
Maha F.M. Soliman
Zoology Department, Faculty of Science, Suez Canal University, Ismailia, Egypt
Nahla S. El-Shenawy
Zoology Department, Faculty of Science, Suez Canal University, Ismailia, Egypt
Pakistan Journal of Biological Sciences
Year: 2003 | Volume: 6 | Issue: 10 | Page No.: 887-897







ABSTRACT
The effect of antioxidative agents, Nigella sativa oil and melatonin, in protection against the oxidative processes induced by schistosomiasis was studied on haematological parameters, granuloma formation and tissue reaction of liver, intestine and kidney of mice experimentally infected with Schistosoma mansoni. N. sativa oil (1.14 g kg-1 orally) or melatonin (3.55 mg kg-1 i.p.) was administrated daily for 30 days starting from 1st day of infection. Haematological results revealed that treatment with melatonin and N. Sativa oil caused significant decrease in total leukocyte count while, RBCs, Hb content and PCV were increased significantly with N. sativa- treatment as compared with infected and melatonin-treated group. Morphometeric and histopathological findings of liver revealed a modulation in the pathological profile of schistosomiasis by reducing the severity of histopathological changes and attenuating hepatic granuloma. N. sativa oil did not interfere on the granuloma evolution in intestine, although, melatonin markedly reduced the fibrotic areas. Also, these antioxidants markedly ameliorated most of the pathological changes occurred in kidney. The effects of the antioxidants may be attributed partly to their early administration before the stimulation of the immune system or due to a modulation in the immune response in general.
PDF Abstract XML References Citation
How to cite this article
Maha F.M. Soliman and Nahla S. El-Shenawy, 2003. Evaluation of the Protective Effect of Two Antioxidative Agents in Mice Experimentally Infected with Schistosoma mansoni: Haematological and Histopathological Aspects. Pakistan Journal of Biological Sciences, 6: 887-897.
DOI: 10.3923/pjbs.2003.887.897
URL: https://scialert.net/abstract/?doi=pjbs.2003.887.897
DOI: 10.3923/pjbs.2003.887.897
URL: https://scialert.net/abstract/?doi=pjbs.2003.887.897
INTRODUCTION
The tropical parasite Schistosoma mansoni causes granulomatous inflammation following lodging of eggs in portal capillaries (Gharib el al., 1999). Experimental schistosome infections of laboratory animals have frequently been used to model the anatomopathologic and pathophysiologic features of the infection in humans as well as for the study of immunity and treatment (Cheever et al., 2002).
Liver and intestinal fibrosis is a major sequel to granulomatous schistosomiasis mansoni mostly responsible for portal hypertension, formation of esophageal varices and intestinal bleeding of infected human and mice (Giboda and Smith, 1997, Bogers et al., 2000). Most pathology in schistosome infected animals is attributed to the host’s reaction to the eggs which is maximal by the 8th week of infection (Cheever et al., 2002). The toxic egg material destroys the host tissue cells and the antigenic material stimulates the development of large inflammatory reactions (granuloma) around the egg material. This granuloma is considered to serve as a protective barrier by sequestering the toxic and antigenic substances secreted continuously from Schistosoma eggs, thereby, preventing further diffusion of these harmful materials (Philips and Lammie, 1986).
Granulomas are composed principally of macrophages, eosinophils and lymphocytes with the proportion of cells varying in different organs (Weinstock and Boros, 1983a). Natural killer cells may comprise over 20% of cells in the granuloma (Remick et al., 1988). Mast cells are infrequent in 8th wk granulomas in most mouse strains and become more frequent in chronic infections (Weinstock and Boros, 1983b). Hepatic granulomas formed around mature eggs of S. mansoni undergo changes in size, appearance and cellular constitution with time (Andrade and Warren 1964). Granulomas in the intestine were observed to be already modulated in the ileum from the beginning, although modulation was occurred also in the colon and ileal Peyer’s patch (Weinstock and Boros 1981, 1983a; Jacobs et al., 1998).
Many approaches were applied in order to prevent or decreased liver fibrosis in schistosomiasis mansoni infections (Giboda and Smith, 1997). Intensive research in the area of immunological intervention brought many promising results for the limitation of extended fibrosis (Czaja et al., 1989; Cheever et al., 1994 and 1995; Wynn et al., 1995) but anthelmintic chemotherapy is still the only applicable treatment for liver scars in infected persons (Giboda and Smith, 1997).
A recent study has demonstrated that in schistosomiasis, a high rate oxidative processes occurs (Gharib et al., 1999) and it may be that the major component of the disease and the host’s response involves the production of reactive oxygen species (El-Sokkary et al., 2002). The role of N. sativa oil in protection against the oxidative processes induced by schistosomiasis and also its role as a remarkable antischistosomiasis has been reported (Mahmoud et al., 2002; El-Shenawy and Soliman, 2003). In addition it has been reported that administration of melatonin is highly effective in ameliorated the S. mansoni-induced perturbations in the liver function biomarkers (El-Shenawy and Soliman, 2003).
In view of these findings, this study was undertaken to evaluate the protective effect of N. sativa oil and melatonin as antioxidant agents against the schistosomiasis mansoni from hematological and histopathological point of view.
MATERIALS AND METHODS
Animals: Swiss albino male mice of CD1 strain (Theodor Bilharz Institute, Cairo, Egypt), weighing 20-22 g were used in this study. They were kept in plastic cages with wire covers controlled temperature and light environment and supplied ad labitum with a standard basal diet and water. The mice were infected transcutaneously by exposing them to 100 cercariae per mouse of S. mansoni (Egyptian strain).
Experimental Groups and Treatment: Mice were randomly classified into four equal groups of eight mice each. Group I was control mice. Group II was the infected untreated mice and served as positive control. In group III, mice were treated with N. sativa (0.1 ml) orally at a dose level of 1.14 g kg–1body weight for 30 consecutive days based on the hepatoprotective dose recommended by EL-Dakhakhny et al. (2000), starting from the first day of infection. In group IV, mice were given melatonin (0.1 ml) at a dose of 3.5 mg kg–1 i.p. (Uz et al., 1997) for 30 days from the first day of infection. All animals were sacrificed 63 days post infection (PI).
Haematological studies: Blood samples used for haematological analysis were collected using capillary tubes introduced into the medial retro-orbital venous plexus (Sanford, 1954). Blood was collected into tubes containing an anticoagulant, ethylene diamine tetra-acetic acid (EDTA).
Red blood cell (RBCs) and total leucocytes (TLC) counts were measured according to Hall and Malia (1991). Hemoglobin (Hb)% and hematocrit value (PCV) were measured as described by Dacie and Lewis (1991). The absolute values of erythrocyte indices, namely mean cell volume (MCV), mean cell hemoglobin (MCH) and mean cell hemoglobin concentration (MCHC) were calculated according to Hall and Malia (1991). Differential leucocytes counts were determined on leishman stained blood smears (Hall and Malia, 1991).
Morphometeric studies: From each animal, three paraffin sections of 5 μm thickness were prepared and stained with haematoxylin / eosin (H and E). The sections were 250 μm distant from the preceding sections to avoid measuring of the same granuloma. Only granulomas containing a single egg in their centers were measured. The mean diameter (μm) of each granuloma was obtained by measuring two diameters of the lesions at right angles to each other with the help of an ocular micrometer. 70 lesions from each group were measured and calculated according to Mahmoud and Warren (1974).
Mean diameter of single granuloma (μm) = Sum of two diameter / 2
For counting of granuloma, twenty fields of 1 mm2 slide–1 were analyzed by light microscopy and calibrated with a millimetric reticule; the mean number of granulomas was computed in 100 mm2 group–1 (Reis et al., 2001). Granulomas were classified according to Li Hsü et al. (1972).
Histopathological studies: Specimen of liver, kidney and intestine were immediately removed out from scarified mice, fixed in 10% buffered formalin, embedded in paraffin, sectioned at 5 μm thickness and stained with H and E for histopathological examination and Masson=s triple stain to visualize total tissue fibrosis (Frazier-Jessen and Kovacs, 1993).
Detection and visual evaluation of hepatic and intestinal glycogen was carried out histochemically by using Periodic acid Schiff’s (PAS) technique for the demonstration of polysaccharide (Hotchkiss, 1948).
Statistical analysis: Statistical analysis was carried out using t-test and analysis of variance (ANOVA) according to Saunders and Tarpp (1990). To elucidate the effect of schistosomal infection, infected animals were compared with control groups while the comparison between infected and infected-treated groups was used to clarify the protective effect of the two anti-oxidant agents.
RESULTS
Table 1 showed the mean values of RBCs counts, Hb%, PCV, MCV, MCH and MCHC. Regarding RBCs count, Hb and PCV of infected mice revealed significant decrease by -43.4, -42.4 and -31.7% compared to uninfected control group, respectively. As a consequence to the alternation in RBCs count and PCV, the mean corpuscular volume (MCV) increased significantly by +18.8% compared to uninfected group. However, early treatment of mice with N. sativa resulted in a % increase of RBCs count, Hb% and PCV, by 25.5, 54.7 and 13.1% (p< .01) as compared with the infected group, respectively.
| Table 1: | Effect of N. sativa Oil and Melatonin on Red Blood Cells and Some Other Related Indices |
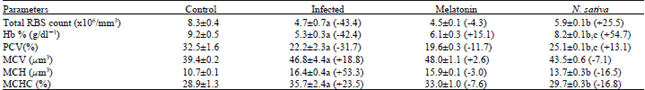 | |
| a Significantly different as compared with normal control P = 0.001. b Significantly different as compared with corresponding infected control P = 0.01. c Significantly different as compared with corresponding melatonin group P = 0.01. The percentage of change from the corresponding infected control is shown in parentheses. | |
| Table 2: | Effect of N. sativa Oil and Melatonin on Differential Leucocytes Count |
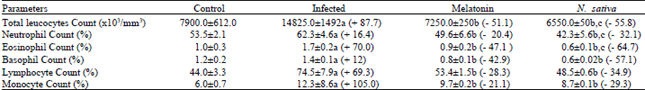 | |
| a Significantly different as compared with normal control P = 0.001. b Significantly different as compared with corresponding infected control P = 0.01. c Significantly different as compared with corresponding melatonin group P = 0.01. The % of change from the corresponding infected control is shown in parentheses. | |
In contrast, melatonin treatment had no significant effect on RBCs count, Hb% and PCV a s compared with the infected group (Fig. 1).
Due to the aforementioned, MCH and MCHC were significantly increased due to the schistosomal infection. However, the increment in MCH and MCHC were significantly reduced with N. sativa-oil treatment group. In contrast, the melatonin did not significantly alter MCH or MCHC (Table 1 and Fig. 2).
Table 2 revealed that the effect of melatonin and N. sativa oil on TLC and differential counts of infected mice with schistosomasis. Under the stress of schistosomal infection, the total leucocytes count was very highly significantly (P<0.001) elevated by 87.7% as compared with the control-uninfected mice. This leukocytosis was significantly decreased due to both N. sativa and melatonin treatment. As regards the absolute count of neutrophils and eosinophil, marked significant increments in its mean values were observed in the infected animals. When the infected mice were treated with the two antioxidant agents, the neutrophilia and eosinophil were significantly reduced. In association with mansonian schistosomiasis, the mean values of basophile were significantly elevated, while in mice treated with N. sativa oil or melatonin the mean was decreased by - 57.1 and - 42.9% as compared with infected mice. The mean value of lymphocytes count in infected animals was raised significantly by + 69.3%.
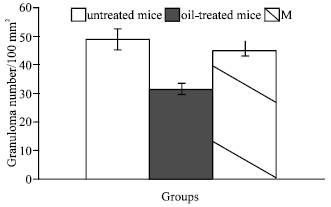 | |
| Fig. 1: | Effect of early treatment with N. sativa oil or Melatonin on Hepatic Granulomas number in S. mansoni infected mice |
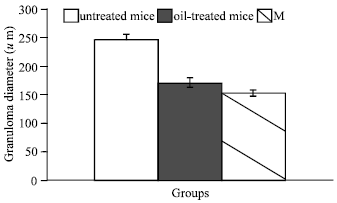 | |
| Fig. 2: | Reduction in Granuloma diameters in mice treated with N. sativa oil or Melatonin when compared with the infected-untreated one (0.001) |
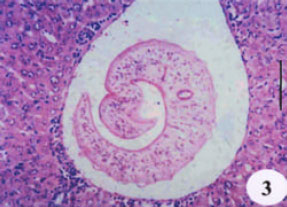 | |
| Fig. 3: | Liver section of mice infected with S. mansoni showing a worm in the hepatic portal vasculature with few tissue reactions around them |
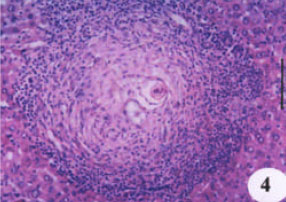 | |
| Fig. 4: | A productive-fibrotic hepatic granuloma showing the presence of eosinophils, macrophages and concentric disposition of collagen fibers |
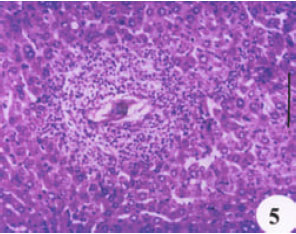 | |
| Fig. 5: | Liver section of mice treated with N. sativa shows a smaller granuloma with few cellular content and collagen fibers |
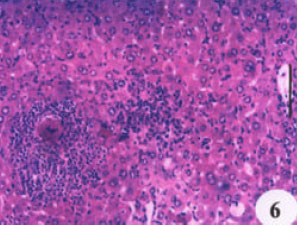 | |
| Fig. 6: | Liver section of mice treated with melatonin showing a markedly suppressive effect on granuloma formation, collagen fibers dispositions and eosinophilic infiltration. (H and E, X200) |
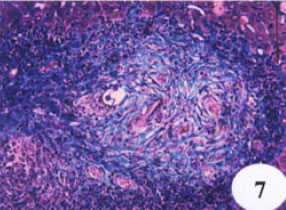 | |
| Fig. 7: | Liver sections of S. mansoni infected mice showing fibrosis in blue |
As the infected animals were treated with N. sativa oil or melatonin, lymphocytosis induced by infection was significantly decreased by -34.9 and -28.3, respectively (Fig. 3). In response to the infection, monocytes count increased significantly by +105% than Fig. 4 showed a productive-fibrotic granuloma surrounded by a layer of necrotic hepatocytes, with the presence of polymorphonuclear eosinophils, lymphocytes macrophages and disposition of thick collagen fibers. Granulomas in liver treated with N. sativa oil were differed from those of untreated-infected mice in showing a tendency to be smaller and with cellular content and collagen fibers becoming more packed (Fig. 5); eosinophils were less numerous as compared with untreated-infected liver sections. However, the liver sections of mice protected with melatonin showed appreciable suppressive effect on the granuloma formation (Fig. 6) as compared with infected liver sections. This suppressive effect was represented in smaller diameter of granuloma with few collagen fibers disposition and eosinophilic infiltrations.
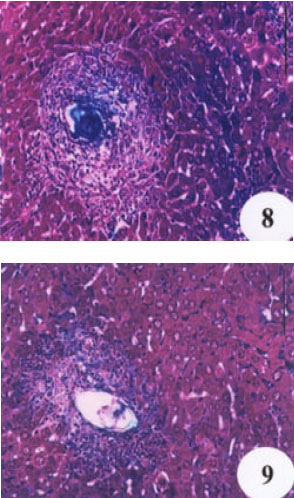 | |
| Fig. 8 and 9: | Liver section of mice treated with either N. sativa oil or melatonin, respectively, showing a minimal accumulation of collagen fibers. (Masson’s triple stain, X 250) |
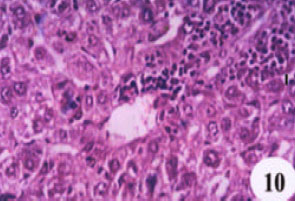 | |
| Fig. 10: | Liver section of mice infected with S. mansoni showing complete alteration of hepatic architecture with necrosis, fatty degeneration and a pronounced periportal cellular infiltration (X400, HandE) |
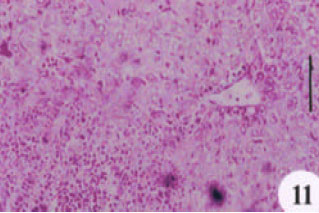 | |
| Fig. 11: | PAS-stained liver section showing a marked depletion in glycogen content |
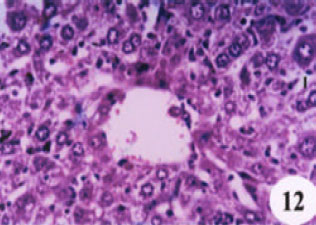 | |
| Fig. 12: | Liver section of mice treated with N. sativa showing no significant improvement in hepatic architecture (X400, HandE) |
Masson stained liver sections showed a minimal accumulation of collagen fibers in both N. sativa oil and melatonin-treated livers as compared with infected-untreated group (Fig. 7, 8 and 9). Focusing on the liver, there was a complete alteration of hepatic architecture which was represented in, variation of hepatocytes sizes, necrosis, cloudy swelling, fatty degeneration, presence of polymorphonuclei hepatocytes and a pronounced periportal inflammatory cellular infiltration around hepatic veins; eosinophils are the most inflammatory cells (Fig. 10). In addition, PAS stained liver showed highly depletion in glycogen content of the liver (Fig. 11). Treatment with oil, did not significantly improve either the liver architecture or the glycogen content as it affected on grnauloma itself (Fig. 12 and 13).
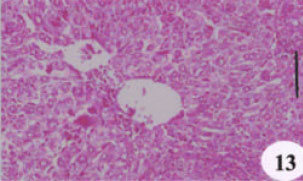 | |
| Fig. 13: | PAS-stained liver section showing also depletion in glycogen content where some hepatocytes reveal PAS negative reaction and few tiny sized glycogen particles observed near the cell margin of the other |
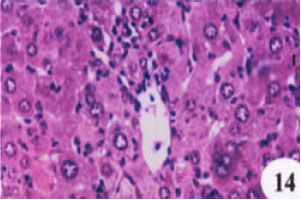 | |
| Fig. 14: | Liver section of mice treated with melatonin showing less evidence of the most of the pathological changes previously mentioned in untreated-infected mice (X400, HandE) |
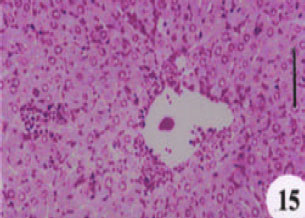 | |
| Fig. 15: | PAS-stained liver section showing moderate accumulations of glycogen seen as pink areas throughout the section. (X 200) |
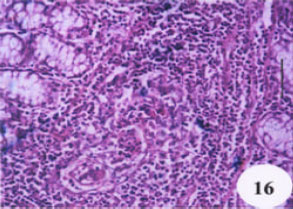 | |
| Fig. 16: | Intestinal section of S. mansoni infected mice showing granuloma formation around schistosoma eggs in submucosa of the intestine |
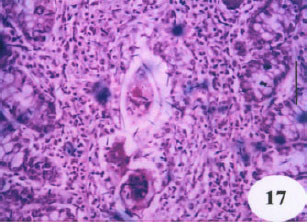 | |
| Fig. 17: | Intestinal section of mice treated with N. sativa showing no effect on granuloma formation |
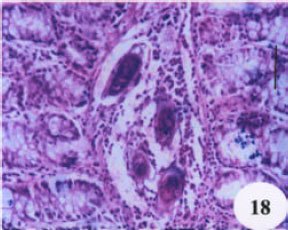 | |
| Fig. 18: | Liver section of mice treated with melatonin showing a noticeable suppressive on fibrosis and granuloma formation. (HandE X200) |
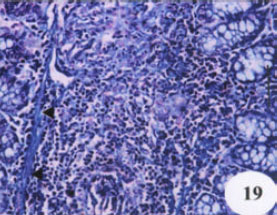 | |
| Fig. 19: | Intestinal sections of S. mansoni infected mice showing many fibrotic areas in blue extended as thin continuous bands (◀) |
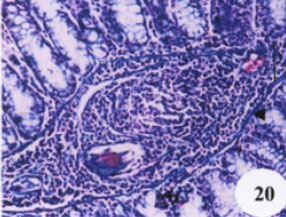 | |
| Fig. 20: | Intestinal section of mice treated with N. sativa oil showing many fibrotic areas in blue (◀) |
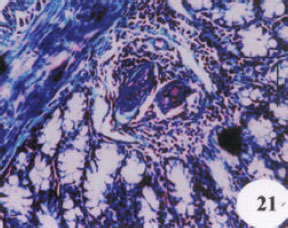 | |
| Fig. 21: | Intestinal section of mice treated with melatonin showing very few fibrotic areas (Masson’s triple stain, X 200) |
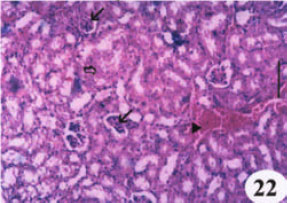 | |
| Fig. 22: | Kidney section of S. mansoni infected mice showing glomerulus congestion (↙), bleeding (◀) and degeneration of some tubules of kidney cortex (⇒) |
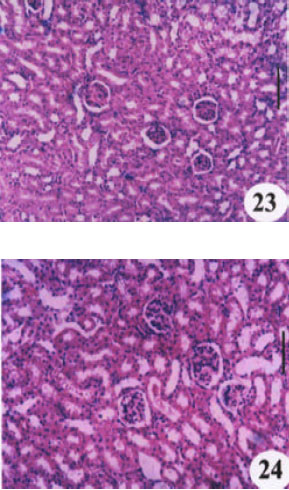 | |
| Fig. 23 and 24: | Kidney section of mice treated with either N. sativa oil or melatonin, respectively, showing amelioration of the previously mentioned changes (HandE, X 200) |
However, treatments with melatonin ameliorated most of the pathological changes previously mentioned in untreated-infected group and this enhancement was shown by the decrease in the numbers and area of necrotic foci with the less evidence of the previously mentioned degenerative change (Fig. 14). Meanwhile a significant improvement in the glycogen content was detected compared to the untreated groups (Fig. 15).
Histopathological findings revealed granulomas formation around schistosoma eggs in muscularis and submucosa of the intestine (Fig. 16). Granulomas showed mixture of productive and exudative features but it is generally smaller than liver granuloma. However, all components of extracellular matrix revealed in the liver were also present in granulomas of intestine, although always in smaller amount; granulomas predominantly consisted of eosinophilic, lymphocytes and macrophages. Treatment of infected mice with N. sativa oil had no suppressive effect on granuloma formation in the intestine compared to the infected-untreated mice (Fig. 17). In contrast, melatonin treatment showed a remarkable suppressive effect on collagen fibers and granuloma formation (Fig. 18). However, Masson’s stained liver sections revealed the formation of typical granuloma of intestine in both infected-untreated and N. sativa oil-treated mice (Fig. 19 and 20). The fibrosis pattern in intestine formed as thin continuous bands of collagen fibers in the submucosa while in liver, fibrosis was mainly detected inside granulomas. Melatonin treatment showed a remarkable suppressive effect on granuloma formation and on fibrosis as shown in Masson’s stained liver section (Fig. 21).
Moreover, histopathological study on kidney of mice infected with S. mansoni revealed the presence of glomerulus congestion, bleeding and degeneration with necrotic cells in some tubules of the kidney cortex (Fig. 22). Both oil and Melatonin administration after S. mansoni infection prevented most of the previously described changes (Fig. 23 and 24).
DISCUSSION
N. sativa oil has a remarkable anthelminthic activity associated with an enhancement in the antioxidant capacity. Meanwhile, melatonin treatment was highly effective in ameliorated the S. mansoni-induced perturbations in the liver function biomarkers (El-Shenawy and Soliman, 2003).
Data obtained in the present work revealed significant decrements in the mean values of RBC count, PCV and Hb content in infected animals. The severity of anemia was observed where the values of the three parameters were recorded being lower than the control. In conformity with the present findings, reduced RBC count, PCV and HB content were also recorded in S. mansoni-infected mice after 50 days (Ahmed, 1993; Abdel-Ghaffar and Qurtam, 2001). From an etiological point of view, anemia induced by S. mansoni infection may be attributed to blood loss and enhanced rate of hemolysis. Erythrocytes may be lost by two ways; from the bleeding induced by extrusion of ova through intestinal wall (Mahmoud and Woodruff, 1973) or due to consumption by adult schistosomes (Omeran, 1992; 887-897). As regards the latter possibility accounting for anemia, Gaafar et al. (1992) concluded that the plasma lipoprotein abnormalities in patients with hepatic schistosomiasis cause changes in lipid composition of erythrocyte membrane that in turn may be responsible, in part, for hemolysis and shortened life span of RBCs.
As the infected animals were treatment with N. sativa oil most of the previously described changes the infection—induced alternation in RBCs count, PCV, Hb content, MCV and MCH were remarkably improved. This improvement in red cell indices after treatment with N. sativa oil could indicate the stoppage of intestinal bleeding and loss off red cells after eradication of parasite. This suggestion seems coincident with the results of the parasitological that previously reported by El-Shenawy and Soliman (2003). Also, Zaoui et al. (2002) reported that therapeutic doses of N. sativa fixed oil had the ability to increase the Hb and PCV levels significantly of the normal mice and this could be the reason to improve the Hb and PCV levels of the infected mice in the present investigation. However, melatonin treatment group did not improve the RBCs count and the other related indices. But one of the obvious responses of infected mice to melatonin treatment was the change of Hb%. These finding are in agreement with a previous study (Tesoriere, 2001) which demonstrated the effect of melatonin on hemoglobin-derived oxoferryl radicals and inhibition of the hydroperoxide-induced hemoglobin denaturation in RBCs. Melatonin has been shown to act as a radical scavenger in various chemical and biological model systems in vitro. Kinetic evidence is provided showing that melatonin inhibits the irreversible degradation of hemoglobin (Hb), when incubated with RBCs (Tesoriere, 2001). Due to the aforementioned, it is clear that the N. sativa oil had more significant effect on RBCs, Hb and PCV of the infected mice than melatonin.
The present work exhibited remarkable increments in total leukocytes of S. mansoni-infected mice as compared with control animals. Shaker et al. (1980) attributed the increased counts of both total leukocytes and neutrophils to the stimulation of the cellular production. Ahmed (1993) and (Kuby, 1997) ascribed the increments in total leukocyte, neutrophils and eosinophil count to the powerful defense reaction and allergic manifestation against the schistosomes and/or their ova. It has been reported that eosinophilia is associated with schistosomiasis where the living ova and worms produce specific stimuli leading to bone marrow, blood and local tissue eosinophilia (El-Hawey et al., 1970).
Total and the differential counts of leukocytic categories reach to the normal level with treatment of infected mice with N. Sativa and melatonin. A decline in lecukocytes with N. Sativa treatment compared to control values is a generalized response pattern of mice (Zaoui et al., 2002). However, the role of melatonin, is may be due to their importance in the total antioxidative defense capacity for the organism.
As in schistosomiasis, GSH level in liver was drastically decreased, while hepatic lipid peroxide content increased as compared with the liver of uninfected mice (El-Shenawy and Soliman, 2003). These results are in line with granulomatous inflammation occurred in the liver. These findings show that the antioxidant capacity of the liver decreased, leading to the generation of lipid peroxides which may play a central role in the pathology associated with schistosomiasis as reported by Mahmoud et al. (2002).
Hepatic fibrosis due to Schistosoma infection is a complex process due to increased synthesis and deposition of extra cellular matrix components (Zero and Reid, 1993). However, in the present study early treatment against schistosomiasis in mice with N. sativa oil induced a noticeable degree of protection represented in less severe pathological changes in the liver, particularly the frequency of the inflammatory reactions; mediating both granuloma sizes and numbers. The decrease in the total number of hepatic granulomas could be related to the antischistosomal effect of N. sativa oil as previously demonstrated (Mahmoud et al., 2002; El-Shenawy and Soliman, 2003). It is worth mentioning that, early treatment with Melatonin, despite its negative effect as antischistosomal drug (El-Shenawy and Soliman, 2003), it had a remarkable suppressive effect on granuloma formation with a well noticeable improvement in glycogen content and architecture of liver and kidney. Previous study on experimental schistosomiasis in mice showed that treatment with N. sativa oil for 2 week starting from week 7 PI resulted in a smaller fibrocellular granuloma with less inflammatory cells (Mahmoud et al., 2002). In addition, El-Sokkary et al. (2002) reported that Melatonin administration after S. mansoni infection prevented most of the changes occurred during schistosomiasis. Taking in consideration the difference in dose, duration and starting time of treatment used in the present study, our results are promising compared to previous discussed results.
Davis et al. (1974) demonstrated that eggs in liver, elicit T-cell dependent responses leading to macrophage activation and granuloma formation around them. In the meanwhile, patients with chronic schistosomiasis often fail to resolve hepatic fibrosis after eradication of the worms by anthelmintic treatment (Mahmoud et al., 2002). Moreover, McCormick et al. (1996) demonstrated that there is an important role of anti-oxidant processes and oxidative stress in mediating liver injury in schistosomiasis due to an increased production of reactive oxygen intermediates by eosinophils and macrophages at the site of granulomatous inflammation. Hence, the suppressive effect of both N. sativa oil or melatonin on granuloma formation and the improvement in the haematological parameters and histopathological state of both liver and kidney is probably due, in part, to the fact that these antioxidants were administered early before granulomas formation, when immune competent cells were not yet stimulated or due to a modulation in the immune response.
REFERENCES
- Cheever, A.W., M.E. Williams, T.A. Wynn, F.D. Finkelman and R.A. Seder et al., 1994. Anti-IL-4 treatment of Schistosoma mansoni-infected mice inhibits development of T cells and non-B, non-T cells expressing Th2 cytokines while decreasing egg-induced hepatic fibrosis. J. Immunol., 153: 753-759.
PubMed - El-Dakhakhny, M., N.I. Mady and M.A. Halim, 2000. Nigella sativa L. oil protects against induced hepatotoxicity and improves serum lipid profile in rats. Arzneimittelforschung, 50: 832-836.
CrossRefPubMedDirect Link - El-Sokkary, G.H., H.M. Omar, A.T. Hassanein, S. Cuzzocrea and R.J. Reiter, 2002. Melatonin reduces oxidative damage and increases survival of mice infected with S. mansoni. Free Rad. Biol. Med., 32: 319-332.
PubMedDirect Link - Hotchkiss, R.D., 1948. A microchemical reaction resulting in the staining of polysaccharide structures in fixed tissue preparations. Arch. Biochem., 16: 131-141.
PubMedDirect Link - Mahmoud, M.R., H.S. El-Abhar and S. Saleh, 2002. The effect of Nigella sativa oil against the liver damage induced by Schistosoma mansoni infection in mice. J. Ethnopharmacol., 79: 1-11.
CrossRefDirect Link - McCormick, M.L., A. Metwalli, M.A. Railsback, J.V. Weinstock and B.E. Britigan, 1996. Eosinophils from schistosome-induced hepatic granulomas produce superoxide and hydroxyl radical. J. Immunol., 157: 5009-5015.
PubMedDirect Link - Tesoriere, L., M. D. Allegra, D. D`Arpa, D. Butera and M. A. Livrea, 2001. Reaction of melatonin with hemoglobin-derived oxoferryl radicals and inhibition of the hydroperoxide-induced hemoglobin denaturation in red blood cells. J. Pineal. Res., 31: 114-119.
PubMed - Zaoui, A., Y. Cherrah, N. Mahassini, K. Alaoui, H. Amarouch and M. Hassar, 2002. Acute and chronic toxicity of Nigella sativa fixed oil. Phytomedicine, 9: 69-74.
CrossRefPubMedDirect Link