
T. Cao
Noveltech, LLC., 16600 43rd Ave N., Minneapolis, MN 55446, USA
J.T. Weil
Center of Excellence for Poultry Science, University of Arkansas, Fayetteville, AR 72701, USA
P. Maharjan
Center of Excellence for Poultry Science, University of Arkansas, Fayetteville, AR 72701, USA
J. Lu
Syngenta, 410 S. Swing Rd., Greensboro, NC 27409, USA
C.N. Coon
Center of Excellence for Poultry Science, University of Arkansas, Fayetteville, AR 72701, USA
LiveDNA: 1.35097
International Journal of Poultry Science
Year: 2020 | Volume: 19 | Issue: 5 | Page No.: 210-218







ABSTRACT
Background and Objective: The requirement of sulfur amino acids for laying hens have been determined but the method in which methionine is regulated has not been studied. The aim of this research was to study the hepatic methionine-metabolizing enzymes and metabolites in laying hens. Materials and Methods: Five hundred forty Dekalb-XL laying hens were housed and fed a control diet until sampling. On day of sampling, six hens were sacrificed at each time period to allow for determination of hepatic enzymatic activities and metabolite concentrations during light and dark periods. Data was analyzed using the general linear models (GLM) procedure with statistical analysis software (SAS). Results: The enzymes and metabolites showed cyclical changes related to light and dark periods. During the light period of the day, layers showed elevated activities of methionine s-adenosyltransferase (EC 2.5.1.6; MAT), cystathionine ß-synthase (EC 4.2.1.22; CS) and cystathionase (EC 4.4.1.1; C-ase) and depressed activities of betaine-homocysteine methyltransferase (EC 2.1.1.5; BHMT) and N5methyltetrahydrofolate-homocysteine methyltransferase (EC 2.1.1.13; MFMT), as compared to the dark period of the day. The hens also had a decreased methionine to cysteine ratio (Met/Cys ratio), an increased methylation ratio (s-adenosylmethionine to s-adenosylhomocysteine ratio; SAM/SAH ratio) and an increased cystathionine (CYST) concentration in the liver during the light period. Conclusion: The changes of the enzymatic activities and metabolite concentrations suggest that the methionine metabolism of laying hens during the light period was in favor of methionine degradation through cysteine synthesis. Alternatively, the metabolism of hens during the dark period was in favor of methionine conservation by limiting the conversion of methionine to cysteine. Thus, feeding hens a higher cysteine diet several hours before lights are turned off may prove beneficial to counteract the limited cysteine synthesis from dietary methionine during the dark period of the day.
PDF Abstract XML References Citation
Copyright: © 2020. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
T. Cao, J.T. Weil, P. Maharjan, J. Lu and C.N. Coon, 2020. The Diurnal Changes of Hepatic Enzymes and Metabolites of Methionine Metabolism in Laying Hens. International Journal of Poultry Science, 19: 210-218.
DOI: 10.3923/ijps.2020.210.218
URL: https://scialert.net/abstract/?doi=ijps.2020.210.218
DOI: 10.3923/ijps.2020.210.218
URL: https://scialert.net/abstract/?doi=ijps.2020.210.218
INTRODUCTION
Sulfur amino acid nutrition for laying hens has been one of the major interests of poultry nutritionists for many decades. Common sulfur amino acids including but not limited to, methionine (Met) and cysteine (Cys), are of great importance in poultry nutrition due to their crucial role in feather formation, metabolism and protein synthesis. Additionally, cysteine can be converted into end products such as taurine and glutathione1. Although critical for metabolism, methionine is the first limiting amino acid. Cysteine follows closely behind, as it can be synthesized through Met via transsulfuration pathways.
While many studies have estimated the requirement of methionine or total sulfur amino acids (TSAA) for laying hens, there is little information about sulfur amino acid metabolism, especially the mechanisms by which the metabolism is regulated2-9. Studies with rats have demonstrated that methionine metabolism is a cycle with a unidirectional outlet formed by the cystathionine synthesis reaction (Fig. 1)10. Methionine can be endogenously metabolized to cysteine; however, the opposite is not possible due to the lack of necessary enzymes. It has been found that methionine-metabolizing enzymes are rich in tissues of broiler chicks11. Additionally, the liver has been shown to contain higher activities of all the major enzymes involved in methionine metabolism and is considered an ideal organ for the study of methionine metabolism12.
Researchers determined that dietary cysteine reduces the methionine requirement of rats, the sparing effect of dietary cysteine for methionine has been reported in rats, fish, pigs, humans, broilers and laying hens13-21,10,22-26. The sparing effect may suggest there is a precise regulatory mechanism of methionine metabolism. Studies with rats have shown the distribution of homocysteine between remethylation and cystathionine synthesis which may provide a major regulatory site10,15. The competition between protein synthesis and the formation of S-adenosylmethionine for available methionine may provide a second potential regulatory site10,27-30. The theory for the existence of a major regulatory site is still premature, especially when applied to poultry.
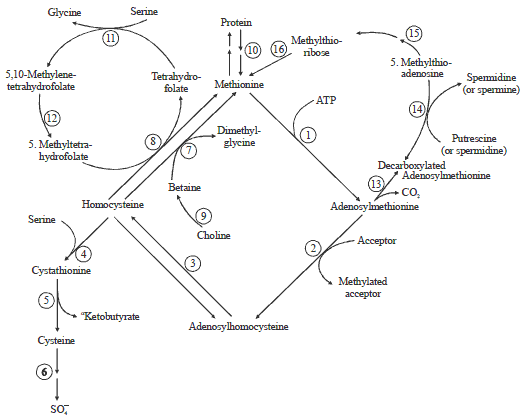 | |
| Fig. 1: | Methionine metabolism in mammals |
The numbers represent the following enzymes or sequences; 1: Methionine adenosyltransferase (EC 2.5.1.6), 2: AdoMet-dependent transmethylations, 3: Adenosylhomocysteinase (EC 3.3.1.1), 4: Cystathionine-b-synthase (EC 4.2.1.22), 5: g-cystathionase (EC 4.4.1.1), 6: Further metabolism of cysteine, 7: Betaine-homocysteine methyltransferase (EC 2.1.1.5), 8: Methylfolate-homocysteine methyltransferase (EC 2.1.1.13), 9: Choline + betaine aldehyde dehydrogenases (EC1.1.99.1 and EC1.2.1.8), 10: Equilibrium between free and protein methionine, 11: Serine hydroxymethylase (EC 2.1.2.1), 12: Methylenetetrahydrofolate reductase (EC 1.7.99.5), 13: AdoMet decarboxylase (EC 4.1.1.50), 14: Spermidine (spermine) synthase (EC 2.5.1.16 and EC 2.5.1.22), 15-16: Methylthioadenosine phosphorylase (EC 2.4.2.28)+methionine formation via methylthioribose-1-phosphate. Pathway drawing of methionine and cysteine metabolism is from the publication of Finkelstein29 | |
The poultry industry has two different production systems for producing meat and eggs, namely broilers and layers. Laying hens are subjected to environmental changes and physiological variations during a 24 h period. Hens, during the light period of a day, are actively consuming feed and are inactive during the dark period of the day. Furthermore, researchers have shown that light affects reproduction, behavior and also activates the development of vitamins and enzymes31,32. Considering the changes in light and dark periods each day, the periodic nutrient delivery to the liver may stimulate changes in methionine metabolism. Additionally, the demand for amino acids may vary during the day for the periodic needs, for egg protein synthesis, particularly albumen synthesis33,34. Unlike yolk protein, which is continuously synthesized in the liver and accumulated in the ovum until ovulation occurs, the albumen is synthesized in the magnum and supplied to the egg during a 3-3.5 h period when the ovum is in the magnum of the oviduct32. The periodic need of amino acids for albumen synthesis may, in turn, influence amino acid metabolism. Wurtman35 discussed the advantages of using a daily rhythm of intracellular enzyme activities as a model system to study amino acid metabolism. Diurnal changes of enzyme activities are better indications of parallel alterations in the fates of their substrates with physiological significance. The variations of the enzyme activities may provide evidence about how the enzymes are regulated in the body. To the best of the author's knowledge, there have been no reported studies on the diurnal changes of methionine-metabolizing enzymes. The studies on the diurnal changes of rat liver SAM and SAH concentrations have shown that the levels of the metabolites oscillate significantly during the day, suggesting that the activities of the enzymes in the metabolism may differ during this time36,37. In order to study the regulatory mechanism of methionine metabolism, an experiment to study the daily variation of hepatic methionine-metabolizing enzymes and metabolites with laying hens was conducted.
MATERIALS AND METHODS
All procedures regarding the use of live animals in this study were carried out in accordance with the Animal Use Protocol #03008, which was approved by the University of Arkansas Institutional Animal Care and Use Committee.
Animal care and sample collection: Dekalb-XL laying hens, 69 weeks of age, were fed a diet (Table 1) containing 2,899 kcal ME kg–1, 19.5% CP with 0.81% digestible TSAA (0.5% methionine and 0.31% cystine).
| Table 1: | Control diet composition of ingredients and nutrient analysis |
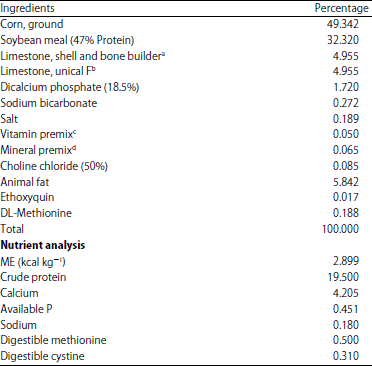 | |
aLimestone, Shell and Bone Builder, had mean particle size of 3260.61 microns and consited of 39. 63% Ca (Shell and Bone Builder, ILC Resources, Des Moines, Iowa). bLimestone, Unical F, had mean particle size of 2166.85 microns and consisted of 39.53% Ca (Unical F, ILC Resources, Des Moines, Iowa). cThe vitamin premix provided the following per kilogram of diet; Vitamin A: 3,300 IU, Vitamin D3: 3,307 IU, Vitamin E: 40.6 IU, Vitamin K: 1.46 mg, Pantothenic acid: 23.06 mg and Vitamin B12: 0.017 mg. dThe mineral premix provided the following in milligram per kilogram of diet; Mn: 91.88, Fe: 0.027, Zn: 82.65, Cu: 9.48 and I: 1.04. eNonnutritive filler (cellulose), Brown Co., Berlin, NH | |
Analysis for percent digestible amino acids for methionine and cysteine in diets was performed as discussed by Ekmay et al.38. Hens were housed at 21°C and were provided a daily photoperiod of 16 h and 15 min light (4:00-20:15) and 7 h and 45 min dark (20:15-4:00). Six hens were sacrificed every 3 h by carotid exsanguination over a 24 h period. Immediately after removal from the body cavity, livers were frozen and stored in liquid nitrogen for further processing. A group of 40 hens from the same treatment were used to determine the hourly feed intake.
Enzymes and metabolites assay: The enzyme extracts were prepared following the procedure introduced by Finkelstein and Martin39. A Tissuemizer® was used to homogenize liver samples in 4 volumes of ice-cold 1 mM potassium phosphate, pH 7.5, at 85% maximum speed for 1 min. After the homogenate was centrifuged at 27,000 g for 20 min at 4°C, the supernatant was decanted into Sephadex G-25 columns for the removal of small molecular weight solutes through filtration. The protein-containing fraction was then lyophilized and stored at -40°C until further analysis. All assays in this experiment were conducted in less than 10 days, which is the maximum time that enzyme activity has been shown to remain unchanged39,40. Immediately before the enzyme assay, the enzyme extract was reconstituted with water and the final volume was adjusted such that 0.5 mL was the equivalent of 1 g of wet tissue. The enzyme activities were determined with the published methods described by Mudd et al.40 for MAT and CS, by Mudd et al.41 and Finkelstein et al.42 for MFMT, by Greenberg43 for C-ase and by Finkelstein and Mudd14 with a modification in determining the product formed in the assay (Cao and Coon11 for BHMT.
The metabolite extract was prepared following the procedure described by Cao and Coon11. Liver tissue with 3 volumes of 0.2 M HClO4 was homogenized with a Tissuemizer® at 85% maximum speed for 1 minute at 4°C. After the homogenate was centrifuged at 27,000 g for 10 min, the supernatant was filtered through a 0.22 μm Millex-GS Millipore filter. The filtrate was treated for blocking thio-group of metabolite44. Fifty μL of 250 mM 2-mercaptoethanol in 700 mM HClO4 and 67 μL of 6 N NaOH were added to 767 μL filtrate. After mixing, 50 μL of 200 mM iodoacetic acid was added to each sample. After 30 min of standing, 66 μL of 6 N HCl was added to each sample. Concentrations of methionine, S-adenosylmethionine, S-adenosylhomocysteine, homocysteine (Hcy), cystathionine and cysteine were then analyzed for each treated sample by HPLC11. Due to the reaction of blocking thio-groups of metabolites occurred due to the presence of 2-mercaptoethanol, the metabolites of oxidized forms may proceed to their corresponding reduced forms. The determined concentration was the total amount of metabolite including both oxidized and reduced forms.
Statistical analysis: Data were analyzed using analysis of variance (ANOVA), followed by Duncan’s Multiple-Range (DMR) test using the statistical analysis system (SAS45). Differences of p<0.05 were considered statistically significant.
RESULTS AND DISCUSSION
The liver activities of methionine-metabolizing enzymes and concentrations of metabolites of laying hens over a 24 h period are presented in Table 2 and 3, respectively. Hepatic enzyme activities of MAT, BHMT, CS and C-ase showed significant variations during the day with a non-significant change of enzyme MFMT. Additionally, metabolites in liver, as well as Met, SAM, SAH, Hcy, Cysta and Cys, showed significant variations during the day.
Met:Cys ratio: Dietary Met:Cys ratio is subject to a change in the liver after the transportation of the amino acid to the liver through the portal vein. The change of the ratio is expected, mainly due to the extensive metabolism occurring in the organ because the liver contains the highest specific activities of the methionine-metabolizing enzymes in layers, broilers and other species10,30. Due to the minimal contribution of amino acids from the aorta, there is only slight possibility that the blood supply from the aorta will change the Met:Cys ratio46. Figure 2 shows that the liver Met:Cys ratio (between 0.2 and 1.2) is lower than the dietary ratio (1.61). The lower Met:Cys ratio in the liver is an indication that the liver functions as a degradation site of methionine for cysteine.
| Table 2: | Activity variations of methionine-metabolizing enzymes over 24 h in laying hen livers (nmoles min–1 g–1 wet liver) |
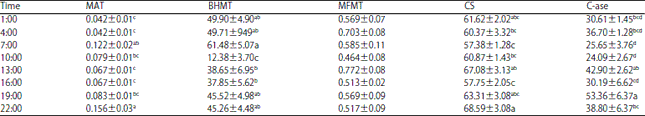 | |
Each value is Means±SEM (n = 6). a-dMeans within a column with no common superscript differ significantly (p<0.05) | |
| Table 3: | Concentration variations of methionine-metabolizing metabolites over 24 h in laying hen livers (nmoles g–1 wet liver) |
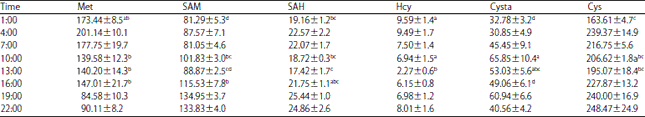 | |
Each value is Means±SEM (n = 6). a-dMeans within a column with no common superscript differ significantly (p<0.05) | |
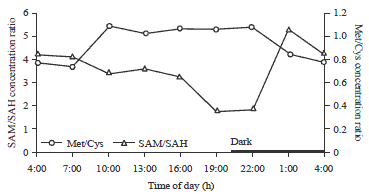 | |
| Fig. 2: | Diurnal changes of transmethylation reactions and the conversion of methionine to cysteine in laying hen livers |
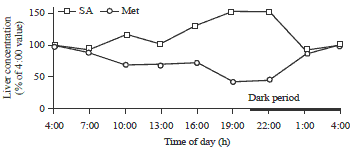 | |
| Fig. 3: | Diurnal changes of liver methionine and SAM concentrations |
The progressive decrease of the ratio in liver from 0.84-0.36 from 7:00-22:00 coincided with the feed consumption activities from 4:00-20:15 with delayed responses. The lower ratio of SAM/SAH during the light period of the day suggests an increased conversion of methionine to cysteine, as the liver degrades the extra methionine supplied in the diet. Conversely, the elevated ratio during the dark period is an indication that the conversion of methionine to cysteine is significantly reduced in order to conserve methionine from catabolism. The conservation of methionine that occurs during this period is a result of the lack of amino acid consumption at this time.
The daily rhythm of the Met:Cys ratio is positively correlated with liver methionine concentrations (R2 = 0.802) and negatively correlated to liver cysteine concentrations (R2 = -0.563). Figure 3 shows that liver methionine concentration level changed in an opposite direction as the SAM level over the 24 h period with the magnitude of change being maintained at the same level for both metabolites. Considering SAM is strictly produced endogenously from methionine, the opposite movement of the liver methionine and SAM concentrations, as observed during the light period, is a result of the reaction catalyzed by the enzyme MAT. This was confirmed by the elevated MAT activities from 4:00-22:00 during the day (Fig. 4).
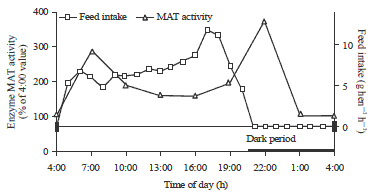 | |
| Fig. 4: | The relationship between MAT activity and feed intake (feed intake was determined every hour with 40 laying hens) |
The enzymatic MAT activity showed an earlier peak of 2.9 fold higher at 7:00 and a second peak of 3.7 fold higher at 22:00 than the value at 4:00. Figure 4 also shows that the MAT activity was possibly stimulated by feed consumption. Layers showed an earlier feed consumption peak at 6:00 with the second and the largest peak at 18:00. The peak of MAT activity at 7:00 (Table 2) was delayed about 1 h later than the earlier feed consumption peak, while the second peak of MAT activity was delayed approximately 5 h later than the second fee consumption peak. The different responding times of MAT activity to feed consumption may be explained by the function of the crops of laying hens. Immediately before the dark period, hens consumed additional feed. With a large amount of feed in the crop, a longer delay and greater response of the enzyme MAT was observed when comparing the second feed consumption peak to the first feed consumption response (Fig. 4).
Transmethylation: In mammals, the transmethylation reactions are controlled primarily through the methylation ratio (SAM/SAH)10,30,47. A higher ratio may lead to an increased rate of transmethylation reactions. Figure 2 demonstrates that the SAM/SAH ratio was elevated from 10:00-22:00 with a responding delay of about 3 h to the feeding period. The increased SAM/SAH ratio is an indication of increased transmethylation reaction rates in livers of laying hens during the light period of the day. These increased transmethylation reactions most likely occurred in order to remove the elevated SAM level in livers since there was an increased SAM (Table 3) formation from methionine in the system during this period. There is also a possibility that the need for the methyl donor to other biologically important materials (DNA, phospholipids and proteins) was increased during this period48. The decrease in SAM/SAH ratio, observed during the dark period, is likely a response to counteract the lack of methionine supplied to the hen.
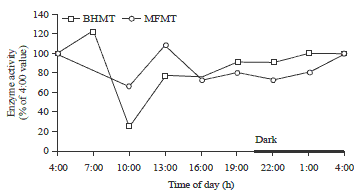 | |
| Fig. 5: | Diurnal changes of liver enzyme activities of BHMT and MFMT of laying hens |
Increased SAH or decreased SAM as observed during 4:00-7:00, resulting in lower ratio of SAM/SAH, can also be associated with DNA hypomethylation and thus the process tends to play a role in preserving SAM levels49. The observations of diurnal changes in SAM:SAH ratio as found in this study are consistent with other studies in mice models48,50.
Methionine remethylation: BHMT and MFMT are two methionine-conserving enzymes in the methionine metabolic pathway29,30. Both enzymes catalyze the remethylation reactions of homocysteine for methionine resynthesis. The present study found that both BHMT and MFMT showed decreasing activities during the light period (hens were actively consuming feed) and increasing activity during the dark period (hens were inactive) (Fig. 5). The enzyme activity was significantly lower at 10:00 for BHMT (25% of the 4:00 value) (p<0.05). Although the MFMT activities were not significantly different over the 24 h period, the lower activity at 10:00 (66% of 4:00 value) tended to show a similar change as enzyme BHMT. The decreased activities of the two methionine-conserving enzymes during the light period may have occurred in order to limit methionine resynthesis from homocysteine, keeping the homocysteine from further degradation through the transsulfuration reactions. The increased activity of the two enzymes during the dark period may be physiologically significant for the methionine conservation since dietary methionine was limited during this period. Methyl transferase activity could be regulated by cellular SAM content, where it was reported that the higher activity of methyltransferase could be observed in methionine-deprived conditions51.
Transsulfuration: Figure 6 shows elevated cystathionine concentrations in the livers of hens during the light period of the day as compared to the livers of hens during the dark period. The cystathionine level peaked at 10:00 and 19:00 during the day.
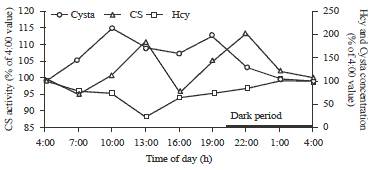 | |
| Fig. 6: | Diurnal changes of CS activity and concentrations of Hcy and Cysta |
The two peaks of cystathionine concentration coincided with the two peaks of SAM (Fig. 6). Finkelstein10,30 has suggested that the SAM is an activator of the enzyme CS. In this experiment, the enzyme CS had a very large Kd (dissociation constant) value (Kd = 1-25 mM) compared to the methionine-conserving enzymes BHMT and MFMT (Kd = 0.002 and 0.06 mM, respectively). This would suggest that without a significant increase of homocysteine concentration and CS activity in the system, the production of cystathionine from homocysteine could still be significantly increased. In the current study, the CS activity peaks were observed at 13:00 and 22:00. The three hour delay of the peaks of CS activity, compared to the peaks of cystathionine concentration, is an indication that the peaks of cystathionine concentration were not due to the increased level of enzyme CS. The surges of cystathionine levels at 10:00 and 22:00 are likely due to the increased SAM, the activator of enzyme CS and the larger Kd value of the enzyme. The increased CS activities of hens during the light period of the day were possibly induced by the higher levels of SAM, causing a sustained function of the enzyme to keep the higher rate of cystathionine formation from homocysteine30,52. The increased formation of cystathionine synthesis of hens during the light period of the day is supported by the increased homocysteine consumption in livers (Fig. 6). The lower level of homocysteine during the light period was caused by the increased CS activities, due to enzyme activation and the increase in enzyme quantity. This may further suggest that the reaction of homocysteine to cystathionine is regulated through the CS activity rather than through the substrate to product ratio. The decreased cystathionine formation because of the reduced enzyme CS activity during the dark period is another indication that methionine metabolism was in favor of methionine conservation when there was no amino acid consumption.
Enzyme C-ase is an alternative transsulfuration enzyme that catalyzes the reaction of the cystathionine to cysteine29,30. The enzyme showed decreased activities from 4:00-10:00 followed by two separate increases at 13:00 and 19:00.
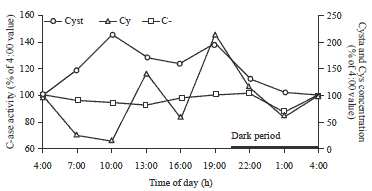 | |
| Fig. 7: | Diurnal changes of C-ase activity and the concentrations of related metabolites of laying hen livers |
During the dark period, the C-ase activity returned to the 4:00 value (Fig. 7). Since the enzyme C-ase is also involved in reactions other than the transsulfuration reaction, the variation of the enzyme over the 24 h period cannot be completely explained by the change of the transsulfuration reactions53,54. However, the two-peak surge of C-ase activity of hens during the light period of the day in the present study is possibly related to the surges of SAM concentration, cystathionine concentration and CS activity. The C-ase activity peak (Fig. 7) at 13:00 is likely related to the higher CS activity, while the peak at 19:00 could be related to higher cystathionine and SAM concentrations. The lower C-ase activity from 4:00-10:00 could be the continuation of the lower activity observed during the dark period. In general, there was an increased C-ase activity in livers of hens during the light period of the day, as compared to livers of the hens during the dark period. This could be an additional indication that during the light period, methionine degradation was favored as opposed to cysteine synthesis.
Although, the current findings are crucial to better understanding methionine metabolism in laying hens, further research should be conducted in order to ensure reproducibility. To the best of the author’s knowledge, the diurnal changes of methionine-metabolizing enzymes have not been examined; therefore, future studies should be conducted in order to examine the influence of hormones on enzymatic activities during the light and dark periods. Additionally, various light intensities should be examined, considering light affects the hen’s reproduction, behavior and development of enzymes31,32. Daily variations of hepatic enzymes and metabolites related to the total sulfur amino acids exist, causing the need for additional experimentation.
CONCLUSION
The change of the enzyme activities and metabolite concentrations over the 24 h period in the present study suggests that the methionine metabolism of laying hens during the light period of the day is in favor of the methionine degradation through cysteine synthesis, while the metabolism of layers during the dark period tends to conserve methionine by limiting the conversion of methionine to cysteine. This may indicate that hens can be fed a lower cysteine diet during the early feeding hours of the day, considering dietary methionine can be efficiently converted to cysteine. Feeding hens a higher cysteine diet a few hours before dark periods may be beneficial because the feed kept in crops may provide hens sufficient cysteine for egg protein synthesis even though the conversion of methionine to cysteine is limited during the dark period.
SIGNIFICANCE STATEMENT
This study discovered the regulatory mechanism for which methionine metabolism is controlled in laying hens. This study will help the researcher understand specific pathways and subsequent control mechanisms regarding methionine metabolism as related to hens housed in light and dark production settings.
REFERENCES
- Bin, P., R. Huang and X. Zhou, 2017. Oxidation resistance of the sulfur amino acids: Methionine and cysteine. BioMed Res. Int., Vol. 2017.
CrossRefDirect Link - Fisher, C. and T.R. Morris, 1970. The determination of the methionine requirement of laying pullets by a diet dilution technique. Br. Poult. Sci., 11: 67-82.
CrossRefDirect Link - Jensen, L.S., L. Falen and G.W. Schumaier, 1974. Requirement of white leghorn laying and breeding hens for methionine as influenced by stage of production cycle and inorganic sulfate. Poult. Sci., 53: 535-544.
CrossRefDirect Link - Schutte, J.B. and E.J.V. Weerden, 1978. Requirement of the hen for sulphur‐containing amino acids. Br. Poult. Sci., 19: 573-581.
CrossRefPubMedDirect Link - Parsons, C.M. and R.W. Leeper, 1984. Choline and methionine supplementation of layer diets varying in protein content. Poult. Sci., 63: 1604-1609.
CrossRefDirect Link - Calderon, V.M. and L.S. Jensen, 1990. The requirement for sulfur amino acid by laying hens as influenced by the protein concentration. Poult. Sci., 69: 934-944.
CrossRefDirect Link - Schutte, J.B., J. de Jong and H.L. Bertram, 1994. Requirement of the laying hen for sulfur amino acids. Poult. Sci., 73: 274-280.
CrossRefPubMedDirect Link - Strathe, A.B., A. Lemme, J.K. Htoo and E. Kebreab, 2011. Estimating digestible methionine requirements for laying hens using multivariate nonlinear mixed effect models. Poult. Sci., 90: 1496-1507.
CrossRefDirect Link - Finkelstein, J.D., 1998. The metabolism of homocysteine: Pathways and regulation. Eur. J. Pediatr., 2: S40-S44.
CrossRefPubMedDirect Link - Mato, J.M. M.L. Martínez-Chantar and S.C. Lu, 2013. S-adenosylmethionine metabolism and liver disease. Ann. Hepatol., 12: 183-189.
CrossRefDirect Link - Womack, M. and W.C. Rose, 1941. Partial replacement of dietary methionine by cystine for purposes of growth. J. Biol. Chem., 141: 375-379.
Direct Link - Sowers, J.E., W.L. Stockland and R.J. Meade, 1972. L-methionine and L-cystine requirements of the growing rat. J. Anim. Sci., 35: 782-788.
CrossRefDirect Link - Finkelstein, J.D. and S.H. Mudd, 1967. Trans-sulfuration in mammals: The methionine sparing effect of cystine. J. Biol. Chem., 242: 873-880.
Direct Link - He, J.Y., B. Han, L.X. Tian, H.J. Yang, S.L. Zeng and Y.J. Liu 2016. The sparing effect of cystine on methionine at a constant TSAA level in practical diets of juvenile Nile tilapia Oreochromis niloticus. Aquacult. Res., 47: 2031-2039.
CrossRefDirect Link - Baker, D.H., W.W. Clausing, B.G. Harmon, A.H. Jensen and D.E. Becker, 1969. Replacement value of cystine for methionine for the young pig. J. Anim. Sci., 29: 581-584.
CrossRefDirect Link - Roth, F.X. and M. Kirchgessner, 1987. Biological efficiency of dietary methionine or cystine supplementation with growing pigs. J. Anim. Physiol. Anim. Nutr., 58: 267-280.
CrossRefDirect Link - Roth, F.X. and M. Kirchgessner, 1989. Influence of the methionine: Cysteine relationship in the feed on the performance of growing pigs. J. Anim. Physiol. Anim. Nutr., 61: 265-274.
CrossRefDirect Link - Chung, T.K. and D.H. Baker, 1992. Maximal portion of the young pig's sulfur amino acid requirement that can be furnished by cystine. J. Anim. Sci., 70: 1182-1187.
CrossRefDirect Link - Buono, M.D., L.J. Wykes, D.E.C. Cole, R.O. Ball and P.B. Pencharz, 2003. Regulation of sulfur amino acid metabolism in men in response to changes in sulfur amino acid intakes. J. Nutr., 133: 733-739.
CrossRefDirect Link - Graber, G. and D.H. Baker, 1971. Sulfur amino acid nutrition of the growing chick: Quantitative aspects concerning the efficacy of dietary methionine, cysteine and cystine. J. Anim. Sci., 33: 1005-1011.
CrossRefDirect Link - Graber, G., H.M. Scott and D.H. Baker, 1971. Sulfur amino acid nutrition of the growing chick: Effect of age on the capacity of cystine to spare dietary methionine. Poult. Sci., 50: 1450-1455.
CrossRefDirect Link - Sasse, C.E. and D.H. Baker, 1974. Sulfur utilization by the chick with emphasis on the effect of inorganic sulfate on the cystine-methionine interrelationship. J. Nutr., 104: 244-251.
CrossRefDirect Link - Soares, J.H.Jr., 1974. Experiments on the requirement of inorganic sulfate by the chick. Poult. Sci., 53: 246-251.
CrossRefDirect Link - Schutte, J.B., E.J.V. Weerden and H.L. Bertram, 1984. Protein and sulphur amino acid nutrition of the hen during the early stage of laying. Arch. Geflugelk, 48: 165-170.
Direct Link - Stipanuk, M.H. and N.J. Benevenga, 1977. Effect of cystine on the metabolism of methionine in rats. J. Nutr., 107: 1455-1467.
CrossRefDirect Link - Young, V.R., 1987. 1987 McCollum award lecture. Kinetics of human amino acid metabolism: nutritional implications and some lessons. Am. J. Clin. Nutr., 46: 709-725.
CrossRefDirect Link - Finkelstein, J.D., 1990. Methionine metabolism in mammals. J. Nutr. Biochem., 1: 228-237.
CrossRefDirect Link - Finkelstein, J.D., 2000. Pathways and regulation of homocysteine metabolism in mammals. Semin. Thromb. Hemost., 26: 219-225.
CrossRefPubMedDirect Link - Biyatmoko, D., 2014. Effects the combinations of light color and intensity of light to age at first laying and production egg of alabio laying ducks. Int. J. Biosci., 5: 80-85.
CrossRefDirect Link - Er, D., Z. Wang, J. Cao and Y. Chen, 2007. Effect of monochromatic light on the egg quality of laying hens. J. Applied Poult. Res., 16: 605-612.
CrossRefDirect Link - Hiramoto, K., T. Muramatsu and J. Okumura, 1990. Protein synthesis in tissues and in the whole body of laying hens during egg formation. Poult. Sci., 69: 264-269.
PubMedDirect Link - Pens, Jr. A.M. and L.S. Jensen, 1991. Influence of protein concentration, amino acid supplementation and daily time of access to high- or low-protein diets on egg weight and components in laying hens. Poult. Sci., 70: 2460-2466.
CrossRefPubMedDirect Link - Wurtman, R.J., 1969. Time-dependent variations in amino acid metabolism: Mechanism of the tyrosine transaminase rhythm in rat liver. Adv. Enzyme Regul., 7: 57-67.
CrossRefDirect Link - Sánchez, V.C., R. Hernández-Muñoz, L. Sánchez, S. Vidrio, L. Yáñez and J. Suárez, 1991. Twenty-four-hour changes of S-Adenosylmethionine, S-Adenosylhomocysteine adenosine and their metabolizing enzymes in rat liver; possible physiological significance in phospholipid methylation. Int. J. Biochem., 23: 1439-1443.
CrossRefDirect Link - Schalinske, K.L. and R.D. Steele, 1992. Variations in S-adenosylmethionine, S-adenosylhomocysteine and adenosine concentrations in rat liver. Biofactors, 3: 265-268.
PubMedDirect Link - Ekmay, R.D., M. De Beer, S.J. Mei, M. Manangi and C.N. Coon, 2013. Amino acid requirements of broiler breeders at peak production for egg mass, body weight and fertility. Poult. Sci., 92: 992-1006.
CrossRefDirect Link - Finkelstein, J.D. and J.J. Martin, 1984. Methionine metabolism in mammals. Distribution of homocysteine between competing pathways. J. Biol. Chem., 259: 9508-9513.
PubMed - Mudd, S.H., J.D. Finkelstein, F. Irreverre and L. Laster, 1965. Transsulfuration in mammals: Microassays and tissue distribution of three enzymes of the pathway. J. Biol. Chem., 240: 4382-4392.
Direct Link - Mudd, S.H., H.L. Levy and G. Morrow, 1970. Deranged B12 metabolism: Effects on sulfur amino acid metabolism. Biochem. Med., 4: 193-214.
CrossRefDirect Link - Finkelstein, J.D., W.E. Kyle and B.J. Harris, 1971. Methionine metabolism in mammals. Regulation of homocysteine methyltransferases in rat tissue. Arch. Biochem. Biophys., 146: 84-92.
CrossRefDirect Link - Greenberg, D.M., 1962. Cystathionine and homoserine cleavage: Cystathionine + H2O → Cysteine + α-ketobutyric acid + NH3 Homoserine + H2O → α-KetobuCyric acid + NH3. Methods Enzymol., 5: 936-942.
CrossRefDirect Link - Cooper, J.D.H. and D.C. Turnell, 1982. Fluorescence detection of cystine by o-phthalaldehyde derivatisation and its separation using high-performance liquid chromatography. J. Chromatogr. B: Biomed. Sci. Applic., 227: 158-161.
CrossRefDirect Link - Garcia, R.A.G. and M.H. Stipanuk, 1992. The splanchnic organs, liver and kidney have unique roles in the metabolism of sulfur amino acids and their metabolites in rats. J. Nutr., 122: 1693-1701.
CrossRefDirect Link - Hoffman, D.R., D.W. Marion, W.E. Cornatzer and J.A. Duerre, 1980. S-adenosylmethionine and S-adenosylhomocysteine metabolism in isolated rat liver. J. Biol. Chem., 255: 10822-10827.
Direct Link - Xia, L., S. Ma, Y. Zhang, T. Wang, M. Zhou, Z. Wang and J. Zhang, 2015. Daily variation in global and local DNA methylation in mouse livers. PloS One, Vol. 10, No. 2.
CrossRefDirect Link - Caudill, M.A., J.C. Wang, S. Melnyk, I.P. Pogribny and S. Jernigan et al., 2001. Intracellular S-adenosylhomocysteine concentrations predict global DNA hypomethylation in tissues of methyl-deficient cystathionine β-synthase heterozygous mice. J. Nutr., 131: 2811-2818.
CrossRefDirect Link - Sengupta, A., S.D. Rhoades, E.J. Kim, S. Nayak, G.R. Grant, P. Meerlo and A.M. Weljie, 2017. Sleep restriction induced energy, methylation and lipogenesis metabolic switches in rat liver. Int. J. Biochem. Cell Biol., 93: 129-135.
CrossRefDirect Link - Tisdale, M.J., 1980. Changes in tRNA methyltransferase activity and cellular S-adenosylmethionine content following methionine deprivation. Biochim. Biophys. Acta (BBA)-Nucl. Acids Protein Synth., 609: 296-305.
CrossRefDirect Link - Lu, S.C., 2000. S-adenosylmethionine. Int. J. Biochem. Cell Biol., 32: 391-395.
CrossRefPubMedDirect Link - Matsuo, Y. and D.M. Greenberg, 1958. A crystalline enzyme that cleaves homoserine and cystathionine. I. isolation procedure and some physicochemical properties. J. Biol. Chem., 230: 545-560.
PubMedDirect Link - Matsuo, Y. and D.M. Greenberg, 1959. A crystalline enzyme that cleaves homoserine and cystathionine. IV. Mechanism of action, reversibility and substrate specificity. J. Biol. Chem., 234: 516-519.
Direct Link