
A.T. Owolabi
Department of Botany, University of Calabar, PMB 1115, Calabar, Nigeria
E.E. Ekpiken
Department of Botany, University of Calabar, PMB 1115, Calabar, Nigeria
International Journal of Virology
Year: 2014 | Volume: 10 | Issue: 4 | Page No.: 253-262







ABSTRACT
Aphis spiraecola (Patch), a ubiquitous aphid in West Africa has been implicated in the transmission of most potyviruses reported from Nigeria. The transmission efficiency of two Nigerian strains of MWMV designated as MWMV-lag and MWMV-cor in this study, by two clones of the aphid was studied. The transmission efficiency was determined based on aphid number per plant, varying Acquisition Access Periods (AAPs) and varying post-acquisition starvation times. Both clones of A. spiraecola were found to be efficient vectors of the two MWMV strains causing 100% transmission rate with as few as 5 individual aphids. The result of this study indicates that transmission efficiencies of MWMV-cor and MWMV-lag strains by the Aphis spiraecola clones declined with prolongation of AAPs. Clone I was more efficient in transmitting MWMV-cor than MWMV-lag. For clone II, both virus strains were transmitted with equal efficiency differing by 6.7% where differences occurred. Both clones transmitted MWMV-lag more efficiently that it did MWMV-cor with respect to the sequestration time regimes employed. Percentage differences ranged from 6.6-40% for MWMV-cor and 11.3-30% for MWMV-lag. On clonal basis, clone I did better in retaining and transmitting MWMV-cor that did clone II while MWMV-lag was retained much longer than MWMV-cor by clone II.
PDF Abstract XML References Citation
Received: June 10, 2014;
Accepted: September 10, 2014;
Published: December 01, 2014
How to cite this article
A.T. Owolabi and E.E. Ekpiken, 2014. Transmission Efficiency of Two Strains of Moroccan Watermelon Mosaic Virus
by Two Clones of Aphis spiraecola (Patch). International Journal of Virology, 10: 253-262.
DOI: 10.3923/ijv.2014.253.262
URL: https://scialert.net/abstract/?doi=ijv.2014.253.262
DOI: 10.3923/ijv.2014.253.262
URL: https://scialert.net/abstract/?doi=ijv.2014.253.262
INTRODUCTION
Aphis spiraecola Patch commonly called green citrus or Siam weed aphid is pandemic in distribution and has been reported to be a pest of many economically important crops and ornamentals spanning 90 plant families including Cucurbitaceae, Convolvulaceae Fabaceae, Rosaceae and Solanaceae (Blackman and Eastop, 1994). In Nigeria, the natural host is the Siam weed, Chromolaena odorata (L.), a ubiquitous weed that has become a menace in the country.
The aphid has been reported to vector several economically important viruses including Papaya Ring Spot Virus (PRSV) (Rivas Platero and Larios, 1994), Watermelon Mosaic Virus-2 (WMV-2) (Webb et al., 1994), Plum Pox Virus (PPV), (Gildow et al., 2004), Cucumber Mosaic Virus (CMV) (Gildow et al., 2008), Zucchini Yellow Mosaic Virus (ZYMV) (Katis et al., 2006), Potato virus YNTN (Boukhris-Bouhachem et al., 2011), Pepper Veinal Mosaic Virus (PVMV) (Fajinmi et al., 2011) and Citrus Tristeza Virus (CTV) (Vidal et al., 2012).
In Nigeria, the aphid has also been implicated in the transmission of most potyviruses reported. These include Cowpea Aphid-borne Mosaic Virus (CAMV) (Atiri et al., 1986), Telfairia Mosaic Virus (TeMV) (Shoyinka et al., 1987), Celosia mosaic virus (Owolabi et al., 1998), Senna mosaic virus (Owolabi and Proll, 2001), PRSV (Owolabi et al., 2008), Pepper veinal mottle virus (Fajinmi et al., 2011) and Cucurbita mosaic virus (Owolabi et al., 2011).
The efficiency with which plant viruses are transmitted by specific aphid species has been reported to be influenced by the number of individual aphids and aphid biotypes (Berger et al., 1983; Katis et al., 2006; Moreno et al., 2007; Kalleshwaraswamy and Kumar, 2007), aphid species (Garzo et al., 2004; Gildow et al., 2008) the virus strains (Verbeek et al., 2010; Boukhris-Bouhachem et al., 2011; Mello et al., 2011) and aphid clones (Lucio-Zavaleta et al., 2001; Kanavaki et al., 2006; Symmes and Perring, 2007).
Moroccan Watermelon Mosaic Virus (MWMV), a potyvirus which was first reported by Fischer and Lockhart (1974) from southern Europe including Spain, Italy and France (Quiot-Douine et al., 1990; Roggero et al., 1998; Lecoq et al., 2007) and in East, West, Central and Southern Africa (Van der Meer and Garnett, 1987; Lecoq et al., 2001; Arocha et al., 2008; Yakoubi et al., 2008). The virus which is now considered an emerging threat to cucurbit production (Lecoq et al., 2001; Lecoq, 2004) induces severe mosaic and leaf malformation in susceptible cucurbit hosts (Owolabi et al., 2012; Malandraki et al., 2014).
Recently, two strains of MWMV isolated from Lagenaria breviflora Robert and Coccinia barteri (Hook. f.) Keay were reported from Nigeria (Owolabi et al., 2012). The Lagenaria strain was particularly found to be more virulent than the Coccinia strain, causing severe stunting of cucumber and Cucumeropsis mannii Naudin. Both strains were transmitted in a fore-gut (non-persistent) manner by Myzus persicae (Sulzer) and by a few other aphid species. Preliminary aphid transmission tests in our screenhouse have also shown A. spiraecola to be a veritable vector of the two virus strains. Researches on the economic significance of the virus strains in our screen-house (unpublished data) showed that both portent serious danger to cucurbit production. This study was undertaken to investigate the transmission efficiency of the Nigerian strains of MWMV by two clones of the A. spiraecola. The epidemiological significance of the results is discussed.
MATERIALS AND METHODS
Virus strains: The virus strains used in the study were two strains of MWMV isolated from Lagenaria breviflora and Coccinia barteri described by Owolabi et al. (2012) herein designated MWMV-lag and MWMV-cor, respectively. They were propagated and maintained in Cucumeropsis manni in the screenhouse at 25±3°C.
Sourcing and rearing of aphid cultures: The aphid species used was A. spiraecola. One aphid culture was obtained in Calabar herein referred to as clone 1. The second culture was sourced from Idundu, 30 km north of Calabar herein called clone II. Both locations are in different Local Government Areas of Cross River State, Nigeria.
The insects were dislodged from the natural host by breathing heavily on them. They were carefully transferred by using the tip of moistened artist brush to young seedlings of C. mannii. Rearing was performed by period transfer of apterous individuals of the aphid cultures to young seedlings of the plant every other week in insect proof cages.
Influence of aphid number on per plant on transmission efficiency: To determine the effect of aphid number on transmission efficiency of the insect, the aphid clones were obtained, starved for 1 h and subjected to 1 min Acquisition Access Period (AAP) on symptomatic leaves of C. mannii, the source plant. Groups of 1, 3, 5 and 7 viruliferous aphids were then transferred to each of five seedlings of Cucumis sativus L. (the test plant) for 3 min Inoculation Access Period (IAP). The aphids were considered to have fed when they became still on the source plant. The inoculated plants were left in insect proof cages in the screen-house for two weeks for symptom development.
Influence of varying Acquisition Access Periods (AAP) on transmission efficiency: For the determination of varied acquisition access period on transmission efficiency, individual aphids from both clones were allowed AAPs of 1, 3, 5, 7 min time regimes. Seven aphids were allowed to feed but 5 viruliferous individuals were transferred immediately for the transmission tests. For the 1 min AAP, aphids were transferred singly from the source plant to the test plants to ensure time precision while for the other time regimes the aphids were transferred in groups.
Influence of post-acquisition starvation (sequestration time) on transmission efficiency: For this experiment, groups of aphids of both clones were subjected to starvation periods of 1, 10, 15 and 20 min after they had been allowed 1-3 min AAP from the source plant. Five viruliferous aphids were then transferred to each of five test plants.
In all the experiments, the aphids were starved in petri dishes with lids made of gauze (to ensure aeration), tightly held in place at the rims with transparent tapes. Transferring of viruliferous aphids from source plant to the test plants was carried out using moistened paint brush, after a gentle teasing of the insects from their posterior ends to avoid damage to the stylets. Insect-inoculated plants were sprayed with Pirimor 50-DF (2-Dimethylamino-5,6-dimethylpyrimidin-4-yl dimethyl-carbamate) and left in insect-screened cages in the screenhouse with ambient temperature of 25±2°C. Whether test insects were inoculative was determined on the basis of symptom development in indicator assay plants.
Statistical analysis: Each transmission experiment was repeated trice for each virus/vector combination. The number of test plants for each was five (N = 5). Data were collected and analyzed using Statistical Package for Social Science (SPSS) version 14. Means were separated using the Least Significant Difference (LSD).
RESULTS
Effect of aphid number on transmission efficiency of the virus strains by the aphid clones: The results of the transmission of MWMV-cor and MWMV-lag strains by the two aphid biotypes based on aphid number per plant are presented in Table 1. Generally, the two strains were readily transmitted by both clones of A. spiraecola. Also, percentage transmission of both virus strains by the aphid clones increased as the number of aphids employed per plant increased. Five aphids of both clones were enough to cause 100% transmission of the virus strains. There was a 20% increase in the transmission of MWMV-lag compared to that of MWMV-cor when single aphid of clone I was employed per plant. Conversely, the same margin of increase was observed for the transmission of MWMV-cor in comparison to that of MWMV-lag when three aphids were used. Employing 7 aphids per plant produced a marginal 6.7% increase for MWMV-cor over the figure obtained for MWMC-lag.
| Table 1: | Transmission efficiency of MWMV-cor and MWMV-lag strains by two clones of Aphis spiraecola based on aphid number/plant |
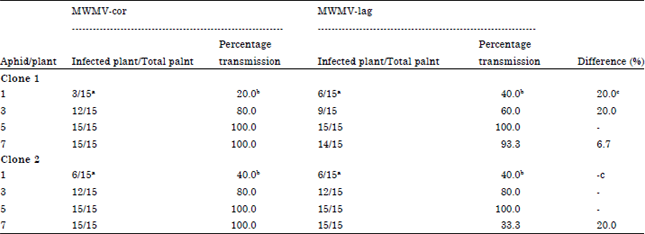 | |
| aNominator is the number of plants infected while the denominator is the total number of plants in 3 trials (5 plants per trial), bPercentage transmission, CDifference in percentage transmission between the two strains | |
| Table 2: | Influence of aphid number on the transmission efficiency of Coccinia and Lagenaria strains of Morocco watermelon mosaic virus by two clones of Aphis spiraecola |
 | |
| Means (obtained by dividing the number of plants infected/the number of trials) followed by the same letter in each column are not significantly different (p = 0.05) using Least Significant Difference (LSD), AL 1: One aphid/plant, AL 3: Three aphids/plant, AL 5: Five aphids/plant, AL 7: Seven aphids/plant | |
For clone II, both strains of virus were transmitted with equal efficiency by the clones when 1, 3 and 5 aphids were employed per plant. However, MWMV-cor was transmitted more efficiently with a 20% increase over the figure obtained for MWMV-lag when 7 aphids were used.
Comparing the transmission of MWMV-cor by both clones, the virus strain was transmitted with equal efficiency by the two clones of the aphid species (columns 3 and 8), except when 1 aphid was employed for the transmission test when clone II was found to be twice efficient in transmitting the virus strain with 40% infection compared to clone I which produced 20% infection.
With regard to the transmission of MWMV-lag by the two clones, clone II caused higher percentage infection (80%) using 3 aphids compared to 60% for clone I. The strain was transmitted with equal efficiency when 1 and 5 aphids were employed (columns 5 and 10). Statistically, the transmission efficiency of MWMV-cor by clone II was only significantly higher (p<0.05) with single aphid inoculation compared to the value for clone I (Table 2). For the MWMV-lag strain, using three and seven aphids led to significantly higher transmission efficiency than recorded for clone I.
Transmission of the virus strains by the aphid clones based on AAPs: The result of the transmission efficiency of MWMV-cor and MWMV-lag strains by the two aphids clones of A. spiraecola based on AAPs are presented in Table 3. Generally, percentage transmission of both MWMV strains by the two aphid clones decreased as AAPs increased.
| Table 3: | Transmission efficiency of MWMV-cor and MWMV-lag by two clones of Aphis spiraecola based on acquisition access period/inoculation access period |
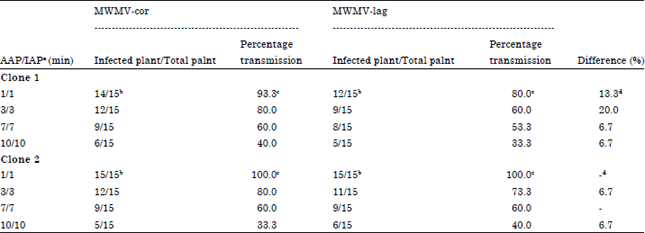 | |
| aAAP/IAP: Acquisition access period/inoculation access period, bNominator is the number of plants infected while the denominator is the total number of plants in 3 trials (5 plants per trial), cPercentage transmission, dDifference in percentage transmission between the two strains | |
Comparing the transmission of MWMV-lag and MWMV-cor by clone I, the clone was more efficient in transmitting the latter than the former. Percentage transmission figures were higher for all the time regimes employed (columns 3 and 5). While percentage transmission values of 93.3, 80, 60 and 40 were obtained for MWMV-cor, the values recorded for MWMV-lag were 80, 60, 53.3 and 33.3 at 1, 3, 7 and 10 AAPs, respectively.
There was no definite trend observed comparing the transmission of MWMV-lag with MWMV-cor by clone II. Transmission was only higher by 6.7% at 10 APP but lower than that of MWMV-cor by the same margin at 3/3 AAP (columns 8, 10).
Comparing the transmission of MWMV-cor by both clones (columns 3 and 8), differences in transmission efficiency were only observed between both strains when allowed 1 and 10 AAPs with a marginal 6.7% difference in both cases. While clone II achieved 100% transmission of the virus strain when given 1 AAP, the percentage transmission value for clone I was 93.3%. On the other hand, when given 10 AAP clone I achieved 40% transmission efficiency compared to 33.3% for clone II.
Clone II transmitted the MWMV-lag strain more efficiently than observed for clone I at all the AAP time regimes employed. While the percentage transmission by clone II was 100% at 1, the value obtained for clone I was 80%. At 3, 7 and 10 APPs, percentage transmission values for clone II were 73.3, 60 and 40%, respectively compared to 60, 53.3 and 33.3% for clone I (columns 5 and 10). Percentage differences ranged from 6.7-20 (column 6).
The values obtained for both clones with respect to MWMV-cor transmission were statistically insignificantly (p<0.05) irrespective of the AAPs. For MWMV-lag transmission, clone II only engendered statistically significant higher value at AIT 1 than clone I (Table 4).
Effect of post-acquisition on transmission of the virus strains by the aphid clones: The results of the transmission of MWMV-lag and MWMV-cor based on retention capability after virus acquisition and transmission (post acquisition starvation or sequestration period) by the two clones of A. spiraecola are presented in Table 5. Generally, percentage transmission of both strains decreased as post-acquisition time increased for both clones.
| Table 4: | Influence of acquisition access period/inoculation access periods on the transmission efficiency of Coccinia and Lagenaria strains of Morocco watermelon mosaic virus by two clones of Aphis spiraecola |
 | |
| Means (obtained by dividing the number of plants infected/the number of trials) followed by the same letter in each column are not significantly different (p = 0.05) using Least Significant Difference (LSD). AIT 1: 1 min acquisition/inoculation access time, AIT 3: 3 min acquisition/inoculation access time, AIT 7: 7 min acquisition/inoculation access time, AIT 10 : 10 min acquisition/inoculation access time | |
| Table 5: | Retention capability of the MWMV-cor and MWMV-lag by two clones of Aphis spiraecola |
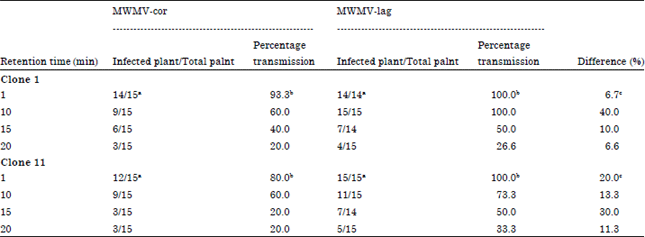 | |
| aNominator is the number of plants infected while the denominator is the total number of plants in 3 trials (5 plants per trial), bPercentage transmission, CDifference in percentage transmission between the two strains | |
Comparing the transmission of MWMV-cor and MWMV-lag by clone I, the result showed that MWMV-lag was transmitted more efficiently than MWMV-cor as percentage transmission values were higher at all the time regimes used (columns 3 and 5). The percentage transmission values were 100% after 1 and 10 min, 50 and 26.6% after 15 and 20 min sequestration periods for MWMV-lag while the corresponding values for MWMV-cor were 93.9, 60, 40 and 20%.
Clone II appeared to be more efficient in retaining MWMV-lag and consequently produced higher percentage transmission values than MWMV-cor. Percentage transmission values were also higher for the former than the latter at all the time regimes employed (columns 8 and 10).
Comparing the transmission of MWMV-cor by both clones, the data showed that clone I appeared more efficient in retaining the virus strain than clone II for shorter sequestration times of between 1-15 min. But further increase in the sequestration time produced similar effect (columns 3 and 8).
For MWMV-lag, except at 20 min post acquisition starvation time when clone I produced 26.6% percentage transmission compared to 33.3% for clone II, the former appeared more efficient than the latter in retaining and consequently transmitting the virus strain (columns 5 and 10). Transmission efficiency of MWMV-cor by clone I was significantly higher (p<0.05) at ST I and ST 15 in comparison to the values obtained for clone II (Table 6). For MWMV-lag transmission efficiency was only significantly higher at ST 10 compared to that of clone II.
| Table 6: | Influence of post-acquisition starvation time on the transmission efficiency of Coccinia and Lagenaria strains of Morocco watermelon mosaic virus by two clones of Aphis spiraecola |
 | |
| Means (obtained by dividing the number of plants infected/the number of trials) followed by the same letter in each column are not significantly different (p = 0.05) using Least Significant Difference (LSD), ST 1: 1 min post-acquisition starvation time, ST 10: 10 min post-acquisition starvation time, ST 15: 15 min post-acquisition starvation time, ST 20: 20 min post-acquisition starvation time | |
DISCUSSION
A. spiraecola is ubiquitous in Nigeria and has been implicated as a vector of most potyviruses reported from the country. Moroccan watermelon mosaic virus on the other hand, is regarded as an emerging threat to cucurbit production wherever it occurs. In this study, the transmission efficiency of two strains of MWMV virus by two clones of A. spiraecola was investigated. Transmission efficiency was determined based on aphid number per plant, varying acquisition access periods/inoculation access periods and ability of the clones to retain and transmit the viruses after varying post-acquisition starvation times.
The results of this study showed a linear relationship between aphid number per plant and the number of infected plants as percentage transmission for both virus strains by the aphid clones increased as the number of aphids employed per plant increased. For both virus strains, five aphids were enough to cause 100% transmission by the two clones. This result lends credence to earlier reports by Katis et al. (2006) and Kalleshwaraswamy and Kumar (2007) when they showed that multiple aphid inoculations led to higher transmission efficiencies of ZYMV and PRSV by their aphid vectors than single aphid inoculations.
Aphid transmission of fore-gut borne (non-persistent or stylet-borne) viruses, according to Hull (2002) is characterized by short acquisition and inoculation access feeding times often a matter of seconds or minutes while long acquisition and retention times reduced efficiency of transmission. The result of this study indicates that transmission efficiencies of MWMV-cor and MWMV-lag strains by the two A. spiraecola clones declined with prolongation of AAPs. However, clone I consistently demonstrated greater efficiency in transmitting MWMV-cor than it did the MWMV-lag. No such consistency was observed for clone II in the transmission of MWMV-cor compared to MWMV-lag. Singh and Singh (2010) reported a decline in the transmission efficiency of PRSV by M. persicae while increasing the acquisition feeding and starvation times. Kostiw and Trojanowska (2011) have also shown that continuous prolongation of the feeding time caused a slow decrease in the effectiveness of transmission of PVYNTN and PVYN by M. persicae.
Regarding post-acquisition periods and transmission efficiencies of the virus strains, both clones showed greater efficiencies in transmitting MWMV-lag than they did MWMV-cor. With longer post-acquisition time employed in this study, clone II was more efficient than clone I in transmitting MWMV-lag while there was parity in the transmission of MWMV-cor by the two clones. Aphid clones have been shown to differ in the efficiency with which they transmit fore-gut viruses depending on the virus strain-vector combinations. Garzo et al. (2004) found no significant difference between clones of Aphid fabae, Aphid gossypii, Myzus persicae and A craccivora in the transmission of different isolates of CMV and ZYMV. Symmes and Perring (2007) also found no significant difference in the transmission efficiencies of ZYMV by two clones of M. persicae obtained and reared on brassicaceous plants. On the other hand, Kanavaki et al. (2006) found that M. persicae nicotianae was more efficient than M. persicae in transmitting PVYN by 12 clonal lineages of both aphids. In their study, Berger et al. (1983) also demonstrated that clones of biotype E was generally more efficient in transmitting two strains of MDMV (MDMV-A and MDMV-B) than biotype C. Verbeek et al. (2010) reported comparable transmission efficiencies for PVYN, PVYNTN and PVYN-Wi by biotype Mp2 of M. persicae. Mello et al. (2011) have also reported no apparent differences in the ability of a clone of M. persicae in transmitting five isolates of PVYO and PVYNW-i.
CONCLUSION
From the result of this study, both clones of A. spiraecola could be said to be efficient vectors of MWMV strains causing 100% transmission rate with as few as 5 individual aphids. Both clones transmitted MWMV-lag, the more virulent of the two strains, more efficiently than MWMV-cor and Clone II appeared to be more efficient than clone I in transmitting both virus strains in several of the tests. The aphid clones could also retain the virus strains for as long as 20 min post acquisition. The implication of both clones retaining the virus strains for this length of time is the prospect of epiphytotics and possible crop losses should the A. spiraecola clones carrying MWMV strains be brought into susceptible cucurbit crops in the fields.
ACKNOWLEDGMENT
The authors greatly acknowledge the staff of the Botanical Garden, University of Calabar, Calabar for helping in raring the aphid cultures.
REFERENCES
- Arocha, Y., N. Vigheri, B. Nkoy-Florent, K. Bakwanamaha and B. Bolomphety et al., 2008. First report of the identification of Moroccan watermelon mosaic virus in papaya in Democratic Republic of Congo. Plant Pathol., 57: 387-387.
CrossRef - Atiri, G.I., D.A. Enobakhare and G. Thottappilly, 1986. The importance of colonizing and non-colonizing aphid vectors in the spread of cowpea aphid-borne mosaic virus in cowpea. Crop Protect., 5: 406-410.
CrossRefDirect Link - Berger, P.H., R.W. Toler and K.F. Harris, 1983. Maize dwarf mosaic virus transmission by greenbug biotypes. Plant Dis., 67: 496-497.
CrossRef - Boukhris-Bouhachem, S., J. Rouze-Jouan, R. Souissi, L. Glais and M. Hulle, 2011. Transmission efficiency of the strain PVYNTN by commonly captured aphids in tunisian potato fields. Plant Pathol. J., 10: 22-28.
CrossRefDirect Link - Fajinmi, A.A., C.A. Odebode and O.B. Fajinmi, 2011. The effect of agro-ecological zones on the incidence and distribution of aphid vectors of pepper veinal mottle virus, on cultivated pepper (Capsicum annuum L.) in Nigeria. J. Central Eur. Agric., 12: 528-542.
Direct Link - Fischer, H.U. and B.E.L. Lockhart, 1974. Serious losses in cucurbits caused by watermelon mosaic virus in Morocco. Plant Dis. Rep., 58: 143-146.
Direct Link - Garzo, E.I., M. Duque and A. Fereres, 2004. Transmission efficiency of different non-persistent viruses infecting melon by four aphid species. Spanish J. Agric. Res., 2: 369-376.
Direct Link - Gildow, F.E., D.A. Shah, W.M. Sackett, T. Butzler, B.A. Nault and S.J. Fleischer, 2008. Transmission efficiency of cucumber mosaic virus by aphids associated with virus epidemics in snap bean. Phytopathology, 98: 1233-1241.
Direct Link - Kalleshwaraswamy, C.M. and N.K.K. Kumar, 2007. Transmission efficiency of Papaya ringspot virus by three aphid species. Phytopathology, 98: 541-546.
CrossRefDirect Link - Katis, N.I., J.A. Tsitsipis, D.P. Lykouressis, A. Papapanayotou and J.T. Margaritopoulos et al., 2006. Transmission of Zucchini yellow mosaic virus by colonizing and non-colonizing aphids in Greece and new aphid species vectors of the virus. J. Phytopathol., 154: 293-302.
CrossRefDirect Link - Kanavaki, O.M., J.T. Margaritopoulos, N.I. Katis, P. Skouras and J.A. Tsitsipis, 2006. Transmission of potato virus Y in tobacco plants by Myzus persicae nicotianae and M. persicae s.str. Plant Dis., 90: 777-782.
CrossRefDirect Link - Kostiw, M. and E. Trojanowska, 2011. Impact of feeding time on PVYN and PVYNTN transmission by Myzus persicae (Sulz.). J. Plant Protect. Res., 51: 429-434.
CrossRef - Lecoq, H., G. Dafalla, C. Desbiez, C. Wipf-Scheibel and B. Delecolle et al., 2001. Biological and molecular characterization of Moroccan watermelon mosaic virus and a potyvirus isolate from eastern Sudan. Plant Dis., 85: 547-552.
CrossRefDirect Link - Lecoq, H., I. Justafre, C. Wipf-Scheibel and C. Desbiez, 2007. Moroccan watermelon mosaic virus newly reported on zucchini squash in France. New Dis. Rep., 16: 19-19.
Direct Link - Gildow, F.E., L. Levy, V.D. Damsteegt, A.L. Stone, W.L. Schneider and D.G. Luster, 2004. Transmission of three North American isolates of Plum pox virus: Identification of aphid vectors and species-specific transmission from infected stone fruits. Acta Hort., 657: 207-211.
Direct Link - Lucio-Zavaleta, E., D.M. Smith and S.M. Gray, 2001. Variation in transmission efficiency among Barley yellow dwarf virus-RMV isolates and clones of the normally inefficient aphid vector, Rhopalosiphum padi. Phytopathology, 91: 792-796.
CrossRefDirect Link - Malandraki, I., N. Vassilakos, C. Xanthis, G. Kontosfiris, N.I. Katis and C. Varveri, 2014. First report of Moroccan watermelon mosaic virus in zucchini crops in Greece. Plant Dis., 98: 702-702.
CrossRefDirect Link - Mello, A.F.S., R.A. Olarte, S.M. Gray and K.L. Perry, 2011. Transmission efficiency of Potato virus Y strains PVYO and PVYN-Wi by five aphid species. Plant Dis., 95: 1279-1283.
CrossRefDirect Link - Moreno, A., E. Bertolini, A. Olmos, M. Cambra and A. Fereres, 2007. Estimation of vector propensity for Lettuce mosaic virus based on viral detection in single aphids. Spanish J. Agric. Res., 5: 376-384.
Direct Link - Owolabi, A.T., M.A. Taiwo, G.A. Thottappilly, S.A. Shoyinka, E. Proll and F. Rabenstein, 1998. Properties of a virus causing mosaic and leaf curl disease of Celosia argentea L. in Nigeria. Acta Virol., 42: 133-139.
PubMed - Owolabi, A.T. and E. Proll, 2001. A mosaic disease of Senna hirsuta induced by a potyvirus in Nigeria. Acta Virol., 45: 73-79.
PubMed - Owolabi, A.T., F. Rabenstein, M.A. Taiwo, F. Ehrigh and E. Maiss, 2011. Characterization and Identification of a potyvirus causing mosaic disease of Cucurbita moschata Duch Ex. Poir in Calabar, South East Nigeria. Int. J. Plant Pathol., 2: 165-176.
CrossRefDirect Link - Owolabi, A.T., F. Rabenstein, F. Ehrig, M. Maiss Edgar and H.J. Vetten, 2012. Strains of Moroccan watermelon mosaic virus isolated from Lagenaria breviflorus and Coccinia barteri in calabar, Southeastern Nigeria. Int. J. Virol., 8: 258-270.
CrossRefDirect Link - Quiot-Douine, L., H. Lecoq, J.B. Quiot, M. Pitrat and G. Labonne, 1990. Serological and biological variability of virus isolates related to strains of papaya ringspot virus. Phytopathology, 80: 256-263.
CrossRefDirect Link - Rivas Platero, G.G. and J.F. Larios, 1994. Epidemiology of Papaya ringspot virus (PRSV) in Zapotitan, El Salvador. Manejo Integrado Plagas, 32: 5-7.
Direct Link - Roggero, P., G. Dellavalle, V. Lisa and V.M. Stravato, 1998. First report of Moroccan watermelon mosaic potyvirus in Zucchini in Italy. Plant Dis., 82: 351-351.
CrossRefDirect Link - Singh, V. and D. Singh, 2010. Studies on natural transmission of papaya ringspot virus disease in Eastern Uttar Pradesh. Ann. Plant Prot. Sci., 18: 188-192.
Direct Link - Symmes, E.J. and T.M. Perring, 2007. Intraspecific variation in zucchini yellow mosaic virus transmission by Myzus persicae and the impact of aphid host plant. J. Econ. Entomol., 100: 1764-1772.
CrossRefDirect Link - Van der Meer, F.W. and H.M. Garnett, 1987. Purification and identification of a South African isolate of Watermelon mosaic virus-Moroccan. J. Phytopathol., 120: 255-270.
CrossRefDirect Link - Vidal, E., A. Moreno, E. Bertolini, M.C. Martinez, A.R. Corrales and M. Cambra, 2012. Epidemiology of Citrus tristeza virus in nursery blocks of Citrus macrophylla and evaluation of control measures. Span. J. Agric. Res., 10: 1107-1116.
Direct Link - Webb, S.E., M.L. Kok-Yokomi and D.J. Voegtlin, 1994. Effect of trap color on species composition of alate aphids (Homoptera: Aphididae) caught over watermelon plants. Florida Entomol., 77: 146-154.
Direct Link - Verbeek, M., P.G.M. Piron, A.M. Dullemans, C. Cuperus and R.A.A. van der Vlugt, 2010. Determination of aphid transmission efficiencies for N, NTN and Wilga strains of Potato virus Y. Ann. Applied Biol., 156: 39-49.
CrossRefDirect Link - Yakoubi, S., C. Desbiez, H. Fakhfakh, C. Wipf-Scheibel, M. Marrakchi and H. Lecoq, 2008. Biological characterization and complete nucleotide sequence of a Tunisian isolate of Moroccan watermelon mosaic virus. Arch. Virol., 153: 117-125.
CrossRefPubMedDirect Link