
Sabry A. El-Naggar
Department of Zoology, Faculty of Science, Tanta University, Tanta, Egypt
Ismail M. Al-Sharkawi
Department of Zoology, Faculty of Science, Tanta University, Tanta, Egypt
Gamal A. Madkour
Department of Zoology, Faculty of Science, Tanta University, Tanta, Egypt
International Journal of Zoological Research
Year: 2011 | Volume: 7 | Issue: 5 | Page No.: 358-368







ABSTRACT
The most important definitive host of Schistosoma mansoni is the human, however, numerous other mammalian species were found to be infected with this parasite. Among these species, the wild rodents are the most common. In this study, the susceptibility of some wild rodents widely distributed in Egypt to S. mansoni infection was evaluated. Five wild species were tested for the susceptibility of S. mansoni infection in vitro; including Mus musculus (black mice), Acomys cahirinus (Cairo spiny mice), Rattus rattus (house rats), Rattus norvegilcus (Norway rats), Rattus norvegicus (albino rats) and Arvicanthis niloticus (Nile rats). Laboratory mice were used as a positive control. Rodents were infected individually with 150 S. mansoni cercariae by tail immersion and housed for 8 week post-infection. The results reported that the Nile rats showed the highest number of worm burden (90 worms), while the Norway rats and the laboratory rats showed the lowest numbers among the tested species. This study also showed that the Nile rats, the house rats and the Norway rats yielded high number of eggs in the liver tissues. In contrast, the Cairo spiny mice, the black mice and albino rats yielded low number of eggs in the liver tissue. As compared to the permissive host albino mice, the Nile rats, the black mice, the Cairo spiny mice and the house rats showed comparable granuloma size. In contrast, albino rats and Norway rats showed a small granuloma size. Alltogether, these data showed that S. mansoni infection to these wild rodents was species dependant.
PDF Abstract XML References Citation
Received: June 08, 2011;
Accepted: September 20, 2011;
Published: December 07, 2011
How to cite this article
Sabry A. El-Naggar, Ismail M. Al-Sharkawi and Gamal A. Madkour, 2011. Susceptibility of Some Wild Rodents Widely Distributed in Egyptian Foci to Schistosoma mansoni Infection under Laboratory Conditions. International Journal of Zoological Research, 7: 358-368.
DOI: 10.3923/ijzr.2011.358.368
URL: https://scialert.net/abstract/?doi=ijzr.2011.358.368
DOI: 10.3923/ijzr.2011.358.368
URL: https://scialert.net/abstract/?doi=ijzr.2011.358.368
INTRODUCTION
The schistosomiasis is endemic in tropical and subtropical areas representing the second in importance to malaria from the socioeconomic and public health standpoints (Sturrock, 2001). It is endemic in 74 developing countries and afflicts between 200 and 300 million people worldwide (Chippaux et al., 1997; Sturrock, 2001; Stothard et al., 2006). Over 600 million people are at risk because of poverty, ignorance and poor housing with little or no sanitation (Cheever et al., 1993; Chitsulo et al., 2000). Although, several natural compounds were tested to treat Schistosomiasis (Soliman and El-Shenawy, 2003; Aly et al., 2006; Abdel-Aziz et al., 2011; Karim, 2011). With all trials to develop anti-schistosmal drug, Praziquantel (PZQ) is still the effective for the optimal treatment (Adam, et al., 2005). Recently, it has been shown that praziquantel-resistant parasites may be emerged due to the continuous treatment with PZQ alone (Danso-Appiah and De-Vlas, 2002). In Egypt, in particular, as a result of the construction of Aswan High Dam, an increase in the prevalence of schistosomiasis has been found and as a direct effect of this Dam, S. mansoni replaced S. haematobium along the River Nile (Abdel-Wahab et al., 1993). The risk of infection in Egypt is still high because of S. haematobium and S. mansoni are well distributed in the endemic areas of delta (El-Khouby et al., 2000), middle and upper Egypt (Hammam et al., 2000). Furthermore, in the last ten years, the epidemiology of schistosomiasis has been extensively investigated, whereas the distribution of schistosomiasis is well documented for most affected areas in Egypt (Gabr et al., 2000; Habib et al., 2000; Hammam et al., 2000; Nooman et al., 2000). As a result of complex water canal irrigation system in Egypt, most of children and farmers were easily infected with the S. mansoni and S. haematobium (Abu-Elyazeed et al.,1998; Barakat et al., 2000).
It is well known that man is the most important definitive host of S. mansoni for maintenance of the life cycle, however, several studies showing that other mammals may contribute in maintaining the life cycle as well (De Amorim, 1953; Barbosa et al., 1962; Barbosa, 1972; Silva and Andrade, 1989; Costa-Silva et al., 2002). In this line, it has reported that most of the mammals with natural infection with S. mansoni were rodents, chimpanzee (Barbosa, 1972; Mansour, 1973a; Renquist et al., 1975; Kawazoe et al., 1978; Maldonado et al., 1994; Morand et al., 1999; Gentile et al., 2006). In Egypt foci, it has found that, the gerbil; Gerbillus g. pyramidium, the Nile rats and the field rats were naturally infected with both of S. mansoni and S. haematobium (Mansour, 1973b). Similarly, in Brazil, it has found that the water-rats; Nectomys squamipes may present different levels of importance in the transmission dynamics of S. mansoni infection and could be considered important wild-reservoirs of this disease (Rodrigues e Silva et al., 1992; D’Andrea et al., 2002; Gentile et al., 2006). In Argentina, northeast on the left river-bank of Parana River, two Olygoryzomys flavescens, two Holochilus braziliensis and one Scapteromys tuncidus were captured and exposed to infection with 40 cercariae per animal. These rodents were able to eliminate viable eggs and hatched miracidiae able to infect the intermediate host of S. mansoni (Borda and Rea, 2006). Not only the rodents have been found to be infected naturally with S. mansoni, but also certain domestic animals such as cattle were capable of transmitting the cycle of infection even human transmissions were controlled (Barbosa et al., 1962). This study aimed to evaluate the susceptibility of some wild rodents distributed widely around the water canal system in Egypt to S. mansoni infection under the laboratory conditions. The study showed that collected species of wild rodents such as Cairo spiny mice, black mice (Sisi), house rats and the Nile rats were susceptible for S. mansoni infection while other species such as Norway rats were resistance to the infection.
MATERIALS AND METHODS
This study was conducted at the Department of Zoology, Faculty of Science, Tanta University, Egypt. Wild rodents were collected from different foci in the Nile Delta; the experiments were done approximately in 2 years.
Laboratory and wild rodents: Adult Swiss albino mice weighting 20±2 g and albino rat were purchased from Teodor Bilharis Institute (TBI), Imbaba, Giza, Cairo, Egypt). Different species of the wild rodents were collected from their foci and transported into the animal facility, Zoology Department. Before starting the experiments, all the animals were acclimatized to the laboratory conditions for 2 weeks and tested for S. mansoni or other parasite infections by stool analysis. Animals were maintained on laboratory standard balanced diet.
Wild rodents under the experimentation: In addition to the laboratory mice (Mus musculus) and the laboratory rats (Rattus rattus), five wild rodent species (8 animals/group) were used in this study including; black mice (Mus musculus), Cairo spiny mice (Acomys cahirinus), house rats (Rattus rattus), Norway rats (Rattus norvegilcus) and the Nile rats (Arvicanthis niloticus).
Cercarial infection and enumeration of worm burden: S. mansoni cercariae were obtained from laboratory infected Biomphalaria alexandrina snails supplied by the Schistosome Biological Supply Program, Theodore Bilharz Research Institute at Warrak EL-Hadar, Imbaba, Cairo, Egypt. The cercariae were collected from the infected snails according the method described by Christensen et al. (1982). The infection with S. mansoni cercariae and the perfusion technique which used in this study were based on Christensen et al. (1982).
Determination of quantitative oograms: For egg count, glass slides and cover slips were weighed on a balance of 0.1 mg sensitivity. Samples of tissue fragment of each liver of infected rodents with S. mansoni were transferred carefully into the previously weighed slides. The weight of each fragment was then calculated. The slides were then microscopically examined under low magnification. All viable eggs were counted according to the criteria as previously described by Pellegrino and de Maria (1966).
Histological preparation: After liver perfusion, the livers were excised, removed, blotted using filter paper and weighed. For histo-pathological preparation, selected liver specimens were fixed in 10% neutral formalin for 24 h. Tissue specimens were then washed in running tap water for 12 h, dehydrated in standard alcoholic series and cleared in xylen before embedding in paraffin wax. Samples were then sectioned at thickness of 6 microns and stained by the conventional H and E method.
Granuloma size measurements: Lesions containing eggs in their centers were selected for measurement. The diameter of each liver granuloma was obtained by measuring two diameters of the lesion at right angles to each other using an ocular micrometer. The mean diameter of at least 50 lesions from each animal of each group was determined. The volume of each lesion was calculated using the following formula: Volume = R x 22/7 x 4/3, where R is the radius of the garnuloma, according to Cheever et al. (1993).
Statistical analysis: All values were expressed as Mean±SD. Statistical analysis was performed by student’s t-test. The p values <0.05 were considered as significant when compared to control.
RESULTS
Infection of wild rodents with S. mansoni was species dependant: To test whether the captured wild rodents; black mice, cairo spiny mice, norway rats, house rats and the nile rats were able to be infected with S. mansoni cercariae. These rodents were infected under the laboratory conditions with 150 S. mansoni cercariae/animal and housed for 8 weeks post- infection. The results showed that compare to the permissive host the laboratory mice (46.0 worms/mouse), the black mice (Sisi) showed almost the same numbers of worm burden. The Cairo spiny mice and the house rats, however, showed slightly low number of worm burden, while the Nile rats showed the highest number of the worm burden (90 worms). In contrast, the Norway and the laboratory rats showed a significant decrease in the total worm burden (Fig. 1, 2).
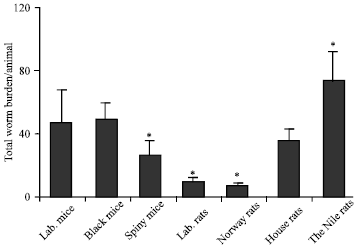 | |
| Fig. 1: | Wild rodents infected with S. mansoni cercariae under the laboratory conditions. The wild rodents infected with 150 S. mansoni cercariae/animal in vitro then, hosted for 8 weeks under the laboratory conditions. Rats and mice were sacrificed, liver perfusion performed to collect the worms, also worms were picked from the intestinal mesenteries around the alimentary canal. The worms were separated and counted under the dissected microscope in each group. *Differences are statistically significant from the control group at p<0.05 |
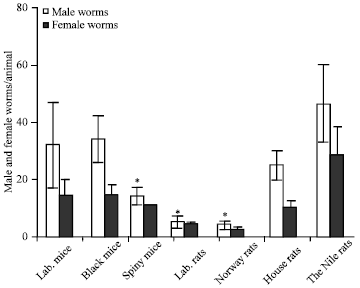 | |
| Fig. 2: | Male and female S. mansoni worms were found in the wild animals infected under the artificial conditions. The wild rodents were infected with 150 S. mansoni cercariae/head, then, hosted for 8 weeks under the laboratory conditions. Rats and mice were sacrificed, liver perfusion were done to collect the worms, also the worms were picked from the intestinal mesenteric. The worms were separated and differentiated into males and females in each group under the dissecting microscope. *Differences are statistically significant from the control group at p<0.05 |
In all infected wild rodents, the number of the adult male worms was higher than the number of the adult females. The number of the male Schistosoma worms in the laboratory mice, black mice and the Cairo spiny mice were 32, 36 and 18, respectively.
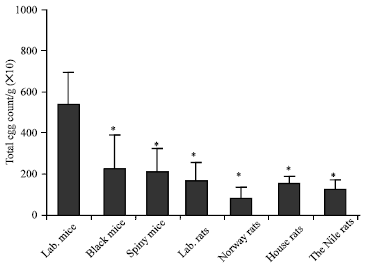 | |
| Fig. 3: | The permissive host laboratory mouse shows the highest number of schistosme eggs per gram of the liver tissues among the tested wild rodents. Rats and mice were sacrificed, and 100 mg of fragmented liver sample tissue were cut and mounted in slides, the total number of the S. mansoni eggs number were counted in each group. *Differences are statistically significant from the control group at p<0.05. |
While the total number of female worms in these species were 12, 12 and 10, respectively. The total number of male Schistosoma worms in the house rats, the Nile rats, the Norway and the laboratory rats were 25, 50, 4 and 5, respectively. The total numbers of female worms in these rodents were 9, 40, 3 and 4, respectively (Fig. 1, 2).
Infected wild rodents with S. mansoni showed viable and dead eggs in their liver tissues: To test whether wild rodents able to produce S. mansoni eggs in their liver tissues after infection with S. mansoni, fragmented samples of liver tissues (0.1 g) were used. The results showed that compared to the permissive host; the laboratory mice, other wild rodent species showed less number of eggs in their liver tissues. The black mice and Cairo spiny mice showed less than the half of the egg counts in the liver of the permissive host. The house rats, laboratory rats and the Nile rats showed nearly one- third the number of liver eggs in the permissive laboratory mice, while the Norway rats showed the lowest liver egg count (Fig. 3). The results showed that most of the eggs which counted in the liver fragmented samples of the Cairo spiny mice, Norway rats and the laboratory rats were dead. On contrast, the majority of the counted eggs in the liver tissues of black mouse and the Nile rats were live similar to those in the permissive host albino mice (Fig. 4). Although, the highest number of the total egg count per gram was found in the permissive host albino mice, the Nile and house rats showed the higher numbers of the total egg counts due to their relative high liver weights while the black mice showed the lowest number of eggs due to their small liver weights (Fig. 5).
Varied in granuloma sizes according the wild rodent species: To test weather the infected rodents able to form a granulomatus reaction in the liver due to egg deposition, liver of each group were fixed in 10% neutral formalin for 24 h and stained with H and E and examined under the microscope to measure the granuloma sizes. The data showed that the liver sections of the infected laboratory mice had the typical characteristic granulomatus reactions. These granulomas were formed mainly of histocytes, lymphocytes and eosinophils. Bilharzial pigments are also observed in Kupffer’s cells (Fig. 7).
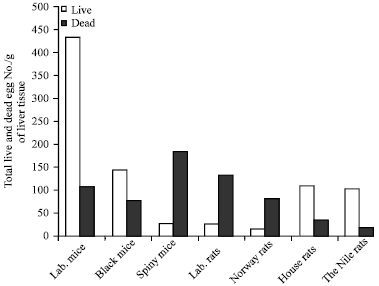 | |
| Fig. 4: | The permissive laboratory mice, black mice, house rat and Nile rat showed high viable eggs while laboratory rat, Cairo spiny mouse, and Norway rat showed high dead ones. Rats and mice were sacrificed, 100 mg of liver tissue were cut and liver fragmented samples were mounted in slides, then, the numbers of the live and dead eggs were counted in each group |
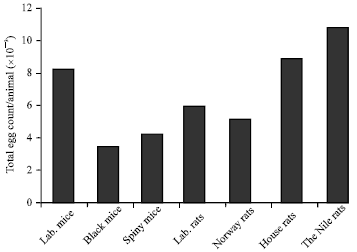 | |
| Fig. 5: | The Nile rat showed the maximum egg load in their livers. The livers from each group weighted and then 100 mg were taken and mounted in clear histological slides, the total S. mansoni eggs (live and dead) were counted in each group, then the total egg numbers per livers were calculated. |
The average granuloma volume in the permissive host was 120±71 mM3. In addition, the granuloma volume in all other tested species was smaller than those of the permissive laboratory mice except for the Nile rats which showed the highest granuloma volume. The average of granuloma size in the liver sections of the Nile rat was 145±68 mM3 (Fig. 6, 7). The liver sections of infected black mice, Cairo spiny mice and the house rats showed the characteristic granulomatus reaction like that in the permissive host (laboratory mice) and the average granuloma size were 45±27 mM3, 83±47 mM3 and 70±5 mM3, respectively. The smallest granuloma size was recorded in Norway rats and in the laboratory rats where the average of the granuloma sizes were 7±5 mM3 and 9±8 mM3, respectively (Fig. 6, 7).
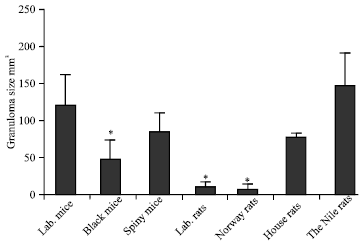 | |
| Fig. 6: | The Nile rats show the biggest granuloma size among the tested wild rodent species.. Tested wild rodents were sacrificed. The livers from each group were fixed in 10% neutral formalin and stained with H and E to count and measure the granulomatus reaction in the liver by using the ocular micrometer. *Differences are statistically significant from the control group at p<0.05 |
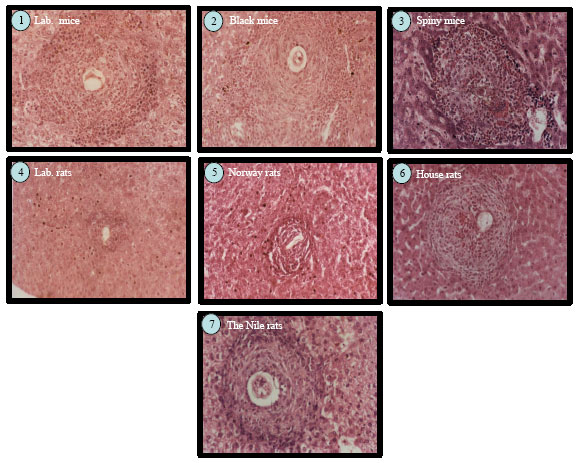 | |
| Fig. 7: | Laboratory rat R. norvigecus and wild Norway rats show the smallest granuloma size amount the tested wild rodents. The tested wild rodents were sacrificed. The livers from each group were fixed in 10% neutral formalin and stained with H and E to count and measure the granulomatus reaction in the liver |
DISCUSSION
Human is considered the main host for the maintenance and the transmission of S. mansoni cycle, however, other vertebrate hosts may be able to transmit the infection (De Noya et al., 1997; Gentile et al., 2006). In this study, it has found that some wild rodent species which are naturally distributed around the irrigation canal systems in Egyptian foci were susceptible to S. mansoni infection under the artificial conditions. The collected wild rodent species showed variability in their susceptibility to S. mansoni (Fig. 1). This variation could be because some rodents were permissive and some others were resistance to S. mansoni infection. In this study, two new species of wild rodents were evaluated to S. mansoni infection; the black mice (Sisi) and the Cairo spiny mice. Other wild rodent species were previously recorded to be naturally infected with S. mansoni. Among the tested wild rodents, the Nile rats which distributed allover the water canal irrigation systems in Egypt have found to be infected with a high number of the schistosoma worms. This result could explain and support what Arafa and Massoud (1990) has found in their studies. They have reported that the Nile rats have found naturally infected with both S. mansoni and S. haematobium (Mansour, 1973a; Arafa and Massoud, 1990). Due to the Nile rats are distributed with a large numbers near the canals of the irrigation system, this could indicate that these rodents might play a role in S. mansoni transmission. The results showed that the Cairo spiny mice, black mice and house rats were found to be infected with S. mansoni similar to the laboratory mice (Fig. 2). Based on this, the tested wild rodents might help in the transmission of S. mansoni infection especially in the regions which contains into a large numbers of these rodents. It has been reported that compared to the Nile rats, the house rats spends little time near the water, but also could be a good reservoir to S. mansoni parasite (Jourdane and Imbert-Establet, 1980). In this study, the Norway rats and the laboratory rats have found to be infected with few number of the schistosoma worms (Fig. 1, 2) therefore, these rodents might be considered as non-permissive hosts to S. mansoni infection. This finding is in agreement other study which showed that the Norway and house rats got the infection with S. mansoni, but did not participate in the transmission of the schistosomiasis (De Noya et al., 1997). In this study, it has found that the number of the eggs was directly proportional to the number of the worm burden except in the Nile rat (Fig. 3). Even, the number of eggs per gram of tissue was high at the permissive laboratory mice however, the Nile rats and the house rats showed the highest number of eggs per animal (Fig. 5) due to their relative high liver weights. The results showed that both of Norway rats and the laboratory rats showed a similar number of eggs while the black mouse showed the lowest number of eggs per animal (Fig. 3). The Nile rats, the black mice and the house rats showed large number of live eggs than the dead ones. In contrast, the Cairo spiny mice, the Norway rats and the laboratory rats showed dead eggs more than the live ones (Fig. 4). These results might be support that the wild rodents which have more viable eggs in their tissues might be participate in the parasite transmission more than those with more dead eggs in their tissues. The current data in agreement with a previous study which reported that house rats eliminate fertile eggs containing infectious miracidia of S. mansoni while the Norway rats and in the white rats, the eggs of S. mansoni were not fertile and were never eliminated by the host (Jourdane and Imbert-Establet, 1980)
Infection of wild rodents with S. mansoni at a rate of infection of 150 cercariae/animal for 8-weeks caused marked degenerative changes in the liver tissue. The histological examination of the liver showed that the main histological degenerative changes were confined to the granuloma formation in the portal tract in all the tested wild rodents. The most prominent changes included granuloma formation. In all the tested wild rodent species, the granulomatous reaction was similar, except less sever in the Norway rats and the laboratory rats. The granuloma size in the Norway rats and laboratory rats were small as compared to the permissive host; Laboratory mouse (Fig. 5, 6). These reductions in the granuloma size may be attributing to the passion of these species to some degree of natural resistance to S. mansoni infection (Bout et al., 1986).
CONCLUSION
Considering the agricultural expansion into new areas in Egypt, there is an urgent need to make detailed studies on role of the wild rodents and domesticated animals on the maintenance and the transmission of the schistosomiasis. In conclusion, there is an urgent need to increase the studies determine the possible roles of the wild rodents in the transmission and maintenance of the life cycle of S. mansoni.
REFERENCES
- Abdel-Wahab, M.F., A. Yosery, S. Narooz, G. Esmat, S. El-Hak, S. Nasif and G.T. Strickland, 1993. Is Schistosoma mansoni replacing Schistosoma haematobium in the fayoum? Am. J. Trop. Med. Hyg., 49: 697-700.
PubMedDirect Link - Abdel Aziz, I.Z., E.A. Ayman and H.E. Sabah, 2011. In vitro anti-schistosomal activity of Plectranthus tenuiflorus on Miracidium, cercaria and schistosomula stages of Schistosoma mansoni. Res. J. Parasitol., 6: 74-82.
CrossRef - Abu-Elyazeed, R.R., N.S. Mansour and J.R. Campbell, 1998. The prevalence and intensity of Schistosoma mansoni infection among residents in two Egyptian villages, at two and twelve months post-treatment with 40 and 60 mg praziquantel per kg body weight at these two villages respectively. J. Egypt. Soc. Parasitol., 28: 9-16.
PubMedDirect Link - Aly, S.A., N.S. El-Rigal and M.Z. Rizk, 2006. Nutritional supplementation with Ailanthus altissima and Ziziphus spina christi to compensate for some metabolic disorders in Schistosoma mansoni infected mice. Pak. J. Biol. Sci., 9: 1700-1706.
CrossRefDirect Link - Adam, I., E. Elwasila and M. Homeida, 2005. Praziquantel for the treatment of schistosomiasis mansoni during pregnancy. Ann. Med. Parasitol., 99: 37-40.
PubMed - Karim, A., 2011. Use of natural products as therapeutic agents for the treatment of schistosomiasis. Asian J. Anim. Vet. Adv., 6: 863-864.
CrossRef - De Noya, B.A., J.P. Pointier, C. Colmenares, A. Theron and C. Balzan et al., 1997. Natural Schistosoma mansoni infection in wild rats from Guadeloupe: Parasitological and immunological aspects. Acta Trop., 68: 11-21.
CrossRefPubMedDirect Link - Arafa, M.A. and M.M. Massoud, 1990. Natural Schistosoma mansoni infection in Arvicanthis niloticus in Ismailia, Egypt. J. Egypt. Soc. Parasitol., 20: 775-778.
PubMedDirect Link - Barakat, R., A. Farghaly, A.G. El-Masry, M.K. El-Sayed and M.H. Hussein, 2000. The epidemiology of schistosomiasis in Egypt: patterns of Schistosoma mansoni infection and morbidity in Kafer El-Sheikh. Am. J. Trop. Med. Hyg., 62: 21-27.
PubMedDirect Link - Barbosa, F.S., I. Barbosa and F. Arruda, 1962. Schistosoma mansoni: Natural infection of cattle in Brazil. Science, 138: 831-831.
CrossRefPubMedDirect Link - Barbosa, F.S., 1972. Natural infection with Schistosoma mansoni in small mammals trapped in the course of a schistosomiasis control project in Brazil. J. Parasitol., 58: 405-407.
PubMed - Borda, C.E. and M.J. Rea, 2006. Intermediate and definitive hosts of Schistosoma mansoni in Corrientes province, Argentina. Memoirs Inst. Oswaldo Cruz, 101: 233-234.
CrossRefPubMedDirect Link - Bout, D., M. Joseph, M. Pontet, H. Vorng, D. Deslee and A. Capron, 1986. Rat resistance to schistosomiasis: Platelet-mediated cytotoxicity induced by C-reactive protein. Science, 231: 153-156.
CrossRefDirect Link - Cheever, A.W., I.A. Eltoum, Z.A. Andrade and T.M. Cox, 1993. Biology and pathology of Schistosoma mansoni and Schistosoma japonicum infections in several strains of nude mice. Am. J. Trop. Med. Hyg., 48: 496-503.
PubMedDirect Link - Chippaux, J.P., D. Boulanger, P. Bremond, G. Campagne, C. Vera and B. Sellin, 1997. The WHO collaborating centre for research and control of schistosomiasis at Niamey, Niger. Memoirs Inst. Oswaldo Cruz, 92: 725-728.
PubMedDirect Link - Chitsulo, L., D. Engels, A. Montresor and L. Savioli, 2000. The global status of schistosomiasis and its control. Acta Tropica, 77: 41-51.
CrossRefPubMedDirect Link - Christensen, N.O., F. Frandsen, P. Nansen and A. Schultz, 1982. A simple restraining chamber for exposing mice to schistosome cercariae by the tail immersion technique. J. Parasitol., 68: 504-504.
PubMedDirect Link - Costa-Silva, M., R. Rodrigues-Silva, M. Hulstijn, R.H. Neves, M. de Souza Panasco, H.L. Lenzi and J.R. Machado-Silva, 2002. Natural Schistosoma mansoni infection in Nectomys squamipes: Histopathological and morphometric analysis in comparison to experimentally infected N. squamipes and C3H/He mice. Memoirs Inst. Oswaldo Cruz, 97: 129-142.
PubMed - D'Andrea, P.S., F.A. Fernandes, R. Cerqueira and L. Rey, 2002. Experimental evidence and ecological perspectives for the adaptation of Schistosoma mansoni Sambon, 1907 (Digenea: Schistosomatidae) to a wild host, the water-rat, Nectomys squamipes Brants, 1827 (Rodentia: Sigmodontinae). Memoirs Inst. Oswaldo Cruz, 97: 11-14.
PubMedDirect Link - Danso-Appiah, A. and S.J. de Vlas, 2002. Interpreting low praziquantel cure rates of Schistosoma mansoni infections in Senegal. Trends Parasitol., 18: 125-129.
CrossRefPubMedDirect Link - De Amorim, J.P., 1953. Experimental and natural infection of rodents with Schistosoma mansoni; preliminary note. Rev. Bras. Malariol. Doencas Trop., 5: 219-222.
PubMedDirect Link - El-Khoby, T., N. Galal, A. Fenwick, R. Barakat and A. El-Hawey et al., 2000. The epidemiology of schistosomiasis in Egypt: Summary findings in nine governorates. Am. J. Trop. Med. Hyg., 62: 88-99.
PubMedDirect Link - Gabr, N.S., T.A. Hammad, A. Orieby, E. Shawky, M.A. Khattab and G.T. Strickland, 2000. The epidemiology of schistosomiasis in Egypt: Minya Governorate. Am. J. Trop. Med. Hyg., 62: 65-72.
PubMedDirect Link - Gentile, R., S.F. Costa-Neto, M.M. Goncalves, S.T. Bonecker and F.A. Fernandes et al., 2006. An ecological field study of the water-rat Nectomys squamipes as a wild reservoir indicator of Schistosoma mansoni transmission in an endemic area. Memoirs Inst. Oswaldo Cruz, 101: 111-117.
PubMedDirect Link - Habib, M., F. Abdel Aziz, F. Gamil and B.L. Cline, 2000. The epidemiology of schistosomiasis in Egypt: Qalyubia Governorate. Am. J. Trop. Med. Hyg., 62: 49-54.
PubMedDirect Link - Hammam, H.M., A.H. Zarzour, F.M. Moftah, M.A. Abdul-Aty and A.H. Hany et al., 2000. The epidemiology of schistosomiasis in Egypt: Qena Governorate. Am. J. Trop. Med. Hyg., 62: 80-87.
Direct Link - Jourdane, J. and D. Imbert-Establet, 1980. Experimental study of the permissivity of the wild rat (Rattus rattus) of Guadeloupe with regard to Schistosoma mansoni. Hypothesis on the role of this host in the dynamics of natural habitats Acta Trop., 37: 41-51.
PubMedDirect Link - Kawazoe, U., L.C. Souza Dias and T.Piza J'de, 1978. Natural infection of small mammals by Schistosoma mansoni, in the Americana reservoir (Sao Paulo, Brazil). Rev Saude Publica., 121: 200-208.
PubMedDirect Link - Maldonado, A., Jr., J.R. Machado e Silva, R. Rodrigues e Silva, H.L. Lenzi and L. Rey, 1994. Evaluation of the resistance to Schistosoma mansoni infection in Nectomys squamipes (Rodentia: Cricetidae): A natural host of infection in Brazil. Rev. Inst. Med. Trop., 36: 193-198.
PubMed - Mansour, N.S., 1973. Schistosoma mansoni and S. haematobium found as a natural double infection in the Nile rat, Arvicanthis n. niloticus, from a human endemic area in Egypt. J. Parasitol., 59: 424-424.
PubMed - Mansour, N.S., 1973. Schistosoma mansoni and Schistosoma haematobium natural infection in the Nile-rat, Arvicanthis n. niloticus from an endemic area in Egypt. J. Egypt Public Health Assoc., 48: 94-100.
PubMedDirect Link - Morand, S., J.P. Pointier and A. Theron, 1999. Population biology of Schistosoma mansoni in the black rat: Host regulation and basic transmission rate. Int. J. Parasitol., 29: 673-684.
CrossRefPubMedDirect Link - Nooman, Z.M., A.H. Hasan, Y. Waheeb, A.M. Mishriky and M. Ragheb et al., 2000. Egypt: Ismailia governorate. Am. J. Trop. Med. Hyg., 62: 35-41.
PubMed - Pellegrino, J. and M. de Maria, 1966. Results of exposing mice to natural pond water harboring a colony of Australorbis glabratus highly infected with Schistosoma mansoni. Am. J. Trop. Med. Hyg., 15: 333-336.
PubMed - Renquist, D.M., A.J. Johnson, J.C. Lewis and D.J. Johnson, 1975. A natural case of Schistosoma mansoni in the chimpanzee (Pan troglodytes versus). Lab. Anim. Sci., 25: 763-768.
PubMedDirect Link - Silva, R.R., J.R. Machado e Silva, N.F. Faerstein, H.L. Lenzi and L. Rey, 1992. Natural infection of wild rodents by Schistosoma mansoni. Parasitological aspects. Memoirs Inst. Oswaldo Cruz, 87: 271-276.
CrossRefPubMedDirect Link - Silva, T.M. and Z.A. Andrade, 1989. Natural infection of wild rodents by Schistosoma mansoni. Memoirs Inst. Oswaldo Cruz, 84: 227-235.
CrossRefPubMedDirect Link - Soliman, M.F.M. and N.S. El-Shenawy, 2003. Evaluation of the protective effect of two antioxidative agents in mice experimentally infected with Schistosoma mansoni: Haematological and histopathological aspects. Pak. J. Biol. Sci., 6: 887-897.
CrossRefDirect Link - Stothard, J.R., N.B. Kabatereine, E.M. Tukahebwa, F. Kazibwe and D. Rollinson et al., 2006. Use of Circulating Cathodic Antigen (CCA) dipsticks for detection of intestinal and urinary schistosomiasis. Acta Trop., 97: 219-228.
CrossRefPubMedDirect Link - Sturrock, R.F., 2001. Schistosomiasis epidemiology and control: How did we get here and where should we go? Mem. Inst. Oswaldo Cruz, Reo de Janerio, 96: 17-27.
Direct Link