
Jehan A.M. Gafer
Animal Health Research Institute, Dokki, Giza, Egypt
H.A. Hussein
Faculty of Veterinary Medicine, Cairo University, Giza, 12211, Egypt
I.M. Reda
Faculty of Veterinary Medicine, Cairo University, Giza, 12211, Egypt
International Journal of Virology
Year: 2009 | Volume: 5 | Issue: 1 | Page No.: 28-35







ABSTRACT
The present study was directed to isolate the PI-3 virus from sheep and goats which naturally affected with respiratory manifestation and to identify the isolated virus using several assays. Two hundred and five nasal swab samples were collected from diseased sheep and goats for the isolation of PI-3 virus. The samples were taken from diseased animal at 7 different Governorates Kaffer el sheikh, Alexandria, EL Behaira, Port Said, Demiatta El-Qalubia and Giza through the winter seasons of years 1999 to 2003. The virus was successfully isolated from three ovine samples after three successive passages on MDBK cells. The isolated viruses were then titrated and identified using different biological, serological and molecular assay.
PDF Abstract XML References Citation
How to cite this article
Jehan A.M. Gafer, H.A. Hussein and I.M. Reda, 2009. Isolation and Characterization of PI-3 Virus from Sheep and Goats. International Journal of Virology, 5: 28-35.
DOI: 10.3923/ijv.2009.28.35
URL: https://scialert.net/abstract/?doi=ijv.2009.28.35
DOI: 10.3923/ijv.2009.28.35
URL: https://scialert.net/abstract/?doi=ijv.2009.28.35
INTRODUCTION
Parainfluenza-3 (PI-3) is the most common virus infection of the respiratory tract of sheep and goats throughout the world. The virus belongs to family Paramyxoviridae of order Mononegaviralis, the non segmented negative single stranded RNA viruses (Pringle, 1991). The virus is generally spherical and 150 to 350 nm in diameter but can be pleomorphic in shape. The genomic RNA contained within a helical nucleocapsid that is enclosed within the lipid bilayer. Inserted into the envelope are glycoprotein spikes that extended approximately 8 to 12 nm from the surface of the membrane (Kingsbury, 1991; Field et al., 1996).
Parainfluenza-3 virus was first isolated from sheep in 1966 (Hore,1966) since then serological surveys have shown that this infection is widely spread in sheep population in many countries (Fenner et al., 1996). In Egypt, the first isolation of PI-3 virus from lambs was carried out in 1971 (Baz, 1971). The virus revealed symptoms like mild pyrexia, coughing and nasal discharge for several days and necropsy has shown area of pneumonia, particularly in ventral parts of the apical lobe (Hore and Stevenson, 1967). The present study was directed to isolate the PI-3 virus from sheep and goats which naturally affected with respiratory manifestation and to identify the isolated virus using several assays.
MATERIALS AND METHODS
Samples
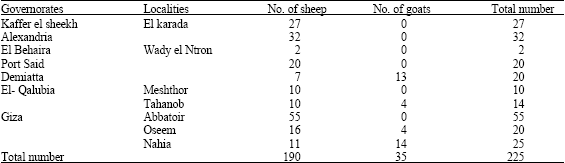
Two hundred and twenty five nasal swabs were collected from sheep and goats suffering from respiratory manifestation at 7 Govenorates through the winter seasons of years 1999 to 2003; the dilated data are shown in Table 1.
The samples were collected in sterile tubes containing 2 mL of sterile PBS and 200 μL of stock antibiotic solution per each sample. The samples were taken on ice bucket. Clarification was done by centrifugation at 5000 rpm for 5 min at 4°C in cooling centrifuge. The supernatant was taken and kept at -70°C till used.
| Table 1: | Number of nasal swab samples collected from sheep and goats in different localities in Egypt |
 | |
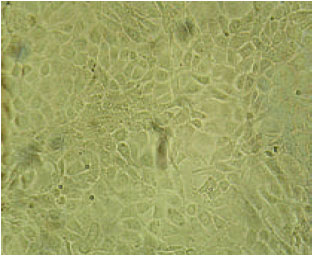 | |
| Fig. 1: | Normal non infected MDBK cells |
Viruses
First local bovine Egyptian PI-3 virus strain 45 was kindly supplied from Rinder Pest like disease Department at Veterinary Serum and Vaccine production Institute Abbassia, Cairo. Second bovine PI-3 virus strain SF4 was kindly supplied from Rabies Department in VACSERA.
Cells
MDBK cells were kindly supplied from Department of Virology, Faculty of Veterinary Medicine Cairo University (Fig. 1).
Heamagglutination Test
Samples were subjected to heamagglutination test to exclude the non-heamaglutinatinating samples from the isolation work. The test was conducted according to Singh and Baz (1966) using 1% G. pig RBCs.
Isolation of PI-3 Virus from HA Positive Samples
The isolation was carried out according to Rulka (1987) using MDBK cells. Three successive passages were done for this isolation.
Titeration of Isolates
The harvest of the inoculated MDBK cells after three passages of the isolates was titrated. The test was performed according to Frey and Liess (1971). Briefly, in a microliter plate 90 μL of MEM free serum were dispensed into each well then 10 μL of the suspected isolate suspension were added to the wells of the 1st column and serial ten fold dilutions were made 50.000 cells/well were added while, being in suspension. The plate was incubated at 37°C in CO2 incubator with daily microscopic examination. The TCID50 was calculated according to Reed and Muench (1938).
Identification by Heamadsorption Technique
The test was conducted according to Elizabeth et al. (1997). After 16 h of inoculation of cells with isolates, the attached cells were fixed for 30 min with formaldehyde in PBS (pH 7.2) then 1% suspension of G. pig RBCs was added. The cells were examined under inverted microscope.
Identification of PI-3 Virus by Detecting Inclusion Bodies
The test was made using Zinker Solution as a fixative according to Clayden (1971).
Identification of the Isolates Using Immunofluorescence
The test was performed according to Vander Heide (1971) using Cell Culture Staining Chamber (CCSC). Each cup was inoculated with 100 μL of the isolate and uninfected cups were included as control. The cups were overlaid with 100 uL of anti PI-3 serum conjugated with FITC (supplied from Department of Virology, Faculty of Veterinary Medicine Cairo University) then examined under fluorescent microscope.
Identification of PI-3 Suspected Isolates by Virus Neutralization Test
The test was performed according to Murakami et al. (1983). Beta neutralization procedure using antibovine PI-3 hyperimmune serum (supplied from Serum and Vaccine Research Institute Abbassia, Cairo). The serum was originally supplied from Imes, Iowa, USA.
Detection of PI-3 Virus by Polymerase Chain Reaction
The genomic RNA was extracted from inoculated tissue culture by means of TRIZOL reagent according to their manufacture instructions (commercially supplied by life technologies CO.) The TRIZOL reagent is improvement to single step RNA isolation method was declared by Chomczynski and Sacchi (1987).
RT-PCR was employed using oligonucleotide primer pair designed from conserved site of fusion protein gene (F gene ) (Table 2) as previously described by Lyon et al. (1997). The sequences of utilized primers as follow:
Analysis of the PCR Product Using Agar Gel Electrophoresis
The analysis was carried out according to Sambrook et al. (1989) using ethidium bromide stained agarose gel 1.5%.
| Table 2: | Cycling protocol for amplification of F gene of PI-3 |
 | |
RESULTS AND DISCUSSION
Results of heamagglutination test are presented in (Table 3). Five HA-positive samples were cultivated and propagated on MDBK cells. Three passages in cell culture were applied and resulted three isolates which were designated as Oseem56/1999, Oseem184/2003 and Nahia70/2000 according to their localities and the year at which the samples were collected (Table 4) present the designation and passages results of the cell culture work. The three isolates as well as local bovine strain 45 were titrated according to infectivity the results presents in (Table 5). It seem to be titers ranged from 105 to 106.6 TCID50/mL with the highest test titer was given by the isolate Oseem56/1999 followed by isolate Oseem 184/2003 and finally the isolate Nahia 70/2000. The isolated viruses were identified using hemadsorption, inclusion body detection, immunofluorescence, SNT as well as RT-PCR (Fig. 2).
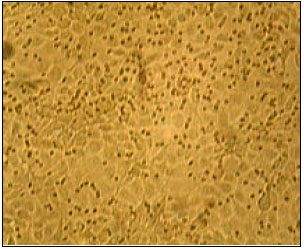
Parainfluenza-3 (PI-3) virus causes, different clinical syndromes in sheep and goat specially respiratory infections (Ackermann and Brodgen, 2000). Serological surveys have shown that infection is widely spread among sheep population in many countries (Fishman, 1965; Pop et al., 1979; Saddour, 1987; Riedmann et al., 1991). Many of serological data indicated that the ovine PI-3 virus is antigenically related to bovine and human strains of PI-3 (Wood et al., 1975). In the present study, we tried to isolate PI-3 virus from naturally infected sheep and goats and RT-PCR was used as a recent tool for detection and identification of the isolated virus. The detection of PI-3 virus by heamagglutination, revealed five positive samples (all of them were from sheep samples representing 2.2% of the 225 collected samples. This low percentage of HA positive may be due to the high amount of virions (at least 105.5-106.5 virions) required to give one HA unit. The isolation was achieved through three passage applied on MDBK cells giving rise to three isolates developed a clear CPE (Table 6).The identification by detecting inclusion bodies revealed large eosinophilic intracytoplasmic (Fig. 3) with no evidence to nuclear inclusions in contrast to bovine PI-3 virus that produced both cytoplasmic and nuclear inclusion similar results are previously reported by Howard et al. (1982). Intracytoplasmic perinuclear florescence in the positive FA-isolates on MDBK cells shown in Fig. 4. It was noticed the three isolates were from lambs which may be indicate the prevalence of Parainfluenza-3 (PI-3) virus that among sheep than in goats.
| Table 3: | Haemagglutination titer of the five HA positive nasal swab samples and their localities |
 | |
| Table (4) | Results of cultivation of the HA positive (ovine) nasal swab samples |
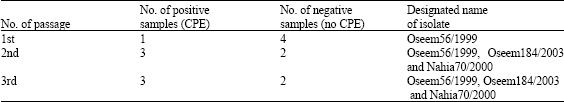 | |
| CPE = cytopathic effect | |
| Table 5: | Results of titration of PI-3 isolates on MDBK cells using infectivity test |
 | |
 | |
| Fig. 2: | Infected MDBK cells with the isolates after fixation and the positive heamadsorption |
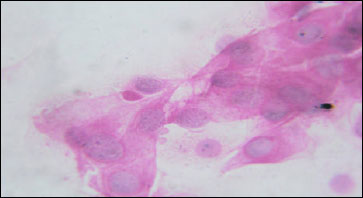 | |
| Fig. 3: | Large eosinophilic intracytoplasmic inclusion body |
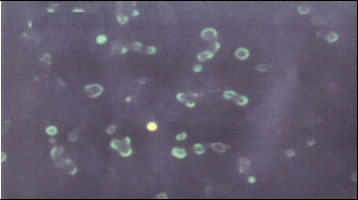 | |
| Fig. 4: | Intracytoplasmic perinuclear fluorescence in the positive FA-isolates on MDBK cells |
| Table 6: | Results of virus neutralization test for PI-3 isolated viruses against antiparainfluenza-3 reference serum |
 | |
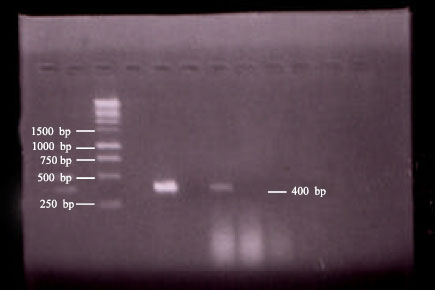 | |
| Fig. 5: | The ethidium bromide stained 1.5% agarose gel electrophoresis of the PCR products. Lane M: The molecular DNA marker. Lane A: The amplified PCR product of approximately 400 bp, the conserved portion of F gene of PI-3 virus strain A45 (positive control). Lane 1- 3: The PCR products of the isolated viruses. Only Oseem56/1999 revealed the expected band (lane 1) |
However, the failure to isolate the virus from goat may be due to the lower number of collected samples from goats compared with those from sheep and the fact that the animals were kept under relatively isolated conditions (Erasmus et al., 1967). The low titer of Oseem184/ 2003 isolate may be taken as criterion for heterogeneity (Gaul et al., 1982). This finding agreed with many of the earlier studies, which have shown that at least three ovine PI-3 strains of different serotypes namely (G2 strain, CsL6 strain and DH-1 strain), do exist (Hore, 1966; St-George, 1969; Howard et al., 1982). The heamadsorption used as an indicator system for the presence of PI-3 virus proved to be more rapid and sensitive than waiting until the cytopathic changes appear. This finding is similar to those described previously by St-George (1969) and Toth and Jankura (1990). The RT-PCR technique waw performed in this study using two primers chosen from conserved site of fusion protein gene (F gene of whate PI-3 viruses). The primers were chosen in this site of genome as it is conserved among PI-3 viruses not specific for ovine PI-3 virus (Fig. 5) Also, both sheep and goats can catch bovine PI-3 virus. Moreover, these primers could help us in identification of the standard bovine PI-3 virus strain and the standardization of the technique. Also, the same primers were developed by Lyon et al. (1997) and Maria et al. (1998). The detection and differentiation of PI-3 virus could be studied by amplification and sequencing of another part of the genome for example HN gene (Ruth et al., 1994; Echevarria et al., 2000; Vecherov et al., 2003). Indeed, This study reports the isolation and characterization of parainfluenza-3 virus from sheep and goats in Egypt.
REFERENCES
- Ackermann, M.R. and K.A. Brogden, 2000. Response of ruminant respiratory tract to Mannheimia (pasterella) heamolytica. Microbes Infect., 2: 1079-1088.
Direct Link - Chomczynski, P. and N. Sacchi, 1987. Single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction. Anal. Biochem., 162: 156-159.
CrossRefPubMedDirect Link - Echevarria, J.E., D.D. Erdman, H.C. Meissner and L. Anderson, 2000. Rapid molecular epidemiologic studies of human PI-3 virus based on direct sequencing of amplified DNA from a multiplex RT-PCR assay. J. Virol. Method, 88: 105-109.
Direct Link - Elizabeth, J.H., G. Paul and W. Richard, 1997. The bovine PI-3 haemagglutinin,neuraminadase,glycprotein expressed in baculovirus protect against experimental BPIV-3 challenge. Vaccine, 15: 730-738.
Direct Link - Fishman, H.R., 1965. Presence of neutralizing antibody for myxovirus parainfluenza-3 in sheep sera. Proc. Soc. Exp. Biol. Med., 118: 725-727.
PubMedDirect Link - Lyon, M., C. Leroux, T. Greenland, J. Chastang, J. Patet and J.F. Mornex, 1997. Presence of unique PI-3 strain identified by RT-PCR in visna-maedi virus infected sheep. Vet. Microbiol., 51: 95-104.
Direct Link - Maria, Z., B. Tim, J.S. Carol, M.G. John and N.W. Katherine, 1998. Moleucular epidemiology of consecutive outbreaks of parainfluenza-3 in bone marrow transplant unit. J. Clin. Microbiol., 6: 2289-2293.
Direct Link - Reed, L.J. and H. Muench, 1938. A simple method of estimating 50 percent end points. Am. J. Epidemiol., 27: 493-495.
Direct Link - Riedmann, S., M.I. Montecions, N. Tadich and G. Reinhardt, 1991. Serological survey for antibodies to parainfluenza-3 virus in sheep in chile. Vet. Rec., 128: 572-572.
Direct Link - Ruth, A.K., L.F. Jean, B.B. Linda, H.Y. Robert, 1994. Rapid detection of parainfluenza virus type 3 RNA in respiratory specimen. Use of reverse transcription-PCR-EIA. J. Clin. Microbiol., 32: 484-488.
Direct Link - Sambrook, J., E.F. Fritsch and T.A. Maniatis, 1989. Molecular Cloning: A Laboratory Manual. 2nd Edn., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York, USA., ISBN-13: 9780879695774, Pages: 397.
Direct Link - Toth, T.E. and D. Jankura, 1990. Analysis of bovine parainfluenz-3 reblication in bovine embryonic lung cell by in direct fluorescent antibody and haemadsorption assays. J. Virological Methods, 27: 113-119.
Direct Link - Vecherov, A.E., P.K. Aianot, A.M. Timina and V.V. Lisitsin, 2003. Detection and differentiation of bovine parainfluenza-3 virus strains studied by amplification and sequencing of the HN gene. Vopr. Virusol., 48: 46-59.
Direct Link