
T. Dzenda
Department of Veterinary Physiology and Pharmacology, Ahmadu Bello University, Zaria, Nigeria
J.O. Ayo
Department of Veterinary Physiology and Pharmacology, Ahmadu Bello University, Zaria, Nigeria
C.A.M. Lakpini
National Animal Production Research Institute, Ahmadu Bello University, Zaria, Nigeria
A.B. Adelaiye
Department of Human Physiology, Ahmadu Bello University, Zaria, Nigeria
International Journal of Zoological Research
Year: 2011 | Volume: 7 | Issue: 1 | Page No.: 49-58







ABSTRACT
Adult African giant rats (Cricetomys gambianus, Waterhouse) were live-trapped in Zaria, Nigeria over a period of three years, cutting across all the seasons of each year. They were sexed and weighed immediately after capture, with the aim of determining changes in their live weights with seasons and sex. The overall live weight (Mean±SEM) of the African giant rats was 1.21±0.01 kg (n = 363). The live weight of African giant rats captured during the rainy season (1.17±0.02 kg) was significantly lower than that during the harmattan (1.25±0.02 kg; p<0.01) and hot-dry (1.23±0.02 kg, p<0.05) seasons. The male live weight of 1.28±0.01 kg was significantly higher (p<0.0001) than that of the female (1.14±0.01 kg). Male-biased sexual size dimorphism was observed during all the seasons, but was highest during the hot-dry season. The seasonal variation in live weight may be because food was more available in the wild during the harvest (dry) seasons (harmattan and hot-dry seasons) than the planting (rainy) season. The relatively low ambient temperature and relative humidity of the harmattan season was more favourable to growth, which may account for the higher live weight recorded during the season. The sex variation in live weight may be as a result of differential growth rate/time between the sexes. In conclusion, the African giant rats are lightest during the rainy season and the live weight of the male is higher than that of the female regardless of the season.
PDF Abstract XML References Citation
Received: April 21, 2010;
Accepted: June 08, 2010;
Published: August 21, 2010
How to cite this article
T. Dzenda, J.O. Ayo, C.A.M. Lakpini and A.B. Adelaiye, 2011. Seasonal and Sex Variations in Live Weights of Captive African Giant Rats Cricetomys gambianus, Waterhouse) in the Northern Guinea Savannah Zone of Nigeria. International Journal of Zoological Research, 7: 49-58.
DOI: 10.3923/ijzr.2011.49.58
URL: https://scialert.net/abstract/?doi=ijzr.2011.49.58
DOI: 10.3923/ijzr.2011.49.58
URL: https://scialert.net/abstract/?doi=ijzr.2011.49.58
INTRODUCTION
Seasonal variations in the Live Weight (LW) of man (Simondon et al., 2007), other mammals (Purusothaman et al., 2008), birds (Sargisson et al., 2007) and fishes (Samsun et al., 2007) have been described both in the temperate and tropical regions of the world. In temperate regions generally, LW has been found to be higher in winter and spring than in summer, while in the tropics it is higher in the dry season than the rainy season. Different reasons have been suggested for the seasonal variation in LW. For example, most species have been observed to be heavier at times of cold stress as a result of increased thyroid activity in response to the low ambient temperature associated with the season (Westerterp, 2001). Hypovitaminosis D, which is most prevalent in winter, increased foetal birth and neonatal weights in rats (Eyles et al., 2003) and guinea pigs (Rummens et al., 2002). Vitamin D suppresses cell proliferation and promotes cell elimination via apoptosis in a variety of tissues (DeLuca et al., 1990). Thus, its absence in the prenatal period may lead to inappropriately high cell numbers, which could subsequently influence the size of the offspring (McGrath et al., 2005). In addition, melatonin may have a relationship with seasonality, vitamin D, growth and bone mass (Cardinali et al., 2003). The nutritional status of animals in the wild varies with season and may also contribute to the seasonal differences in LW (Colares et al., 2000).
Sexual dimorphism, widespread in the animal kingdom, describes differences between the sexes in size, shape and many other traits (Kupfer, 2009). Males and females of nearly all animals differ in their body size, a phenomenon called Sexual Size Dimorphism (SSD). The SSD and seasonal variation in SSD are common among rodents. Kraskov et al. (2005) observed significant SSD in 7 desert rodent species in both Summer and Winter. In guinea pigs, the larger size of males has been attributed to significant differences in the activational effects of testosterone on body size (Czaja, 1984). The subterranean rodent, Ctenomys talarum (tuco-tuco) in Argentina differs in demographic traits and shows significant SSD in both LW and length, with males being larger than females (Zenuto, 1999). Although, the cause of sex variation in AGRs has not been investigated, various factors such as genetic selection (Stillwell et al., 2010), phenotypic plasticity in growth rate (Alonso et al., 2009), sexual selection (Kupfer, 2009), differences in environmental sensitivity (Irie and Morimoto, 2008), feeding habits (Goldschmidt et al., 2009), levels of physical activity (Ghosh, 2004) and sex hormones (John-Alder et al., 2007) have been suggested as probable causes for the SSD observed in other species. The SSD has been traditionally described using body size measures, including LW, single linear measures capturing total length, or the length of some specific body part (Hood, 2000). Body size is one of the most important quantitative traits of an animal because of its pervasive effects on physiological, ecological and life-history processes (John-Alder et al., 2007). Relative body size (sex with larger body size/sex with smaller body size) has been used as a measure of SSD (Tamate and Maekawa, 2006). There is a dearth of information on the effect of season on LW of the African Giant rat (Cricetomys gambianus, Waterhouse).
The aim of the present study was to measure the LWs of captive AGR in order to determine their seasonal and sex variations in the Northern Guinea Savannah zone of Nigeria. The occurrence of SSD was also examined and the available information on feed consumption, growth rate and age at maturity of both sexes of the AGR was used in conjunction with meteorological data to evaluate the influence of sexual selection on such variability.
MATERIALS AND METHODS
Study species: The African Giant rat (Cricetomys gambianus Waterhouse, 1840) (AGR) is found throughout tropical and subtropical Africa, south of the Sahara desert down to about 27° South latitude (Novak, 2009). Their native range stretches from the Atlantic Ocean coast of West Africa, east across the Congo Basin to the Indian Ocean coast of East Africa (Peterson et al., 2006) and southwards into the Transvaal and KwaZulu-Natal provinces of South Africa (Malekani et al., 2002). Aside from bushes, AGRs are also found near human habitation, where they live in holes under garages or under piles of stones or bricks. Mean LW is said to vary greatly among groups of AGRs, but individuals normally weigh between 1 and 2 kg, with males being heavier than females (Novak, 2009). Available data on SSD in LW of the AGR vary widely and are largely based on a few numbers of animals. For example, Halcrow (1958) used 6 AGRs (1 male and 5 females), Ajayi (1977) used 17 AGRs (10 males and 7 females) and Perry et al. (2006) used only 5 AGRs (2 males and 3 females) to infer sex differences in weight. The present study was performed to undertake a broad based study on SSD in LW of AGRs in their natural habitat, spanning over a period of 3 years (2006-2009) and utilizing larger numbers of AGRs of either sex. In addition, attempt has been made to assess the influence of season on the general LW and SSD in the AGR.
Study area: The study was conducted in Samaru (11°10/N, 07°38/E), Zaria, located in the Northern Guinea Savannah zone of Nigeria, at an altitude of 686 m above maximum sea level. The zone is characterised by three major seasons: harmattan (November-February), hot-dry (March-April) and rainy (May-October) seasons (Igono et al., 1982; Ayo et al., 1999).
Animal trapping: Metal live box traps of various sizes and manufacturers were baited with peanuts, maize and baked cake made from ground beans. Trapping occurred both in bushes and residential areas. Since the AGRs are nocturnal animals, traps were set before dusk and checked at dawn. The traps were retrieved during the day to prevent capture of diurnal, non-target taxa. The trap, capture and handling methods utilized in the study were in accordance with the American Society of Mammalogists’ guidelines (Gannon et al., 2007) and were approved by the Ahmadu Bello University Animal Care and Use Committee. A total of 363, apparently, healthy adult AGRs were live-trapped from bushes and residential areas in and around Zaria over a 3-year period (2006-2009) cutting across all three seasons of each year.
Live weight measurements: Measurements of LWs were taken from each of the 363 captured AGRs as soon as they were brought from the wild using a weighing balance (50 kgx200 g, Model 250, Salter, England). The difference between the weight of an empty metal cage and that of the same cage containing each AGR was recorded as the LW of the AGR. Captive AGRs with LWs of at least 0.8 kg were considered as adult (Ajayi, 1977). LWs of pregnant females were not included in the data obtained. After the measurements, all the AGRs were housed in metal cages specially constructed for further experiments.
Statistics: The data obtained were expressed as mean± standard error of the mean (Mean±SEM). Seasonal variations were analyzed using one-way Analysis of Variance (ANOVA) with Tukey’s post-hoc test, while two-tailed paired Student’s t-test was used to determine sex differences in the values, using GraphPad Prism, version 4.03 for Windows (GraphPad Software, San Diego, California, USA, http://www.graphpad.com). Correlation analysis between LWs of the AGR and meteorological parameters was done using one-tailed Pearson’s correlation. In all cases, values of p<0.05 were considered significant. SSD was calculated as a function of mean male weight divided by mean female weight (Lucherini et al., 2006).
RESULTS AND DISCUSSION
Daily meteorological data for the locality, covering the experimental period were obtained from the Meteorological Data Processing Unit, Department of Soil Science, Faculty of Agriculture, Ahmadu Bello University, Zaria, Nigeria, located about 1 km from the experimental site. The analyzed meteorological data for the study period are presented in Table 1. The highest ambient (dry-bulb) temperature of 29.31±0.47°C was recorded during the hot-dry season, while the lowest value of 23.39±0.36°C was obtained during the harmattan season. The highest relative humidity value of 66.78±0.48% was obtained during the rainy season, while the lowest value of 16.44±0.28% was recorded during the harmattan season.
| Table 1: | Seasonal changes in meteorological conditions prevailing during the study period |
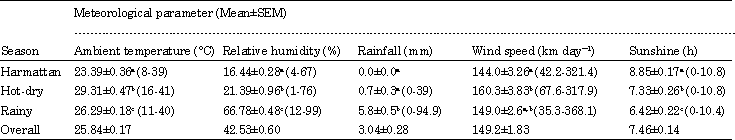 | |
| Mean values of each parameter with different superscript letters and/or not sharing a letter are significantly different (p<0.05). Range values (minimum-maximum) are indicated in brackets with means | |
| Table 2: | Seasonal and sex variations in live weights of captive African giant rats (Cricetomys gambianus, Waterhouse) |
 | |
The mean amount of rainfall during the rainy season was 5.83±0.52 mm, while only 0.74±0.31 mm was recorded in the hot-dry season. No rainfall was recorded during the harmattan season. The lowest wind velocity of 144.0±3.26 km day-1 was obtained during the harmattan season, while the highest value of 160.3±3.83 km day-1 was measured during the hot-dry season. The lowest sunshine duration of 6.42±0.22 h was recorded in the rainy season; while the highest value of 8.85±0.17 h was obtained in the harmattan season, when usually there was no cloud cover. The predominant wind directions during the months without rainfall (November-March) were South-East and North-East, while South-West and North-West winds dominated during the rainy season. The transition months of April and October were predominated by south-west and south-east winds.
The LWs of the AGRs recorded throughout the study period are presented in Table 2. The highest recorded individual LW of the AGRs during the study period was 1.7 kg. The overall mean LW was 1.21±0.01 kg (n = 363). The mean LW of the AGRs captured during the rainy season was 1.17±0.02 kg and the value was significantly lower than those obtained during the harmattan (1.25 ± 0.02 kg; p<0.01) and hot-dry (1.23±0.02 kg; p<0.05) seasons. The mean LW of male AGRs was 1.28±0.01 kg and the value was significantly higher (p<0.0001) than that of 1.14±0.02 kg recorded in the females. The mean LW of male AGRs was higher than that of the females regardless of season, but the difference was highest during the hot-dry season. The overall SSD was 1.12 in favour of male AGRs. The highest SSD value of 1.15 (p<0.0001) was recorded during the hot-dry season; while the lowest value of 1.09 (p<0.01) was recorded during the harmattan season. The LW was negatively correlated with ambient temperature (r = -0.2167; p>0.05), relative humidity (r = -0.9861; p< 0.01), rainfall (r = -0.9891; p<0.01) and wind speed (r = -0.0092; p>0.05), but was positively correlated with sunshine (r = 0.9097; p<0.05) (Table 3).
The overall mean LW value of the AGR obtained in the present study (1.21±0.01 kg) fell within the range of mean adult LW reported by other investigators (Ajayi, 1975, 1977; Perry et al., 2006). The value was higher than that of 0.86±0.03 kg reported by Onyeanusi et al. (2009) in the same locality. Perry et al. (2006) used adult AGRs of 1.0-1.4 kg and considered a LW Mean values between sexes with different superscript letters are significantly different (a,bp<0.01; c,dp<0.0001; e,fp<0.001); Mean values between seasons with different or not sharing superscript numbers (1,2) are significantly different (p<0.05).
| Table 3: | Correlation coefficients between live weight of the African giant rat and meteorological parameters |
 | |
| nsNon significant correlation; * Significant correlation (p<0.05); ** Very significant correlation (p<0.01) | |
Range values (minimum-maximum) are indicated in brackets with means of 0.5 kg to be juvenile. Adult LWs of 0.8-1.7 kg were used in the present study. According to a review on the bioprofile of the AGR (Novak, 2009), mean LW varies greatly among groups of AGRs, but individuals normally weigh between 1 and 2 kg. However, adult LWs as low as 0.7 kg (Mott, 2004) and as high as 2.8 kg (Cooper, 2008) have been reported for the AGR. Thus, geographic variation in adult LW may occur in the AGR as is the case with other species (Lucherini et al., 2006). In addition, a distinction has to be made between captive and domesticated AGRs with regards to the availability of quality feed, since this may affect LW at maturity (Ajayi and Tewe, 1978).
The results obtained in the present study also showed that season had a significant effect on LWs of the AGRs captured from the wild. Thus, the highest LWs in both sexes were obtained during the harmattan season. Two reasons may be responsible for this finding. First, the lowest temperature and humidity were recorded during the harmattan season, indicating that the AGRs experienced more cold stress during the season compared with the other seasons. Ambient temperature values during the day as low as 8°C were recorded during the harmattan season and, since the lower critical temperature for AGRs has been reported to be 21°C (Knight, 1988), indicating that the AGR would have been under cold stress during the season. Furthermore, the low relative humidity during the harmattan season favours increased evaporation of water from the body which is usually accompanied by evaporative heat loss, occurring when heat conservation is needed to combat the effects of cold stress. Thus, the relative humidity was negatively correlated with LW (p<0.05) and values as low as 4% were recorded during the harmattan season and this exacerbated the cold stress. The finding of the present study is in agreement with the result obtained by Westerterp (2001) that cold stress stimulates thyroid activity through the hypothalamus, leading to increased metabolic rate, feed consumption and, consequently, increased LW. Secondly, the harmattan season coincided with the harvest period of the year when food was most abundant and available in the wild for the AGRs to eat and thus increase their LW. Moreover, AGRs are known to store a lot of food in their burrows during the harvest season (Novak, 2009). The observation on food availability for the AGR during the harmattan season agrees with that of Susic et al. (2005), who reported that food availability plays a key role in the seasonality of LWs in lambs and the finding of Colares et al. (2000) that the nutritional status of animals in the wild varies with season and may also contribute to the seasonal differences in LW.
The combined effects of meteorological and nutritional factors may be responsible for the increased LW recorded during the harmattan season in the present study and this fact is in agreement with the report by Stillwell et al. (2010), who showed that diet quality and developmental temperature are arguably the most important ecological and environmental variables that influence body size. The long duration of the harmattan season, lasting from November to February allowed the adequate interaction of the different factors that favour increased LW. The harmattan season prevailing during the present study period was actually a dry season with no rainfall and the lowest relative humidity during the year (Table 1).
The hot-dry season was actually hot with recorded ambient temperatures some times reaching 41°C during the day, which is well above the upper critical temperature of 34°C for the AGR (Knight, 1988). The AGRs were, therefore, subjected to heat stress during the season. The effects of heat stress on LWs of animals are opposite to those of cold stress, decreasing metabolic rate and feed consumption, leading to decreased body weight (Westerterp, 2001; AL-Mansour, 2004). The LW of the AGRs during the hot-dry season was less than that of the harmattan season, although the difference was not statistically significant. There are many possible explanations for this non-significant difference. First, the hot-dry season follows immediately the harmattan season, covering a very short period between March and April. Thus, the duration of the heat stress was probably too short to impose a significant decrease in LW of the AGRs. Conditions of the hot season leading to a drop in LW have been shown to be caused by a drop in feed consumption during the season (Concannon et al., 2001), which agrees with our preliminary observations on the AGR (unpublished data) that a significant drop (p<0.001) in feed consumption occurred during the hot-dry season. According to Concannon et al. (2001), spontaneous declines in feed consumption of woodchucks (another large rodent species) during the hot Summer began 2-6 weeks before resulting in declines in LW. The hot-dry season in the present study lasted only 8 weeks and 5 days (March-April). It is, therefore, conceivable that the effect of the hot-dry season on LW of the AGR in the present study may have become significant only towards the end of the short season or even after the season was over. Secondly, the low relative humidity of the hot-dry season during the present study favoured evaporative heat loss and became succour for the heat stress, because heat stress results from the combined effects of high relative humidity and ambient temperature (Broucek et al., 2008). Thirdly, the AGRs are nocturnal and spend their day time in burrows that are cool during a hot day (Novak, 2009), therefore, avoiding the thermally stressful conditions of the hot-dry season. This possibly explains the non-significant correlation between LW and ambient temperature in the present study.
The results also showed that the lowest LWs for the AGR were recorded during the rainy season. Infact, LW was negatively correlated with rainfall (p<0.01). The ambient temperature conditions of the rainy season were moderate and largely within the thermoneutral zone of 21-34°C for the AGR (Knight, 1988), therefore, the drastic drop in LW may not be attributed to heat stress. Limited feed availability may be the apparent cause of the low LW, since the rainy season coincided with the clearing and planting period when food was most scarce in the wild. The finding agrees with that of Susic et al. (2005) who attributed seasonal variation in LW to feed availability. Studies in humans in tropical Africa also showed reduced LW during the rainy season; which was attributed to strong physical activity during field work, leading to negative energy balance from the onset of agricultural labour (Simondon et al., 2007). Similarly, the duration of gestation in women reduced during the rainy season (Rayco-Solon et al., 2005), resulting in lower birth weights.
The result of the present study on the AGR is in agreement with that on infants (Prentice et al., 1983), adult women (Simondon et al., 2007) and avian species (Clark, 1979) in the tropics, in which LW was found to be higher in the dry season than the rainy season in all cases. The present study for the first time reports seasonal variations in LWs of AGRs in their natural habitat.
The fact that the male adult LW was significantly higher than that of the female agrees with the findings of Ajayi (1977) in the rain forest zone of Nigeria and those of Perry et al. (2006) and Cooper (2008) in other ecological zones, who have also reported higher LWs in male AGRs, although seasonal variations in SSD was not considered in the studies. Besides, the researchers carried out their experiments using few numbers (n = 5-17), whereas the present study utilized large numbers of AGR (n = 363). No SSD values have been previously given for the AGR. Cox et al. (2006) reported that SSD studies are environmentally sensitive even within the same season.
Hence, it is conceivable that SSD values may differ between domesticated and wild AGRs.
Although, the cause of sex variation in AGRs was not investigated in the present study, various factors have been suggested as probable causes for the SSD observed in other rodent species. Differential growth rates in rats from puberty onward contribute to the larger size of adult males. A rise of circulating androgens in males at a critical period has been shown to play a role in this increased growth (Slob and Bosch, 1975). Gender differences in the circulating GH profiles of rats are responsible, to varying degrees, for observed sexual dimorphisms including body growth (Agrawal and Shapiro, 2001). According to Ewer (1967), the growth rate of AGRs peaks at about 25 days for body length and 35 days for tail length and is similar between male and female young, suggesting that males attain their larger size due to a prolonged growing period.
Results of some of our preliminary work on the AGR indicate that feed consumption is significantly higher in males than females, which may favour male-biased SSD. In the present study, ambient temperature and relative humidity values were significantly lower during the harmattan season, favouring growth, which apparently accounted for the significantly higher LWs in both sexes and, consequently, lower SSD during the season. The male-biased SSD was highest during the hot-dry season, implying that male AGRs responded more favourably than female AGRs to the thermally stressful conditions of the season. Individual ambient temperatures of up to 41°C were recorded during the hot-dry season. AGRs are sensitive to heat. Ajayi (1975) observed two captive AGRs that died from heat prostration in temperatures above 32°C. The sexes commonly respond differently to environmental variation and interactions between environmental variables have different effects on the sexes, affecting patterns of SSD (Stillwell et al., 2010).
The present study demonstrates, for the first time, seasonal and sex variations in LWs of AGRs in their natural habitat. The LWs of AGRs captured from the wild in the Northern Guinea Savannah zone of Nigeria vary with season and SSD is male-biased across all seasons. Geographic variation in LW of the AGR within its ecological range needs to be investigated, since different ranges of adult LWs are reported.
ACKNOWLEDGMENTS
Authors acknowledge the assistance of Mr. Gbaden Achinga and his associates in the trap and restraint of the AGRs. We are grateful to the Department of Soil Science, Faculty of Agriculture, Ahmadu Bello University, Zaria, Nigeria, for making available to us the meteorological data used in the study.
REFERENCES
- Agrawal, A.K. and B.H. Shapiro, 2001. Intrinsic signals in the sexually dimorphic circulating growth hormone profiles of the rat. Mol. Cell. Endocrinol., 173: 167-181.
CrossRef - Ajayi, S.S., 1975. Observations on the biology, domestication and reproductive performance of the African giant rat (Cricetomys gambianus, Waterhouse) in Nigeria. Mammalia, 39: 343-364.
Direct Link - Ajayi, S.S., 1977. Live and carcass weights of giant rat (Cricetomys gambianus, Waterhouse) and domestic rabbit (Oryctolagus cuniculus L). Afr. J. Ecol., 15: 223-227.
Direct Link - Ajayi, S.S. and O.O. Tewe, 1978. Performance of the African giant rat (Cricetomys gambianus Waterhouse) on commercial rations and varying dietary protein levels. Lab. Anim., 12: 109-112.
CrossRef - Al-Mansour, M.I., 2004. Seasonal variation in basal metabolic rate and body composition within individual sanderling bird Calidris alba. J. Biol. Sci., 4: 564-567.
CrossRefDirect Link - Alonso, J.C., M. Magana, J.A. Alonso, C. Palacin, C.A. Martin and B. Martin, 2009. The most extreme sexual size dimorphism among birds: Allometry, selection and early juvenile development in the great bustard (Otis tarda). The Auk, 126: 657-665.
Direct Link - Ayo, J.O., S.B. Oladele, S. Ngam, A. Fayomi and S.B. Afolayan, 1999. Diurnal fluctuations in rectal temperature of the Red Sokoto goat during the harmattan season. Res. Vet. Sci., 66: 7-9.
CrossRefDirect Link - Broucek, J., P. Kisac, M. Uhrincat, A. Hanus and F. Benc, 2008. Effect of high temperature on growth performance of calves maintained in outdoor hutches. J. Anim. Feed Sci., 17: 139-146.
Direct Link - Cardinali, D.P., M.G. Ladizesky, V. Boggio, R.A. Cutrera and C. Mautalen, 2003. Melatonin effects on bone: Experimental facts and clinical perspectives. J. Pineal Res., 34: 81-87.
PubMedDirect Link - Colares, E.P., I.G. Colares, A. Bianchini and E.A. Santos, 2000. Seasonal variations in blood parameters of the Amazonian manatee, Trichechus inunguis. Braz. Arch. Biol. Technol., 43: 165-171.
Direct Link - Concannon, P., K. Levac, R. Rawson, B. Tennant and A. Bensadoun, 2001. Seasonal changes in serum leptin, food intake, and body weight in photoentrained woodchucks. Am. J. Physiol. Regulatory Integrative Comp. Physiol., 281: R951-R959.
Direct Link - Cooper, R.G., 2008. Care, husbandry and diseases of the African Giant rat (Cricetomys gambianus). J. South Afr. Vet. Ass., 79: 62-66.
PubMed - Cox, R.M., V. Zilberman and H.B. John-Alder, 2006. Environmental sensitivity of sexual size dimorphism: Laboratory common garden removes effects of sex and castration on lizard growth. Functional Ecol., 20: 880-888.
Direct Link - DeLuca, H.F., J. Krisinger and H. Darwish, 1990. The vitamin D system: 1990. Kidney Int. Suppl., 29: S2-S8.
PubMedDirect Link - Eyles, D., J. Brown, A. Mackay-Sim, J. McGrath and F. Feron, 2003. Vitamin D3 and brain development. Neuroscience, 118: 641-653.
PubMedDirect Link - Gannon, W.L., R.S. Sikes and The Animal Care and Use Committee of the American Society of Mammalogists, 2007. Guidelines of the American society of mammalogists for the use of wild mammals in research. J. Mammal., 88: 809-823.
CrossRefDirect Link - Ghosh, A., 2004. Age and sex variation in measures of body composition among the elderly bengalee hindus of Calcutta, India. Coll. Antropol., 28: 553-561.
Direct Link - Goldschmidt, B., A. Mota-Marinho, C. Araujo-Lopes, M.A. Bruck-Goncalves and D. Matos-Fasano et al., 2009. Sexual dimorphism in the squirrel monkey, Saimiri ustus (I. Geoffroy, 1844) (Primates, Cebidae). Braz. J. Biol., 69: 171-174.
Direct Link - Hood, C.S., 2000. Geometric morphometric approaches to the study of sexual size dimorphism in mammals. Hystrix, 11: 77-90.
Direct Link - Igono, M.O., E.C.I. Molokwu and Y.O. Aliu, 1982. Body temperature responses of Savanna Brown goat to the harmattan and hot-dry season. Int. J. Biometeorol., 26: 225-230.
CrossRefDirect Link - John-Alder, H.B., R.M. Cox and E.N. Taylor, 2007. Proximate developmental mediators of sexual dimorphism in size: Case studies from squamate reptiles. Integr. Comp. Biol., 47: 258-271.
Direct Link - Irie, T. and N. Morimoto, 2008. Phenotypic plasticity and sexual dimorphism in size at post-juvenile metamorphosis: Common-garden rearing of an intertidal gastropod with determinate growth. Biol. Bull., 215: 126-134.
Direct Link - Westerterp, K.R, 2001. Seasonal variation in body weight: An experimental case study. J. Therm. Biol., 26: 525-527.
Direct Link - Knight, M.H., 1988. Thermoregulation in the largest African Cricetid, the giant rat (Cricetomys gambianus). Comp. Biochem. Physiol. A: Comp. Physiol., 89: 705-708.
PubMedDirect Link - Kraskov, B.R., S. Morand, H. Hawlena, I.S. Khokhlova and G.I. Shenbrot, 2005. Sex-biased parasitism, seasonality and sexual size dimorphism in desert rodents. Oecologia, 146: 209-217.
Direct Link - Kupfer, A., 2009. Sexual size dimorphism in caecilian amphibians: Analysis, review and directions for future research. Zoology, 112: 362-369.
PubMedDirect Link - Lucherini, M., C. Manfredi, E. Luengos, F.D. Mazim, L. Soler and E.B. Casanave, 2006. Body mass variation in the geoffroys cat (Oncifelis geoffroyi). Rev. Chil. Hist. Nat., 79: 169-174.
Direct Link - Malekani, M., L.M. Westlin, J.J. Paulus and H.C. Potgieter, 2002. Oestrous occurrence in captive female Cricetomys gambianus (Rodentia: Cricetidae). J. Zool., 257: 295-301.
Direct Link - McGrath, J.J., D. Keeping, S. Saha, D.C. Chant, D.E. Lieberman and M.J. O'Callaghan, 2005. Seasonal fluctuations in birth weight and neonatal limb length: Does prenatal vitamin D influence neonatal size and shape?. Early Hum. Dev., 81: 609-618.
PubMedDirect Link - Perry, N.D., B. Hanson, W. Hobgood, R.L. Lopez and C.R. Okraska et al., 2006. New invasive species in southern Florida: Gambian rat (Cricetomys gambianus). J. Mammal., 87: 262-264.
Direct Link - Onyeanusi, B.I., A.A. Adeniyi, J.O. Ayo, C.S. Ibe and C.G. Onyeanusi, 2009. A comparative study of the urinary system of the African giant rat (Cricetomys gambianus Waterhouse) and the Wistar rat. Pak. J. Nutr., 8: 1043-1047.
CrossRefDirect Link - Peterson, A.T., M. Papaş, M.G. Reynolds, N.D. Perry and B. Hanson et al., 2006. Native-range ecology and invasive potential of Cricetomys in North America. J. Mammal., 87: 427-432.
Direct Link - Prentice, A.M., R.G. Whitehead, M. Watkinson, W.H. Lamb and T.J. Cole, 1983. Prenatal dietary supplementation of African women and birth-weight. Lancet, 321: 489-492.
CrossRefPubMedDirect Link - Purusothaman, M.R., A.K. Thiruvenkadan and K. Karunanithi, 2008. Seasonal variation in body weight and mortality rate in Mecheri adult sheep. Livest. Res. Rural Dev., 20.
Direct Link - Rayco-Solon, P., A.J. Fulford and A.M. Prentice, 2005. Maternal preconceptional weight and gestational length. Am. J. Obstet. Gynaecol., 192: 1133-1136.
Direct Link - Rummens, K., R. Van Bree, E. Van Herck, Z. Zaman, R. Bouillon, F.A. Van Assche and J. Verhaeghe, 2002. Vitamin D deficiency in guinea pigs: Exacerbation of bone phenotype during pregnancy and disturbed fetal mineralization, with recovery by 1,25(OH)2D3 infusion or dietary calcium-phosphate supplementation. Calcif. Tissue Int., 71: 364-375.
PubMedDirect Link - Samsun, N., F. Kalayci and O. Samsun, 2007. Seasonal variation in length, weight, and sex distribution of turbot (Scophthalmus maeoticus Pallas, 1811) in the Sinop region (Black sea) of Turkey. Turk. J. Zool., 31: 371-378.
Direct Link - Sargisson, R.J., I.G. Mclean, G.S Brown and K.G. White, 2007. Seasonal variation in pigeon body weight and delayed matching-to-sample performance. J. Exp. Anal. Behav., 88: 395-404.
Direct Link - Simondon, K.B., T. Ndiaye, M. Dia, A. Yam and M. Ndiaye et al., 2007. Seasonal variations and trends in weight and arm circumference of non-pregnant rural Senegalese women, 1990-1997. Eur. J. Clin. Nutr.
CrossRefDirect Link - Slob, A.K. and J.J.V.D.W.T. Bosch, 1975. Sex differences in body growth in the rat. Physiol. Behav., 14: 353-361.
CrossRef - Stillwell, R.C., W.U. Blanckenhorn, T. Teder, G. Davidowitz and C.W. Fox, 2010. Sex differences in phenotypic plasticity affect variation in sexual size dimorphism in insects: From physiology to evolution. Ann. Rev. Entomol., 55: 227-245.
Direct Link - Susic, V., V. Pavic, B. Mioc, I. Stokovic and A.E. Kabalin, 2005. Seasonal variations in lamb birth weight and mortality. Vet. Arh., 75: 375-381.
Direct Link - Tamate, T. and K. Maekawa, 2006. Latitudinal variation in sexual size dimorphism of sea-run masu salmon, Oncorhynchus masou. Evolution, 60: 196-201.
PubMedDirect Link - Zenuto, R.R., 1999. Sexual size dimorphism, testes size and mating system in two populations of Ctenomys talarum (Rodentia: Octodontidae). J. Nat. History, 33: 305-314.
Direct Link