed until activated by a traumatic event or a decrease in immunocompetence (Reckseidler et al., 2001). A high mortality rate of 65% was reported in patients with the septicemic form of melioidosis, which is characterized by dissemination of the bacteria with multi-organ involvement and isolation of the bacteria from the blood and various organs (Puthucheary et al., 1992). In general, the pathogenesis of melioidosis has not been completely elucidated. A previous study has suggested that the surface component of B. pseudomallei might serve as a virulence factor and play a role in the attachment to the host cell surface (Brown et al., 2002).
Biopanning is one of the powerful approaches that are usually carried out with a phage display library. A phage display library is made up of many recombinant phages, each displaying a different peptide or protein as a fusion with coat protein on the surface of filamentous bacteriophages (Smith et al., 1998). Proteins or peptides that interact with the desired target such as antibodies, enzymes, DNA-binding proteins and cell-surface receptors are then screened for their epitopes using biopanning. This approach has been widely applied in the studies of antibodies (Shen et al., 2007; Jiao et al., 2005), cancers (Chen et al., 2007; Akita et al., 2006), drug discovery (Sergeeva et al., 2006) and mimotope or epitope mapping (Tungtrakanpoung et al., 2006). Therefore, in this study, we set out to possibly uncover and characterize the immunogenic surface epitope or mimotope of B. pseudomallei which might play an important role in the pathogenecity regarding host-cell attachment.
MATERIALS AND METHODS
Biopanning: The biopanning process was carried out using serum from a patient with melioidosis. E. coli ER2738 was prepared in 10 mL of Luria-Bertani broth and incubated at 37°C with shaking at 220 rpm. One hundred and fifty microlitres of patient serum in 0.1 M NaHCO3 (pH 8.6) was prepared and incubated overnight in a microtiter plate at 4°C. The solution was then discarded and blocking buffer (10% skim milk in PBS) was added and kept at 4°C for 1 h. After that, the wells were washed 5 times with PBS-T [PBS + 0.1% (v/v) Tween-20)]. A 5x1012 pfu of the phage library in 150 μL of PBS-T was dispensed into the wells and incubated for 1 h at room temperature and the mixture containing unbound phage was then discarded. The wells were washed 10 times with PBS-T followed by trypsinisation for 30 min. The elute containing the selected phage was transferred into a microfuge tube and stored at 4°C overnight for further use.
One microlitre of the phage elute was used for titration. The elute containing the selected phage was enriched by the addition of 10 mL of E. coli ER 2738 and incubated at 37°C for 4 h with vigorous shaking, then centrifuged at 13,000 rpm at 4°C for 10 min, followed by a second centrifugation. A 1/6 volume of PEG/NaCl was added to the supernatant in a new microfuge tube and allowed to precipitate overnight at 4°C. The precipitate was centrifuged at 13,000 rpm, 4°C for 15 min, the supernatant was discarded and the pellet was dissolved in 1 mL of PBS. This suspension was centrifuged at 13,000 rpm for 5 min to pellet residual cells and the supernatant was precipitated again with 1/6 volume of PEG/NaCl and incubated on ice for 60 min. Supernatant was discarded after centrifugation at 13,000 rpm for 10 min and the pellet containing the selected phage was suspended in 200 μL of PBS, 0.02% NaN3 and this was the amplified phage elute.
Phage titration: The amplified elute was titrated on LB/IPTG/Xgal plates. Phage titers corresponding to the plaque formed were determined and this was used to obtain an input volume corresponding to at least 1x1011 pfu. Then a second round of biopanning was carried out using the first round of amplified phage elute as the input phage. The amplified elute from the second round then became the input phage for a third round of biopanning.
Ten-fold serial dilutions of phage in LB were prepared. Two hundred microliters of the mid-log phase E. coli ER 2738 was dispensed into each microfuge tubes. Ten microlitres of each dilution was then aliquoted into separate tubes, mixed and incubated at room temperature for 5 min. Infected cells were transferred to culture tubes containing the 45°C agarose top and immediately poured and spread evenly on LB/IPTG/Xgal plates which were incubated overnight at 37°C and on the following day, the blue plaques were counted.
ELISA assay: Two wells of a microtiter plate were coated overnight at 4°C with 100 μL of 100 μg mL-1 melioidosis patient serum in 0.1 M NaHCO3 (pH 8.6) and 100 μg mL-1 Bovine Serum Albumin (BSA) (as negative control) in 0.1 M NaHCO3 (pH 8.6), respectively. The following day, excess target solution was discarded and each well was filled with blocking buffer and incubated at 4°C for 1 h. After that, the wells were washed 6 times with PBS-T and phage solution was added to the 2 coated wells which were incubated at 4°C for 1 h. The wells were then washed with PBS-T and 200 μL of diluted HRP-conjugated anti-M13 antibody (1:5000) (Pharmacia) was added to each well and incubated at room temperature for 1 h. The plate was washed with PBS-T and HRP substrate solution (0.02% (w/v) ABTS, 50 mM sodium citrate, 30% (v/v) H2O2) was added to each well for enzymatic reaction which led to colour development. Optical Density (OD) readings at 415 nm were obtained using a microplate reader (Multiskan MCC/340P).
Phagemid DNA extraction and sequencing: After the third round of biopanning, 50 μL of phage and 100 μL of the E. coli ER2738 were mixed together in 10 mL of LB broth and incubated at 37°C, with shaking at 220 rpm for 4 h. After that, the culture was spread onto a LB/IPTG/Xgal plate and incubated overnight at 37°C. Five single colonies were randomly selected from the plate and inoculated into separate 10 mL LB medium with 100 μL of E. coli ER2738. These cultures were incubated at 37°C for 4 h with shaking at 220 rpm. Following this, the culture-pellets were collected after centrifugation. Conventional alkaline lysis technique was used for phagemid extraction (Sambrook et al., 1989). Phagemid DNA was then dissolved in 50 μL of dH2O and ready for sequencing using the -96 gIII sequencing primer: 5`-CCCTCATAGTTAGCGTAACG-3`.
Database search and analysis: The nucleotide sequence obtained was then translated into deduced amino acid sequence and subjected to database search against B. pseudomallei proteins. Non-redundant GenBank CDS database was searched using NCBI Blast and the hit representing the highest score, sequence coverage and the lowest E-value were selected.
RESULTS
Biopanning: The agar plates with blue plaques containing 10-fold dilutions of phage titration (data not shown), represent the recombinant phage from the phage library that carries the lacZα gene. Table 1 shows the number of blue plaques counted. Consistent increase of plaque numbers from the first to the third round of biopanning showed that binding, between the peptide displayed by phages and the patient serum, had become more specific in subsequent rounds of biopanning.
 | |
| Fig. 1: | Sequencing result of clone binding to serum of patient with melioidosis (Sequence in bold is the peptide that potentially can bind to patient serum) |
| Table 1: | No. of plaques on LB/IPTG/Xgal plates after biopanning |
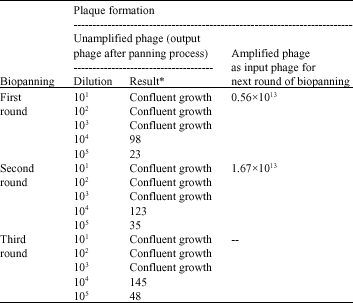 | |
| Input phage for first round of biopanning = 4x1011 pfu. *No. of blue plaques on LB/IPTG/Xgal plate | |
Assay of selected phage display peptides for target binding by ELISA: One well of a microtiter plate was coated with patient serum and another well with BSA as a negative control. This procedure was carried out in triplicate. The colour change in wells coated with patient serum showed that the phage had successfully bound to patient serum. The mean OD415 reading of the ELISA assay in wells coated with patient serum was 0.111, which was approximately 3 fold higher than the negative control BSA (OD415 = 0.035).
Sequencing: Similar DNA sequences were obtained from 5 randomly selected phagemids that expressed reactive peptide. The nucleotide sequence and deduced amino acid sequence of the peptide that potentially can bind to the patient serum are shown in bold in Fig. 1.
Database search and analysis: After searching from non-redundant GenBank CDS database using NCBI Blast, the amino acid sequence revealed homology to part of the sequence, 35FDAWLAAQEF44, of B. pseudomallei site-specific recombinase, a phage integrase family protein, with the score of 22.3, E-value of 23 and sequence coverage of 58.3%.
DISCUSSION
Phage display technology was used for the study of B. pseudomallei by Chan and Nathan (2005) to decipher an antigenic peptide presenting a consensus peptide sequence, TKSMALSG, which closely resembles part of the active site sequence, 435GTSMATPHVAG445, of B. pseudomallei serine metalloprotease. Using the same approach, Nathan et al. (2005) subsequently identified chimeric mouse-human antibody fragments (Fabs) that had neutralizing properties against B. pseudomallei protease. Using two different panning procedures, Zou et al. (2007) obtained seven different scFv phage antibodies that interacted with the heat-killed whole bacterial cells of B. pseudomallei and B. mallei. All these studies prove the usefulness of phage display technology in seeking novel peptides which are of diagnostic and therapeutic value.
In the present study, serum obtained from a patient with melioidosis was used as the template and phage display technology applied to identify the immunogenic mimotope. The mimotope sequence acquired after translating from the nucleotide sequence carried by a phagemid was NKNSFDAWLQSF, partly homologous to B. pseudomallei site-specific recombinase which is a phage integrase family protein. Site-specific recombinases are present in all bacteria and contribute to the evolution of their virulence and antibiotic resistance properties (Hartl and Jones, 2002). Each type of site-specific recombinase binds with a specific sequence in duplex DNA. When the site is present in each of two duplex DNA molecules, the recombinase brings the sites together and catalyzes a reciprocal exchange between the duplexes (Hartl and Jones, 2002). An example of a site-specific recombinase is an enzyme called the Cre recombinase, encoded by a gene in the Escherichia coli bacteriophage P1, which initiates recombination by cleaving and exchanging preferentially on the bottom strand of its loxP target sequence (Van Duyne, 2001).
Other novel antigens of B. pseudomallei obtained in other studies using different approaches are a LolC homologue (Harland et al., 2007), a 18.7 kDa recombinant protein expressed by a B. pseudomallei-specific gene, pBps-1 (Wongprompitak et al., 2001), a 57.1 kDa protein homologous to the chaperonins encoded by groEL gene (Woo et al., 2001a) and a 44.4 kDa protein homologous to the maltose-binding protein encoded by malE (Woo et al., 2001b). Mouse model and cloning methods were being applied in those studies. Other approaches such as proteomic-based approaches have also been extensively used for this purpose. For instance, Harding et al. (2007) reported the identification of nine immunogenic surface proteins of B. pseudomallei following screening B. pseudomallei proteomes with human sera. The utilization of Caenorhabditis elegans host system has also been reported in the identification of novel virulence factors of B. pseudomallei (Gan et al., 2002).
Since a reliable commercial diagnostic kit for melioidosis is still in demand, the discovery of all these immunogenic proteins, as well as the mimotope obtained in our study, with downstream testing for their specificity and sensitivity for melioidosis, brings a ray of hope for the development of a more specific and rapid diagnostic method or vaccine. It is important that the disease is diagnosed in the early acute stage and also in the chronic and subclinical stages, so that appropriate intervention can be carried out.
CONCLUSION
This study has shown the utility of peptide phage display as a method for isolating B. pseudomallei immunogenic mimotopes that interact with serum from patients infected with melioidosis. Further studies should be carried out to evaluate this peptide for its sensitivity and specificity as a diagnostic reagent.
REFERENCES
- Akita, N., F. Maruta, L.W. Seymour, D.J. Kerr, A.L. Parker, T. Asai, N. Oku, J. Nakayama and S. Miyagawa, 2006. Identification of oligopeptides binding to peritoneal tumors of gastric cancer. Cancer Sci., 97: 1075-1081.
PubMedDirect Link - Brown, N.F., J.A. Boddey, C.P. Flegg and I.R. Beacham, 2002. Adherence of Burkholderia pseudomallei cells to cultured human epithelial cell lines is regulated by growth temperature. Infect. Immun., 70: 974-980.
Direct Link - Chan, S.W. and S. Nathan, 2005. Epitope mapping of Burkholderia pseudomallei serine metalloprotease: Identification of serine protease epitope mimics. FEMS Immunol. Med. Microbiol., 43: 37-44.
CrossRefDirect Link - Chen, G., X. Wang, J. Yu, S. Varambally and J. Yu et al., 2007. Autoantibody profiles reveal ubiquilin 1 as a humoral immune response target in lung adenocarcinoma. Cancer Res., 67: 3461-3467.
CrossRefPubMedDirect Link - Gan, Y.H., K.L. Chua, H.H. Chua, B. Liu, C.S. Hii, H.L. Chong and P. Tan, 2002. Characterization of Burkholderia pseudomallei infection and identification of novel virulence factors using a Caenorhabditis elegans host system. Mol. Microbiol., 44: 1185-1197.
CrossRefDirect Link - Harding, S.V., M. Sarkar-Tyson, S.J. Smither, T.P. Atkins and P.C. Oyston et al., 2007. The identification of surface proteins of Burkholderia pseudomallei. Vaccine, 25: 2664-2672.
CrossRefDirect Link - Harland, D.N., K. Chu, A. Haque, M. Nelson and N.J. Walker et al., 2007. Identification of a LolC homologue in Burkholderia pseudomallei, a novel protective antigen for melioidosis. Infect. Immun., 75: 4173-4180.
CrossRef - Jiao, Y., P. Zhao, J. Zhu, T. Grabinski and Z. Feng et al., 2005. Construction of human naive Fab library and characterization of anti-met Fab fragment generated from the library. Mol. Biotechnol., 31: 41-54.
CrossRefDirect Link - Nathan, S., C. Rader and C.F. Barbas, 2005. Neutralization of Burkholderia pseudomallei protease by Fabs generated through phage display. Biosci. Biotechnol. Biochem., 69: 2302-2311.
Direct Link - Puthucheary, S.D., N. Parasakthi and M.K. Lee, 1992. Septicaemic melioidosis: A review of 50 cases from Malaysia. Tran. Roy. Soc. Trop. Med. Hyg., 86: 683-685.
CrossRef - Reckseidler, S.L., D. DeShazer, P.A. Sokol and D.E. Woods, 2001. Detection of bacterial virulence genes by subtractive hybridization: identification of capsular polysaccharide of Burkholderia pseudomallei as a major virulence determinant. Infect. Immun., 69: 34-44.
CrossRefDirect Link - Sambrook, J., E.F. Fritsch and T.A. Maniatis, 1989. Molecular Cloning: A Laboratory Manual. 2nd Edn., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York, USA., ISBN-13: 9780879695774, Pages: 397.
Direct Link - Sergeeva, A., M.G. Kolonin, J.J. Molldrem, R. Pasqualini and W. Arap, 2006. Display technologies: application for the discovery of drug and gene delivery agents. Adv. Drug Delivery Rev., 58: 1622-1654.
CrossRefPubMedDirect Link - Shen, Y., X. Yang, N. Dong, X. Xie, X. Bai and Y. Shi, 2007. Generation and selection of immunized Fab phage display library against human B cell lymphoma. Cell Res., 17: 650-660.
Direct Link - Smith, G.P., V.A. Petrenko and L.J. Matthews, 1998. Cross-linked filamentous phage as an affinity matrix. J. Immunol. Methods, 215: 151-161.
CrossRef - Tungtrakanpoung, R., P. Pitaksajjakul, N. Na-Ngarm, W. Chaicumpa and P. Ekpo et al., 2006. Mimotope of Leptospira from phage-displayed random peptide library is reactive with both monoclonal antibodies and patients sera. Vet. Microbiol., 115: 54-63.
CrossRef - Van-Duyne, G.D., 2001. A structural view of Cre-loxp site-specific recombination. Annu. Rev. Biophys. Biomol. Struct., 30: 87-104.
Direct Link - Wongprompitak, P., C. Thepthai, S. Songsivilai and T. Dharakul, 2001. Burkholderia pseudomallei-specific recombinant protein and its potential in the diagnosis of melioidosis. Asian Pac. J. Allergy Immunol., 19: 37-41.
Direct Link - Woo, P.C.Y., P.K.L. Leung, H.W. Tsoi and K.Y. Yuen, 2001. Cloning and characterisation of malE in Burkholderia pseudomallei. J. Med. Microbiol., 50: 330-338.
CrossRefDirect Link - Woo, P.C., P.K. Leung, S.S. Wong, P. L. Ho and K.Y. Yuen, 2001. groEL encodes a highly antigenic protein in Burkholderia pseudomallei. Clin. Diagn. Lab. Immunol., 8: 832-836.
CrossRef - Zou, N., T. Newsome, B. Li, S. Tsai and S.C. Lo, 2007. Human single-chain Fv antibodies against Burkholderia mallei and Burkholderia pseudomallei. Exp. Biol. Med., 232: 550-556.
Direct Link