
H.A. Hussein
Department of Virology, Faculty of Veterinary Medicine,Cairo University, Giza 12211 Egypt
H.A. Sultan
Department of Diseases of Poultry, Faculty of Veterinary Medicine, Menoufia University, Egypt
A.H. EL-Deeb
Department of Virology, Faculty of Veterinary Medicine,Cairo University, Giza 12211 Egypt
A.A. El-Sanousi
Department of Virology, Faculty of Veterinary Medicine,Cairo University, Giza 12211 Egypt
International Journal of Virology
Year: 2009 | Volume: 5 | Issue: 1 | Page No.: 36-43







ABSTRACT
Re-emerging of H5N1 severe outbreaks in vaccinated chickens housed in poultry farms in Sharkia governorate in Egypt was observed starting from October 2007 and continued for 3 months until control measures have been taken in such farms, besides multiple vaccination during this period were applied using reassortant H5N1 and 6 types of H5N2 vaccines. In the present study, 9922 serum samples were collected from vaccinated chickens including some broiler breeders and commercial layers from the period of December 2006 to February 2008 and tested for the immune response to the multiple vaccination with different H5N1 and H5N2 avian influenza vaccines (reassortant H5N1 and 6 types of H5N2 vaccines) by Hemagglutination Inhibition (HI) test using heterologous H5 antigen which represent non of the vaccine antigens. The samples were collected from 578 houses mostly commercial layers. H5N1 viral nucleic acid was also detected in swab samples collected from mortalities occurred in some of vaccinated birds after vaccination policy has been applied in Egypt at March 2006. The viral RNAs of the detected H5N1 circulating viruses between Feb. 2006 and Feb. 2008 in Sharkia governorate were sequenced. Results of nucleotide sequence analysis of 11 detected viruses confirm the existence and circulation of the H5N1 in Sharkia governorate along the period of study and no H7 viruses were detected. The serum samples collected from different farms were divided into 4 groups based on the date of collection. Hemagglutination Inhibition (HI )results showed that in the first group (samples collected between December 2006 to March 2007) 43% of the tested samples were of HI titers of 6 log 2 or more which we propose such titer as a protective titer against H5N1 virus. In group 2 (samples collected between April and July, 2007), titers in 36.4% of the tested samples were protective. In group 3 (samples collected between August to November, 2007), titers in 33.2% of the samples were protective. The last group contains samples collected from chicken vaccinated twice during the period of collection (December 2007 to February 2008), besides being previously vaccinated with at least 3 vaccine shots. Hemagglutination Inhibition (HI) titers in such group were protective in 54% of the tested samples although these chickens received 5 doses of H5 vaccines. The present study is highlight the possible multiple causes of re-emergence of H5N1 outbreaks in Egypt.
PDF Abstract XML References Citation
How to cite this article
H.A. Hussein, H.A. Sultan, A.H. EL-Deeb and A.A. El-Sanousi, 2009. Possible Causes of Re-Emerging outbreaks of H5N1 Avian Influenza Virus in Vaccinated Chickens in Sharkia Governorate in Egypt. International Journal of Virology, 5: 36-43.
DOI: 10.3923/ijv.2009.36.43
URL: https://scialert.net/abstract/?doi=ijv.2009.36.43
DOI: 10.3923/ijv.2009.36.43
URL: https://scialert.net/abstract/?doi=ijv.2009.36.43
INTRODUCTION
The number of outbreaks of the Highly Pathogenic Avian Influenza (HPAI) H5N1 disease has increased alarmingly in the last 10 years with unprecedented emergence and spread of the virus in Asia, Europe and Africa. The zoonotic infections of humans have resulted in AI being considered one of the most important animal diseases (Capua and Alexander, 2006). Vaccinology for AI has not grown at the same rate as other infectious diseases of animals. Data are being generated from experimental and field research in AI vaccinology, but the success of AI vaccines under field condition where complex task of vaccinating poultry in different farming and ecologic environments still has area of uncertainty (Capua and Marangon, 2006). Vaccination can be a powerful tool to support eradication programs in increasing resistance to field challenge, reduce shedding levels in vaccinating birds and reduce transmission (Van-Der-Goot et al., 2005). Timely information is needed about the efficacy of vaccination using different approaches. In the last year, one of the possible causes of re-emerging of H5N1 breaks is the failure of the AI vaccines of controlling the outbreaks of H5N1 in Egypt. Moreover, escape mutant strain of H5N1 has been recently reported among poultry farms (Taha et al., 2008).
Testing the efficacy of different H5 AI vaccines as a possible cause of vaccination failure in the 2007 H5N1 breaks in Sharkia governorate is an urgent need.
In the present study, analysis of the immune response to the multiple vaccination with different H5N1 and H5N2 avian influenza vaccines (reassortant H5N1 and 6 types of H5N2 vaccines) by Hemagglutination Inhibition (HI) test using heterologous H5 antigen which represents non of the vaccine antigens has been carried out. Also, RT-PCR to screen H5 and H7 AI viruses and sequence analysis of the circulating viruses detected in vaccinated flocks with mortalities in the period between 2006 to 2008 have been applied.
MATERIALS AND METHODS
Serum Samples
Blood samples were collected from wing vein and kept in a slope position at 37°C for 30 min then at 4°C overnight. Sera were then separated by centrifugation at 3000 rpm/ 20 min and stored at -20°C till used. All sera were inactivated at 56°C for 30 min before used.
HI Test
The test was performed according to OIE Manual (2005).
RT-PCR
RNA was extracted from the tracheal swabs collected from dead birds in 11 vaccinated flocks along the period of study. Primers specific for type A influenza (M gene) and H5 and H7 of AIV were designed and RT-PCR protocol was used according to Hussein, (2001). The sequences of the designed primers were as follows:
| • | For detection of any type A influenza viruses (M gene primers MF and MR were used) MF: 5’ AGA TGA GTC TTC TAA CCG AGG TCG 3’ MR: 5’ TGC AAA AAC ATC TTC AAG TCT CTG 3’ |
| • | For typing of the detected viruses (H5 and H7 specific primers were used) H5 Forward: 5’ ACG TAT GAC TAT CCA CAA TAC TCA G 3’ H5 Reverse: 5’ AGA CCA GCT ACC ATG ATT GC3’ H7 Forward: 5’ ATT GGA CAC GAG ACG CAA TG 3’ H7 Reverse: 5’ TTC TGA GTC CGC AAG ATC TAT TG 3’ |
Sequence Analysis
PCR products of the detected viruses were sequenced in the gene sequencing unit at VACSERA.
RESULTS AND DISCUSSION
The zoonotic infections of humans have resulted in AI being considered one of the most important animal diseases (Capua and Alexander, 2006). The development of H5N1 vaccine is recognized as the primary strategy to protect human against a possible H5N1 pandemic (Suguiten et al., 2006). Infections with H5N1 AIV in birds and in humans have been occurring since 1997 and the phylogenetic and antigenic analysis of H5N1 viruses collected over the last period indicated that they have evolved into different sublineages or clades: 2004 viruses are designated clade 1, 2003 viruses clade 1’, some 2005 viruses clade 2 and 1997 viruses clade 3 (WHO 2005, Chen et al., 2006).
H5N1 virus is an influenza A virus first detected in china from geese in Guangdong Province at 1996 (Chen et al., 2004). In 1997, H5N1 AIV caused disease outbreak in poultry in Hong Kong and was transmitted to human causing 6 deaths (Shortridge et al., 1998). Starting from late 2003, H5N1 viruses began to spread and cause disease outbreaks in China (Wan et al., 2005), Japan (Mase et al., 2005), South Korea (Lee et al., 2005), Thailand, Vietnam, Indonesia, Malaysia, Cambodia and Laos (www.oie.int) resulting in the destruction of hundreds of millions of poultry including chicken, ducks and geese. In May 2005 an H5N1 HPAI virus outbreak occurred in wild birds in Qinghai lake in Western China (Chen et al., 2005, 2006) one of the genotypes of H5N1 virus which identified from the wild birds population continued to spread (goose) to countries in Europe, Africa, the Middle East and Middle Asia (www.oie.int) and caused disease and deaths in wild birds and domestic poultry (Ge et al., 2007). Recently over two hundred human cases have been confirmed around the world confirming the public health importance to effectively control avian influenza. Most of HPAI viruses arise by mutation after LPAI viruses have been introduced into poultry, thus mutations caused by several mechanisms leads to spontaneous duplication of purine triplet which resulted in the insertion of a basic amino acid at the HA0 cleavage site and these occurred due to transcription fault by polymerase complex (Perdue et al., 1997). The factors that bring about mutation from LPAI to HPAI are not known, however the wider the circulation the higher mutation will occur. H5N1 genomes characterized by continuous genetic diversity leading to at least four distinct clades which have been identified with gradual genetic changes in the subclades (Table 1).
Timely information is needed about the efficacy of vaccination using different approaches. Specially, in the last two years present field observation suggested that the failure of the AI vaccines to control the outbreaks of H5N1 in Egypt may be one of the possible causes of re-emergence of H5N1 breaks (Table 2). Moreover, escape mutant strain of H5N1 has been recently reported among poultry farms (Taha et al., 2008).
The present study presents a preliminary field evaluation of the efficacy of the commercially available imported vaccines (H5N1 and H5N2) in Sharkia governorate. When 6 log 2 HI titer using heterologous H5 antigen proposed to be the protective titer against mortalities caused by H5N1 infections, the results indicated that between 45.9-66.7% of the vaccinated chicken were considered unprotective (Table 2).
| Table 1: | Mean HI titers in vaccinated chicken flocks with different avian influenza vaccines (H5N1 and H5N2) |
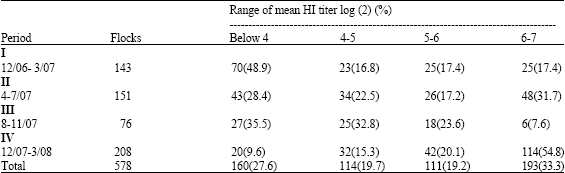 | |
| Table 2: | The number and percentage of the samples of vaccinated chickens showing protective and non protective HI titers for AIV |
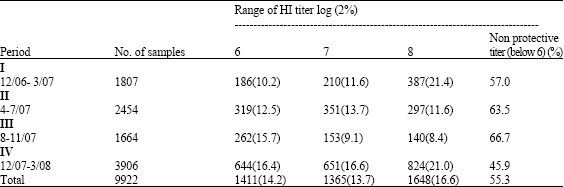 | |
| Table 3: | Field efficacy of the H5 AI vaccines in vaccinated chicken flocks in Sharkia governorate |
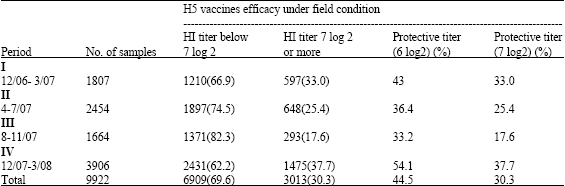 | |
However, the 7 log 2 HI titer which we propose in the study may indicate the real efficacy and immunogenicity of the used vaccines with overall percentage of 30.3 along the two years period of the present study. Indeed, the maximum protection percentage obtained in the field was 37.7% where multiple vaccination at least 4 times were applied (Table 3). Age of birds and vaccine dose were previously reported by others to play an important role in the efficacy of the immune response (Stone, 1987; Swayne et al., 1999). Also, there is a direct relation between the immunogenicity of the inactivated AIV vaccine and the incorporated antigenic mass content and its formulation in the final preparation (Trani et al., 2003). Thus, it is clear that there are many factors influence the success of the vaccination in the field. Although, vaccine application, vaccine dose, route of administration may considered the most important factors, the use of commercial AI vaccines under field condition in Egypt was unsatisfactory and may be one of the main reasons accelerating the evolution of antigenic variants (Sultan and Hussein, 2008). In a recent study, the genetic evolution under vaccination pressure has been reported by Escorcia et al. (2008). Moreover, the genetic variation was due to the effect of long term use of vaccine in H5N2 AIV in Mexico has been reported by Lee et al. (2004). The antigenic drifts occurring in the presence of vaccine pressure resulted in the total failure of the vaccine to prevent the virus shedding when vaccinated birds were challenged with antigenically different strains even within the same lineage (Lee et al., 2005). The current increase in the incidence of AIV infection seen in the field may be related to antigenic drifts which are most likely accumulated along the last two years especially after the extensive use of the vaccines in the field. Like in Mexico, the situation under field condition is similar. It is expected to see more respiratory signs of disease in vaccinated and challenged birds. The possible acceleration in accumulation of mutations in HA gene resulting in the antigenic drifts may be attributed to the use of vaccination (Escorcia et al., 2008). The efficacy of the direct control measures implemented to support the use of commercial vaccine and the vaccine quality may represent the main causes of the vaccination failure under field condition in Egypt. Surveillance programme is a principle aim in the control strategy of HPAI (Capua and Marangon, 2003). Sequence analysis of the recent isolated AIV strains from vaccinated flocks in Egypt showed the occurrence of mutations in critical sites. However, the effect of such mutations on the efficacy of vaccines in field still to be uncertain. The homology percentage between the isolated AI field strains and the used vaccine virus is essential to reduce the spread levels of the AIV in the field (Swayne et al., 2000). In Egypt, many vaccines have been used including H5N1 and H5N2 with different homology percentage to the isolated strains in Egypt. Thus, expected circulation of the H5N1 strains is continuing since its introduction with genomic variations in both H5N1 high and low pathogenic strains. Such observations have been confirmed in Mexico after 13 years of using the inactivated vaccines (Escorcia et al., 2008). Indeed, enforced and restricted biosecurity with vaccination programme are the main measures to be applied in the control of AIV infections. Taken together, the present study clearly demonstrates the failure of the vaccines to induce immunogenic HI titers in more than 60% of the vaccinated birds which give the attention to the need for more control measures in many directions, the good quality vaccine and its application with enforced biosecurity are the main points to be considered.
Yet, data on the effect of vaccination on the transmission of virus within the flock indicate reduction in the reproduction ratio <1 by one week post vaccination (Van-Der-Goot et al., 2005). Thus we believe in vaccination and although it is widely believed that the emergence of new influenza pandemic caused by avian strain is only matter of time, a safe, effective and easy manufactured vaccine is required (Horimoto and Kawaoka, 2005). Vaccination using both conventional and recombinant vector vaccines has been experienced for the last ten years and data under field observation in China, Indonesia and Vietnam and recently in Egypt indicated some uncertainty in the performance of these vaccines. Moreover some experts have questions whether extensive use of recombinant vaccine results in the emergence of antigenic variants that have persisted in the region (Lee et al., 2005; Capua and Maragon, 2006). Recent alternative strategy in developing novel influenza vaccine proposed the use of NDV based live attenuated vaccine carrying the HA gene of HPAI wild bird isolate (Ge et al., 2007). This approach overcome the cost of production and the labor some administration which are the limitations of wide application of fowl pox based recombinant vaccine in the field. Although, generation of NDV expressing HA gene of H5N1 using reverse genetics demonstrate its potential use as bivalent vaccine against both pathogenic AI and pathogenic NDV. Yet, the vaccine efficacy may be affected by the replication ability of the vector used for recombination which in other studies by Swayne et al. (2003), only 40% protection was obtained. Also, the insertion of foreign gene in NDV genome may affect the immunogenicity and vaccine efficacy (Ge et al., 2007). In addition discrepancies in HI antibodies level was observed in 3 different studies (Veits et al., 2006; Park et al., 2006; Ge et al., 2007). Recent vaccination approach using live attenuated influenza A H5N1 candidate vaccine using reverse genetics provide promising finding under experimental study in mice and ferret, however a single dose did not induce complete protection against replication of challenge in the respiratory tract (Suguiten et al., 2006). Thus it is important to generate and carefully evaluate the live attenuated H5N1 vaccine because it is not known whether the vaccine will be associated with some residual virulence or over attenuated and it is difficult to obtain balance between attenuation and immunogenicity. Hence, it is important to generate novel approach in the development of new H5 vaccine.
In conclusion, the study proposes 7 log 2 HI titer using heterologous H5 antigen to face emerging H5N1 breaks. Also, the study recommends the use of heterologous antigen for evaluation of the efficacy of the used H5 AI vaccine. HPAI and LPAI of H5N1 virus are still circulating and evolving since its introduction in Feb. 2006 till now and no H7 viruses were detected along the 2 years of this study. Sequence analysis of the 11 detected H5 field viruses in Sharkia governorates confirmed their relatedness to H5N1 (Table 4).
| Table 4: | The obtained nucleotide sequences of the 11 detected avian influenza viral RNA in samples collected from mortalities in the vaccinated flocks during the study |
 | |
| All viruses were found to be H5N1 after sequence analysis | |
Suboptimal protection of the vaccinated chickens using different H5 AI vaccines considered one of the major possible causes of re-emerging of H5N1 breaks in 2007. Finally, HI titers of 7 log 2 can be used to represent the immunogenic capacity of the evaluated vaccine.
REFERENCES
- Capua, I. and D.J. Alexander, 2006. The challenge of avian influenza to the veterinary community. Avian Pathol., 35: 189-205.
Direct Link - Capua, I. and S. Marangon, 2003. The use of vaccination as an option for the control of Avian influenza. Avian Pathol., 32: 335-343.
Direct Link - Capua, I. and S. Marangon, 2006. Control of avian influenza in poultry. Emerg. Infect. Dis., 12: 1319-1324.
PubMedDirect Link - Chen, H., G. Deng, Z. Li, G. Tian and Y. Li et al., 2004. The evolution of H5N1 influenza viruses in ducks in southern China. Proc. Natl. Acad. Sci., 101: 10452-10457.
CrossRef - Chen, H., Y. Li, Z. Li, J. Shi and K. Shinya et al., 2006. Properties and dissemination of H5N1 viruses isolated during an influenza outbreak in migratory waterfowl in Western China. J. Virol., 80: 5976-5983.
Direct Link - Chen, H., G.J.D. Smith, S.Y. Zhang, K. Qin and J. Wang et al., 2005. Avian flu: H5N1 virus outbreak in migratory waterfowl. Nature, 436: 191-192.
Direct Link - Escorcia, M., L. Vazquez, S.T. Mendez, A. Rodriguez-Ropon, E. Lucio and G.M. Nava, 2008. Avian influenza: Genetic evolution under vaccination pressure. Virol. J., 5: 15-15.
Direct Link - Ge, J., G. Deng, Z. Wen, G. Tian and Y. Wang et al., 2007. Newcastle disease virus-based live attenuated vaccine completely protects chickens and mice from lethal challenge of homologous and heterologous H5N1 avian influenza viruses. J. Virol., 81: 150-158.
Direct Link - Horimoto, T. and Y. Kawaoka, 2005. Infuenza lessons from past pandemics, warning from current incidents. Nat. Rev. Microbiol., 3: 591-600.
Direct Link - Hussein, H.A., 2001. Molecular characterization of the Egyptian strain (Iman) of BVDV and its RNA synthesis in cell culture. Arab J. Biotech., 4: 23-36.
Direct Link - Lee, C.W., D.A. Senne and D.L. Suarez, 2004. Effect of vaccine use in the evolution of Mexican lineage H5N2 avian influenza virus. J. Virol., 78: 8372-8381.
CrossRefDirect Link - Lee, C.W., D.L. Suarez, T.M. Tumpey, H.W. Sung and Y.K. Kwon et al., 2005. Characterization of highly pathogenic H5N1 avian influenza A viruses isolated from South Korea. J. Virol., 79: 3692-3702.
Direct Link - Mase, M., K. Tsukamoto, T. Imada, K. Imai and N. Tanimura et al., 2005. Characterization of H5N1 influenza A viruses isolated during the 2003-2004 influenza outbreaks in Japan. Virology, 332: 167-172.
Direct Link - Stear, M.J., 2005. OIE manual of diagnostic tests and vaccines for terrestrial animals (mammals, birds and bees) 5th Edn. Volumes 1 and 2. World Organization for Animal Health 2004. ISBN: 9290446226. Parasitology, 130: 727-727.
CrossRefDirect Link - Park, M.S., J. Steel, A. Gracia-Sastre, D. Swayne and P. Palesa, 2006. Engineered viral vaccine constructs with dual specificity: avian influenza and Newcastle disease. Proc. Natl. Acad. Sci. USA., 103: 8203-8208.
Direct Link - Perdue, M.I., M. Gracia, D. Senne and M. Fraire, 1997. Virulance-associated sequence duplication at the hemagglutinin cleavage site of avian influenza viruses. Virus Res., 49: 173-186.
Direct Link - Shortridge, K.F., N.N. Zhou, Y. Guan, P. Gao and T. Ito et al., 1998. Characterization of avian H5N1 influenza viruses from poultry in Hong Kong. Virology, 252: 331-342.
Direct Link - Stone, H.D., 1987. Efficacy of avian influenza oil-emulsion vaccine in chicken of various ages. Avian Dis., 31: 483-490.
Direct Link - Suguiten, A.L., J. McAuuliffe, K.L. Mills, H. Jin and G. Duke et al., 2006. Live, attenuated influenza A H5N1 Candidate vaccines provide broad cross-protection in mice and ferrets. PLoS Med., 3: e360-e360.
Direct Link - Swayne, D.E., J.R. Beck, M. Gracia and D.H. Stone, 1999. Influence of virus strain and vaccine mass on efficacy of H5 avian influenza inactivated vaccines. Avian Pathol., 28: 245-255.
Direct Link - Swayne, D.E., M. Gracia, J.R. Beck, N. Kinney and D.L. Suarez, 2000. Protection against diverse highly pathogenic H5 avian influenza viruses in chicken immunized with a recombinant fowlpox vaccine containing an H5 avian influenza hemagglutinin gene insert. Vaccine, 18: 1088-1095.
Direct Link - Swayne, D.E., D.L. Suarez, S. Schultz-Cherry, T.M. Tumpey and D.J. King et al., 2003. Recombinant paramyxovirus type 1-avian influenza-H7 virus as a vaccine for protection of chicken against influenza and Newcastle disease. Avian Dis., 47: 1047-1050.
Direct Link - Taha, M., A.M. Ali, S.A. Nassif, A. Khafagy and E.A. El-Ebiary et al., 2008. Evolution of new escape mutant highly pathogenic avian influenza H5N1 viruses with multiple nucleotide polymorphism in Egypt December, 2007. Proceedings of the 2nd International Conference of Virology, April 2008, Dokki, Giza, Egypt.
- Trani, L., P. Cardioli, E. Falcon, G. Lombardi, A. Moreno, G. Sala and M. Tollis, 2003. Standardization of an Inactivated avian influenza vaccine and efficacy against A/chicken/Italy/1347/99 high pathogenicity virus infection. Avian Dis., 47: 1042-1046.
Direct Link - Van der Goot, J.A., G. Koch, M.C.M. de Jong and M. van Boven, 2005. Quantification of the effect of vaccination on transmission of Avian influenza (H7N7) in chickens. Proc. Natl. Acad. Sci. USA., 102: 18141-18146.
CrossRefDirect Link - Veits, J., D. Wiesner, W. Fucchs, B. Hoffmann and H. Granzow et al., 2006. Newcastle disease virus expressing H5 hemagglutinin gene protects chicken against Newcastle disease and avian influenza. Proc. Natl. Acad. Sci. USA., 103: 8197-8202.
Direct Link - Wan, X., T. Ren, K. Luo, M. Liao and G. Zhang et al., 2005. Genetic characterization of H5N1 avian influenza viruses isolated in southern China during the 2003-04 avian influenza outbreak. Arch. Virol., 150: 1257-1266.
Direct Link