
Amal A. Mouhamed
Department of Poultry Diseases, Animal Health Research Institute, Agriculture Research Center, Giza, Egypt
Moemen A. Mohamed
Department of Poultry Diseases, Faculty of Veterinary Medicine, Assiut University, Assiut, Egypt
Bakheet M. Bakheet
Department of Poultry Diseases, Faculty of Veterinary Medicine, Assiut University, Assiut, Egypt
Khaled A. Aziz El-Din
Department of Poultry Diseases, Animal Health Research Institute, Agriculture Research Center, Giza, Egypt
Chang-Seon Song
Avian Disease Laboratory, College of Veterinary Medicine, Konkuk University, Seoul, Republic of Korea
LiveDNA: 82.32514
International Journal of Poultry Science
Year: 2020 | Volume: 19 | Issue: 5 | Page No.: 193-209







ABSTRACT
Background and Objective: Newcastle disease (ND) is a highly contagious viral disease of avian species and represents a major threat to the poultry industry worldwide. Regardless of which type of vaccine is used, birds are still able to become infected by NDV and can transmit the disease to others. This study aimed to obtain improved understanding of the variety and interrelationships of NDV isolates. Materials and Methods: A total of 116 tissue/organ samples were subjected to virus isolation and pathogenicity assessment in vivo by intracerebral pathogenicity index (ICPI) determination. Molecular characterization was performed by one-step RT-PCR to obtain a 535 bp fragment, including the fusion gene cleavage site. The purified PCR products of 12 isolates were selected for DNA sequencing. Results: Nucleotide and deduced amino acid sequence analysis of the cleavage site of the F gene of all field isolates revealed the motif 112R-R-Q-R-R-F117 at the C-terminus of the F2 protein and F (phenylalanine) at the N-terminus of the F1 protein (residue 117), indicating that these strains were velogenic. The nucleotide sequence analysis revealed that our isolates showed the greatest nucleotide identities (99.3%) with the velogenic strains from Jordan, Israel and Turkey, suggesting that the virus circulating in Egypt probably extends from the Middle Eastern region. Phylogenetic analysis showed that our isolates could be classified into three genotypes (VIId, VIIa and II), indicating that VIId is the predominant circulating genotype in Egypt, where 10 isolates were clustered. One isolate for genotype VIIa and one for genotype II were also observed. A low evolution rate, with Ka/Ks ratios ranging from 0.01-0.02, indicated negative or purifying selection. The minimum evolutionary distance detected was 0.09 to genotype VIId, whereas the maximum distance was 0.21 to genotype II, from which most commercial live virus vaccine strains are derived. Conclusion: The control of NDV by vaccination still faces new challenges and evaluating the effectiveness of the current commercial vaccine strains against circulating NDV strains has become a necessity.
PDF Abstract XML References Citation
Copyright: © 2020. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Amal A. Mouhamed, Moemen A. Mohamed, Bakheet M. Bakheet, Khaled A. Aziz El-Din and Chang-Seon Song, 2020. Molecular Studies on Newcastle Disease Virus Isolates in Relation to Field Vaccine Strains in Egypt (2012-2015). International Journal of Poultry Science, 19: 193-209.
DOI: 10.3923/ijps.2020.193.209
URL: https://scialert.net/abstract/?doi=ijps.2020.193.209
DOI: 10.3923/ijps.2020.193.209
URL: https://scialert.net/abstract/?doi=ijps.2020.193.209
INTRODUCTION
Newcastle disease (ND) is an economically significant disease whose listed notifiable disease status with OIE marks its importance to both commercial poultry producers and poultry-trading countries1. Newcastle disease virus (NDV) belongs to the genus Avulavirus in the family Paramyxoviridae and it is a well-characterized member among the avian paramyxovirus serotypes that belong to serotype 12. The genome of NDV consists of six transcriptional units (3 -N-P-M-F-HN-L-5 )3. The F and HN proteins form spike-like projections on the outer surface of the viral envelop and are the neutralizing and protective antigens of NDV. The F protein is synthesized as an inactive precursor (F0) that is cleaved by a host cell protease into two biologically active F1 and F2 subunits4. Cleavage of the F protein is a prerequisite for virus entry and cell-to-cell fusion. The sequence of the F protein cleavage site is well characterized as a major determinant of NDV pathogenicity in chickens5,6.
All NDV strains belong to a single serotype7,8. However, genetic and antigenic diversity are recognized for NDV isolates and the detection of progressive changes in strains isolated over successive years indicates that NDV is a continually evolving virus9,10. Based on genome length and F gene sequence, strains of NDV have been classified into class I (nine genotypes, 1-9) and class II (eleven genotypes, I-XI). Class I strains are usually avirulent. A recent study also proposed that class I viruses comprise a single genotype, while class II includes 15 genetic groups, including 10 previously established (I-IX and XI) and five new genotypes (X, XII, XIII, XIV and XV)11. Class II includes both virulent and avirulent strains and the avirulent vaccine strains LaSota and B1in present use worldwide. Currently, the predominant circulating strains associated with disease outbreaks worldwide are categorized as genotypes V, VI and VII of class II12-15.
Most outbreaks of NDV arise in non-vaccinated susceptible animals and therefore, countries worldwide currently maintain a stringent vaccination policy7. Regardless of which type of vaccine is used, birds are still able to be infected by NDV and can transmit the disease to others. Because vaccination cannot prevent disease transmission or virus shedding and the disease can occur in vaccinated birds9,14, the role of vaccinations limited to safeguarding the individual bird from significant disease by providing protective antibodies that can quickly respond to the introduction of an ND virus9,16,17.
However, any strain of NDV can be used as a vaccine because all NDVs are in one serotype and all vaccines should prevent clinical disease and death from ND. However, some studies have elucidated that vaccines formulated with strains more homogenous to the challenge virus can decrease the shedding amount of challenge virus in oropharyngeal swabs from vaccinated birds and significantly decrease the number of birds that shed virus, as the goal of vaccination should be to decrease shedding two to three logs parallel to non-vaccinated animals18-21.
NDV strains with varying degrees of virulence circulate among avian species2. Small genomic changes resulting from replication errors can result in alterations in virulence. For example, insertion of basic amino acids in the fusion gene cleavage site can confer the virus the ability to systemically replicate and give rise to severe disease. Molecular changes should be monitored to analyze alterations in the cleavage site, which can identify a potential increase in virulence20.
Our study aims to obtain improved understanding of the variety and interrelationships of NDV isolates through the following: (1) Isolation and biological characterization of NDV isolates by the intracerebral pathogenicity index (ICPI) in one-day-old chicks, (2) Identification of the NDV pathotypes by rapid detection of the fusion (F) gene using RT-PCR and (3) F gene sequencing of the region covering the cleavage site of the F protein of isolates to show the genotype that our isolated strains belong to and the degree of divergence (lineages) between the lentogenic vaccines used (HB-1 and LaSota) and the isolated field strains. These data will allow us to determine the degree of similarity that may provide information to gain improved protection that reflects on reducing virus shedding and transmission.
MATERIALS AND METHODS
Sample collection: This study was conducted from 2012-2015 on 116 samples (organs and tissues) from commercial chicken flocks processed either separately or as a pool, since the brain and intestinal samples were processed separately from other samples22. Mortality was observed in commercial chicken farms between 18-23 days old, ranging from 20-100% in some flocks, with deaths occurring within 24-48 h after the onset of clinical signs. The morbidity was as high as 95-100% in some flocks.
Virus isolation: Virus was isolated following the protocol of the World Organization for Animal Health (OIE) and European standards (OIE23; CE, 92/66) and organ samples were prepared and treated to obtain a bacterial -free suspension ready for embryonated egg inoculation, as described in Terregino and Capua22,24,25.
NDV Detection and Identification: The harvested allantoic fluid was tested for haemagglutination activity (HA); if no HA activity was detected following two passages in eggs, the sample was considered negative. When HA activity was detected, the presence of bacteria was excluded by culturing24.
Haemagglutination inhibition test (HI test) (α procedure: diluted serum constant antigen): Since the HI test is both qualitative and quantitative, a known amount of antigen, in most cases 4HA units, must be used24. Therefore, we prepared hyperimmune sera against NDV using 15 adult healthy chickens according to the method described by Iqbal et al.26. The chickens were selected from flocks where avian influenza vaccines were not used. Serum from all the birds was collected and observed for haemagglutination activity after inactivation at 56°C for 30 min.
Five out of 15 birds were kept as controls, while the remaining 10 were injected intramuscularly with NDV (LaSota strain+mineral oil) according to the schedule described in Table 1. Blood samples from the birds were taken periodically to observe their antibody titer against NDV by HI test. The chickens were bled after having the highest antibody titer and blood was collected aseptically without anticoagulant. Serum was separated from the clotted blood and purified and antisera were aliquoted and stored at -20°C27. The haemagglutination inhibition test was according to the protocol of Terregino and Capua24.
Evaluation of pathogenicity using intracerebral pathogenicity indices: A simple method for recording results and calculating indices is shown in Table 2. Any APMV-1 giving a value of 0.7 or greater in an ICPI test was counted as a virulent ND virus. The ICPI is the mean score per bird per observation over the 8-day period. The most virulent isolates have an ICPI close to 2.0 and lentogenic and asymptomatic enteric viruses have values of 0.0-0.625.
Molecular characterization of NDV isolates
RNA extraction: The extraction protocol recommended by the manufacturer was followed using a Thermo Scientific GeneJet Viral DNA and RNA Purification Kit (#k0821, Thermo Scientific)28.
Partial F gene analysis of the isolated ND strains: One- step reverse transcriptase- polymerase chain reaction (RT-PCR) for the extracted viral RNA samples was performed as follows. PCR amplification and sequencing were performed using the following degenerate primers, 5-ATG GGC (C/T)CC AGA (C/T)CT TCT AC-3 (forward) and 5 - CTG CCA CTG CTA GTT GTG ATA ATC C-3 (reverse), specific to the fusion (F) protein gene29,30. The one- step RT-PCR quick-start protocol using a QIAGEN® One Step RT-PCR Kit (cat. No. 210210) was followed.
Gel extraction and DNA sequencing: Specific bands for the F gene were excised and purified by using a QIAquick Gel Extraction Kit (cat. No. 28706) according to the manufacturer’s instructions. The purified DNA encoding the F region was sequenced by direct sequencing in one direction. Sequencing reactions were completed using an ABI PRISM® BigDye™ Terminator Cycle Sequencing Ready reaction kit v2.0 (Perkin Elmer, USA) and an automated DNA sequencer (ABI PRISM® 377 DNA Sequencer).
Nucleotide and deduced amino acid sequence analysis: The percentages of nucleotide identity and sequence editing were determined using the BioEdit software package version 7.01. Nucleotide analysis, prediction of amino acid sequences and alignments were performed by Molecular Evolutionary Genetics Analysis version 4.01 (MEGA 4). Detection of synonymous and non-synonymous substitution rates was carried out using the Nei-Gojobori method in MEGA 4.
| Table 1: | Inoculation schedule |
 | |
| Table 2: | Determination of the intracerebral pathogenicity index (ICPI) |
 | |
10 birds observed for 8 days = 80 observations. Index = Mean score per bird per observation = 109/80 = 1.36 | |
Nucleotide sequences of partial F genes of studied isolates were later deposited in GenBank31.
Phylogenetic analysis: The substantiality of the genetic grouping and topology of a phylogenetic tree was affirmed by comparing F gene sequences of our isolates and published sequences relevant to class I and class II NDVs. The phylogenetic tree was structured using selected sequences. Phylogenetic analysis was carried out by comparing the variable portion of the F gene between 47-581 (535 bp) nucleotide sequences using the Clustal W multiple alignment method with Lasergene31. The phylogenetic tree was structured using the neighbor-joining method after 1,000 bootstrap replicates. The representative strain sequences from each genotype and different geographical areas were retrieved from GenBank, as shown in the phylogenetic tree.
Evolutionary study: Selective pressure for the putative F protein was determined using synonymous non-synonymous analysis program (SNAP) services (http://hcv.lanl.gov/content/sequence/SNAP/SNAP.html)32. The ratio of nonsynonymous to synonymous substitutions (Ka/Ks ratio) for each amino acid site in the F- coding region was used to scan for evidence of positive or negative selection.
RESULTS
Detection and Identification of NDV: One hundred isolates were identified from 116 samples collected from euthanized diseased chickens or recently deceased chickens by isolation in embryonated chicken eggs (ECEs). Embryo deaths were recorded between the 2nd and 5th days post inoculation.
Haemagglutination Inhibition test: The positive HA samples underwent a haemagglutination inhibition (HI) test using a previously prepared ND-antiserum. Out of 102 HA positive samples, 100 (98.04%) samples were inhibited by the ND antiserum.
Characterization of NDVs
In vivo pathotyping of NDVs isolated by ICPI: Fifteen HI-positive samples underwent the biological characterization of NDV by using the intracerebral pathogenicity index (ICPI) for classification as lentogenic, mesogenic or velogenic. The deaths in the first 24 h were considered non-specific. The chicks injected intracerebrally showed clinical signs 3-5 days post injection (Fig. 1). The chicks that died during the period of the test underwent post -mortem examination (Fig. 2).
 | |
| Fig. 1(a-f): | The clinical signs appeared on chicks after IC inoculation by different isolates of NDV: (a) General signs (dullness, closed eyes and ruffled feathers) and dropped wings, (b) Petechial haemorrhages on the toes of both legs, © Twisting of the head and neck, (d) Dropping of wings and Paralysis of both wings with lateral recumbency, (e) Paralysis of both legs with lateral recumbency and (f) Opisthotonus |
 | |
| Fig. 2(a-f): | The postmortem lesions which were observed after Intracerebral inoculation of chicks by different isolates of NDV: (a) White granules around the eye (unilateral), (b) Congestion of the blood vessels of brains (both cerebrum and cerebellum), © Yellow edema covering the neck from the cerebellum, (d) Liquefied brain, (e) Congested inflamed kidneys with ureate deposition in both ureters and (f) Enteritis all over the small and large intestine |
 | |
| Fig. 3(a-c): | Agarose gel electrophoresis (1%) of amplified partial fusion gene (535 bp fragment) of NDV field isolates that used after for sequencing, M: 100 bp DNA ladder |
| Table 3: | The ICPI values and pathotypes |
 | |
The results were as follows: 6.7% of the isolates had an ICPI value >1.5-2 for the highly virulent NDV isolates (velogenic), whereas 93.3% had lower values ranging from 0.5-1.5 for the moderately virulent isolates (mesogenic), out of the maximum value of 2 (Table 3).
Molecular characterization of the isolated NDV strains: Twelve samples were examined and a partial 535 bp fragment of the fusion gene was successfully amplified (Fig. 3).
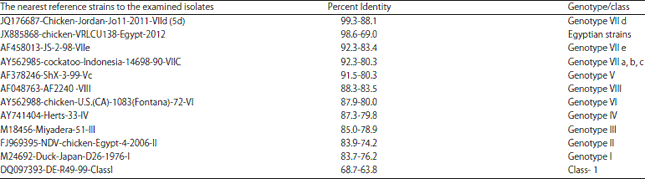
| Table 4: | The percent of identity between the examined strains and reference strains from representing classes and genotypes |
 | |
Partial nucleotide sequencing of the F gene region covering nucleotide positions 47-581 was performed for the 12 examined strains.
Proteolytic cleavage site analysis of the F0 protein: The F protein cleavage site revealed the participation of three basic amino acid residues at positions 113, 115 and 116 at the F2 protein and a phenylalanine at position 117 at the N-terminus of the F1 protein for all strains, which means that all the isolates were categorized into the velogenic group with the motif 112R-R-Q-R-R-F117.
Nucleotide analysis: In specific comparison with the Egyptian strains, our isolates showed the highest similarity percentages with JX885868-chicken-VRLCU138-Egypt-2012 (98.6%), JX173098-NDV-chicken-Giza-Egypt-MR0-2012(98.2%) and KP209305-NDV-chicken-Egypt-FA-8-2014(95.8%), followed by a lesser degree of homology with FJ969394-chicken-Egypt-3-2006 (84,6%), JX193771-NDV-chicken-Behaira-Egypt-MR6-2012 (83.3%) and JX193772-NDV-chicken-Fayoum-Egypt-MR7-2011 (73.6%).
The highest similarity detected between the examined isolates and reference strains from genotype VIId ranged from 99.3% to88.1%, while the greatest nucleotide identity (99.3%) was recorded with a velogenic strain from Jordan (JQ176687-chicken-Jordan-Jo11-2011-VIId (5d).In addition, the examined isolates were highly similar to JN979564-Turkey-Israel-111-2011-Israel VIId (5d), FJ608338-SH-Zhejiang-06-VIId, DQ486859-GM-VIId, AY630436-Kr-188-02-VIId, AY630438-ch-501-02-VIId, DQ439937-NDV05-085-VIId and AF358786-TW-2000-VIId, with nucleotide sequence homology ranging from 95.6-88.1%).
The homologies of nucleotide sequences were relatively low with the other genotypes, with the lowest similarity recorded with genotype I (83.7-76.2%) and genotype II (83.9-74.2%). Live virus vaccines used commercially are derived from those genotypes where the low similarity was recorded with commercially available vaccines; AY170137-Komarov-45-LK-II was 81.9%- 74.9%, AF077761-LaSota-II was 82.2- 74.8%, M24696-Lassato-46-II was 82.2-74.8%, Y18898-CLONE30-II was 82.2-74.8% and M24695-HB1-47-II was 82.7-75.2%.
Seven major epitopes have been characterized, including the fusion inhibition and neutralization of the F- protein. Individual amino acids at residues 72, 74, 75, 78, 79 and 343 and a stretch of amino acids from residues 157-171 have been shown to be critical for both the structures and functions of the F-gene.
Analysis of these seven neutralizing epitopes located at positions 72, 74, 75, 78, 79, 157-171 of the F protein showed an F (phenylalanine) for K (lysine) amino acid substitution at position 78 in all strains. No recombination events were recognized but a point mutation in one of the neutralizing epitopes (F78 for K) of the F-protein was distinct in the field isolates.
Nucleotide similarity: Nucleotide similarity percentages between the 12 field isolates and representative strains of different classes and genotypes were determined (Table 4).
Amino acid analysis: Shown in Fig. 4.
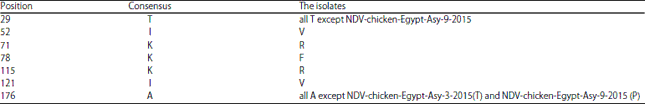
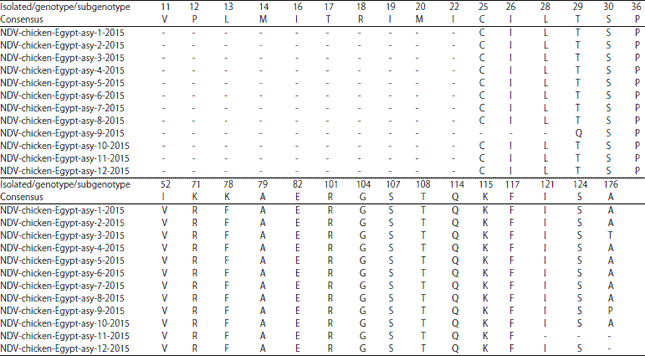
Amino acid substitutions: Summarized in Table 5 and 6.
Phylogenetic analysis: A phylogenetic tree was constructed using 121 partial genome nucleotide sequences of representative viruses categorized as class I and class II (genotypes I-XIII and XVI) available at GenBank. The results revealed that most of the examined isolates (10 strains) were in class II and of subgenotype VIId, with one strain in subgenotype VIIa and another in genotype II, as shown in the phylogenetic tree (Fig. 5).
 | |
| Fig. 4: | Amino acid alignment of 12 NDV field isolates. continued colour in the columns indicates position where the sequence is identical to that of the reference strain. The unshaded cells show the amino acids differences among examined isolates |
| Table 5: | Amino acids substitutions for the examined isolates |
 | |
Alignment of the partial F gene nucleotide sequences revealed that the 10 isolates grouped under genotype VIId shared 99.3-88.1% nucleotide identity with NDV strains of genotype VIId. Additionally, the nucleotide similarity for the isolate belonging to genotype VIIa was 92.3-80.3% with the global strains of this genotype and the strain belonging to genotype II had similarity ranging from 83.9-74.2% with the reference strains of the same genotype.
Evolutionary distance analysis: Evolutionary distance analysis revealed that the maximal genetic distances were between our isolates and previously established strains from genotype II (0.21), represented by strain JN193503-NDV-ostrich-Ismailia-2010-II. For example, the evolutionary distance between our isolates and the vaccine strainsY18898-CLONE30-II, M24696-Lassato-46-II, JF950510-LaSota-2010-II, EU289029-VG-GA-II and AY289002-VGGA-87-II was 0.20. The evolutionary distances showed that our strains were significantly closer to strains from genotype VIId than to the vaccine strains, with minimal genetic distances of 0.09. The most closely related strains from this genotype to our isolates were JQ176687-chicken-Jordan-Jo11-2011-VIId (5d), JN979564-turkey-Israel-111-2011-Israel VIId (5d), FJ608338-SH-Zhejiang-06-VIId, DQ486859-GM-VIId, DQ659677-strain-NA-1-VIId and AY135754-1SACK00184-Saudi Arabia-2003-VIId (5d), with evolutionary distances ranging from 0.01-0.08.
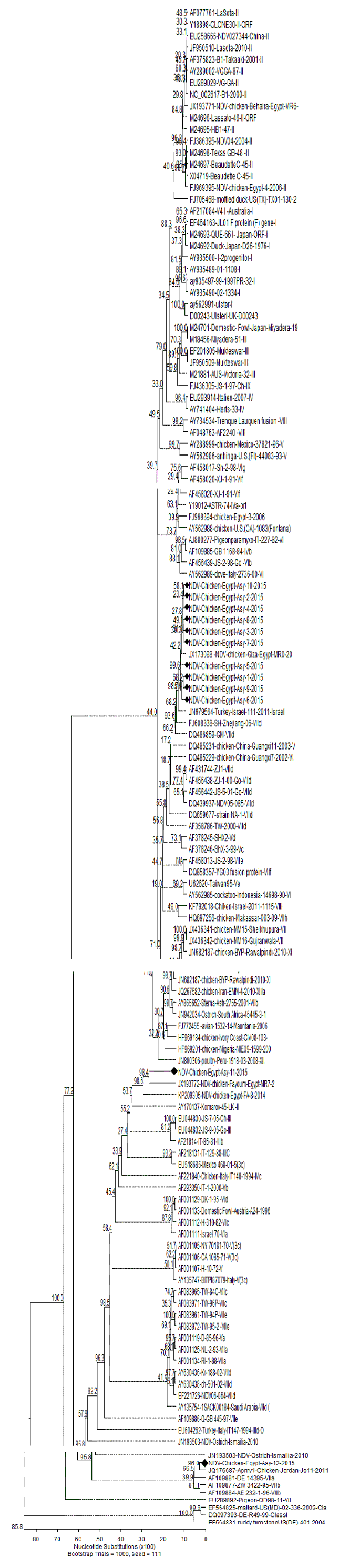 | |
| Fig. 5: | Phylogenetic analysis and relationship among 12 NDV field strains on basis of partial Fusion-protein gene nucleotide sequences between position 47 and 581. Phylogenetic relatedness of the 12 NDV field strains with representative NDV strains from class I and class II different genotypes identified by their GenBank accession numbers |
| Table 6: | Amino acid residue substitution of F gene for NDV strains of different genotypes |
 | |
Evolutionary distances: The evolutionary distance between amino acid sequences of the partial fusion protein gene of our strains and representative strains from different classes and genotypes was determined (Table 7).
Ka/Ks ratio: The Ka/Ks ratio ranged from 0.01-0.02, which is a value less than one indicating negative or purifying selection.
| Table 7: | Estimation of evolutionary distance between amino acid sequences of the partial fusion protein gene of our strains and representing strains from different classes and genotypes |
 | |
| Table 8: | Ka/Ks ratio of 12 field strains |
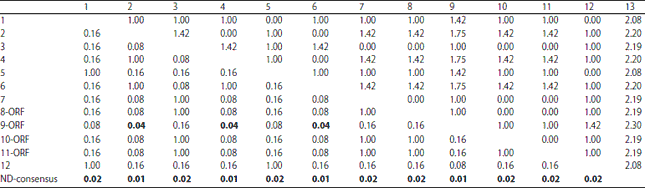 | |
The calculated Ka/Ks ratios (0.517) for the F gene of the 12 examined strains predicted a low evolution rate, with no positive selection sites identified (Table 8).
Accession numbers: Strains are submitted to GenBank through BanKit and we got accession numbers for two nucleotide sequences and the other nucleotide sequences are under processing. The two nucleotide sequences accession numbers are: MK829159 and MK829160.
DISCUSSION
Newcastle disease (ND) is one of the most important viral diseases33 and causes huge economic losses to the commercial poultry farmers worldwide34-36. The recent recurrent of ND outbreaks in vaccinated flocks raises questions about the vaccine efficacy. In Egypt, NDV outbreaks are occurring frequently despite strict vaccination programs and the epidemiology of virulent NDV isolates from these outbreaks is not known37-39.
Virus isolation and haemagglutination inhibition tests were carried out according to the OIE22 report using hyperimmune serum against NDV, which was previously prepared according to the report by Pansota et al.27. The results obtained from the HI test were compatible with those reported by Alders et al.40, Manin et al.41, Peroulis and O’Riley42 and Singh et al.43.
Biological characterization of field isolates was carried out using the intracerebral pathogenicity index (ICPI) in day-old chicks, as ICPI is considered as the most sensitive and widely used test for measuring virulence, according to Terregino and Capua25 and OIE22. Signs that appeared in chicks subjected to the ICPI test from the 3-4th day post intracerebral injection for the most velogenic strain from our isolates were specific nervous signs, including paralysis of one or both legs and wings, twisting of the head and neck, muscular tremors, in coordination in gait and opisthotonos followed by death (Fig. 1). The postmortem examination of chicks injected with the most velogenic strains revealed congestion of the blood vessels of the brain (cerebrum and cerebellum) and haemorrhages in the proventriculus, severe enteritis, congested lungs and congested inflamed kidneys with ureate deposition in both ureters (Fig. 2), which was in agreement with what was reported by Alexander44.
ICPI determination of 15 isolates in day-old chicks resulted in one velogenic isolate (ICPI = 1.6) with a value greater than 1.5, which is typical of velogenic NDV strains, according to Lima et al.45 Carrasco et al.46 and the remaining isolates were mesogenic (ICPIs ranged from 0.85-1.5), with values from 0.5-1.5 (Table 3).
The examined NDV strains contain the F protein cleavage site motif sequence112R-R-Q-R-R-F117 (Fig. 4), which is the major determinant of virulence for NDV strains, according to Rott and Klenk47. This high proportion of basic amino acids and the presence of a phenylalanine at position 117 (F117) are characteristic of highly virulent strains, as previously described in other vNDV strains48-52.
There was a difference in the ICPI results and deduced amino acids of the cleavage site, which can be explained by some drawbacks to ICPI in the interpretation of pathotype results. For example, a previous study by Pearson et al.53 reported 10 NDV isolates from pigeons to have ICPI values between 1.2 and 1.45 and a range of intravenous pathogenicity index (IVPI) values from 0.0-1.3, suggesting that the viruses were virulent; however, the lowest mean death time (MDT) recorded was 98 h which is characteristic of lentogenic viruses. Although, the standard pathogenicity indices can often offer a good estimate of the virulence, they do not always correlate exactly with what is observed in animal experiments, especially when the virus is administered to adult birds via a possible natural route of inoculation54,55.
In other words, the MDT and ICPI do not always correlate with the clinicopathologic syndrome, or the “disease-inducing ability” of each strain. Furthermore, it should be noted that the status of "notifiability", as indicated by the OIE (ICPI ≥0.7 and/or virulent F protein cleavage sequence), covers a broad span of pathogenic potential, encompassing viruses that can cause very severe lesions or none23.
The predicted amino acid sequences of the partial F gene of the 12 examined strains were compared with those from different strains pertaining to classes I and II (genotypes I-XVIII) (Fig. 4). All seven neutralizing epitopes important for both structure and function of the protein were conserved12,56,57.
To study the phylogenetic relationship of the isolated strains with the other global ones, nucleotide and amino acid sequence analyses of the F gene were performed for the 12 isolates and then compared with 121 NDV strains from GenBank representing several NDV genotypes and subgenotypes from various regions of the world at disparate time periods (Fig. 5).
The results of nucleotide sequence analysis revealed that the highest similarity detected between the examined isolates and strains from genotype VIId ranged from 99.3-88.1%, with the greatest nucleotide identity (99.3%) recorded with a velogenic strain from Jordan (JQ176687-Chicken-Jordan-Jo11-2011-VIId (5d). In addition, the examined isolates were highly similar but to a lesser degree than those mentioned previously, to JN979564-Turkey-Israel-111-2011-Israel VIId (5d), FJ608338-SH-Zhejiang-06-VIId, DQ486859-GM-VIId, AY630436-Kr-188-02-VIId, AY630438-ch-501-02-VIId, DQ439937-NDV05-085-VIId and AF358786-TW-2000-VIId, which displayed nucleotide sequence homology from 95.6-88.1%.This high identity proves that the virus circulating in the study area probably extends from the Middle Eastern region and belongs to group VIId, as also reported by Radwan et al.39 NDV genotype VII appeared in the late eighties and became the predominant genotype among NDV infections in chickens34.
In specific comparison with the Egyptian strains, our isolates showed the highest similarity percentages with JX885868-chicken-VRLCU138-Egypt-2012(98.6%), JX173098 -NDV-chicken-Giza-Egypt-MR0-2012(98.2%) and KP209305-NDV-chicken-Egypt-FA-8-2014(95.8%), followed by a lesser degree of homology with FJ969394-chicken-Egypt-3-2006 (84,6%), JX193771-NDV-chicken-Behaira-Egypt-MR6-2012 (83.3%) and JX193772-NDV-chicken-Fayoum-Egypt-MR7-2011 (73.6%).
The homologies of nucleotide sequences were relatively low with the other genotypes, where the similarity with genotype I was 83.7-76.2% and with genotype II was 83.9-74.2%, whereas live virus vaccines used commercially are derived from those genotypes. The homology between our isolates and the strain AF217084-V4 I-Australia-I was 83.1-78.1%, with ay562991-ulster-I was 81.6%- 76.2%, with Y18898-CLONE30-IIwas 82.2-74.8%, with NC_002617-B1-2000-II was 82.2-74.8%, with M24697-BeaudetteC-45-II was 83.1-74.8%, with M24696-Lassato-46-II was 82.2-74.8%, with FJ969395-NDV-chicken-Egypt-4-2006-II was 83.9-76.4%, with FJ386395-NDV04-2004-II was 82.5-75.2%, with AY289002-VGGA-87-II was 81.2-74.8%, with AY170137-Komarov-45-LK-II was 81.9-74.9%, with AF077761-LaSota-II was 82.2-74.8% and with M24695-HB1-47-II was 82.7-75.2%.
This low percentage of homology between the vaccine strains belonging to group II and our isolates may explain the vaccine failure cases despite good biosecurity and vaccination practices in recent years. The same field observations have also been reported from other countries, such as in Miller et al.9.
It has been found that all the pathogenic strains belong to one serotype and that, upon vaccination, vaccine strains from virulent and less virulent strains from genotypes I and II can protect against disease from any genotype but fail to prevent viral shedding58,59. As a result of this continuous infection and shedding, NDVs have opportunities to acquire specific mutations that facilitate evasion of the immune response induced by vaccination60.
A few cross-protection tests indicated that current commercial vaccines (LaSota) do not induce enough immunity against newly emerging genotypes and that there are at least a few strains known as immune-escaping antigenic variants responsible for recent ND outbreaks, as reported by Qin et al.35 and Yu et al.61.
NDV-chicken-Egypt-Asy-11-2015 shared similarities with the Komarov strain that was possibly generated from the vaccine strain either through direct exposure of birds to the live vaccine or to infectious Komarov-like strains circulating in rural poultry. Recently, Rui et al.62 also proposed vaccine strains as N-terminal donors in recombinant NDV from chickens. These observations highlight the relevance that live vaccine strains may play a role in the evolution of ND viruses.
Some evidence, however, suggests that the current commercial vaccine could provide enough protection even in challenge with heterologous strains of different genotypes12. However, it is widely accepted that disease is preventable with the use of current ND vaccines but viral shedding is affected by phylogenic and antigenic distance9,18.
Seven major epitopes have been characterized involving the fusion inhibition and neutralization of F protein located at positions 72, 74, 75, 78, 79 and 157-171, which are significant for both structure and function of the F- gene57,63,64. We identified seven neutralizing epitopes located at positions 72, 74, 75, 78, 79 and 157-171 of the F protein that showed an F for K amino acid substitution at position 78 in all strains. No recombination events were recognized but a point mutation in one of the neutralizing epitopes (F78 for K) of the F protein was characterized in the field isolates.
To localize the NDV strains into the phylogenetic tree and to compare the isolates with other viruses isolated worldwide, one tree was constructed using 121 partial genome nucleotide sequences of representative viruses classified as class I and class II (genotypes I-XIII and XVI) available at GenBank. The results revealed that 10 strains belong to subgenotype VIId, one strain to subgenotype VIIa and another to subgenotype II (Fig. 5). The genotype VIId isolates were the most prevalent in the study site, which was the same result as some previous reports37-39,65,66.
The Ka/Ks ratio ranged from 0.01-0.02, which is a value less than one, indicating that negative or purifying selection is driving the selective removal of alleles that are deleterious. This effect can result in stabilizing selection through the purging of deleterious variations that arise. The calculated Ka/Ks ratios (0.517) for the F gene of the 12 examined strains predicted a low evolution rate, with no positive selection sites were identified (Table 8), indicating that all NDV proteins are under strong purifying and negative selection pressures, as reported by Miller et al.67 and Chong et al.68.
CONCLUSION
All the viruses isolated in this study were defined as velogenic based on the ICPI, which ranged from 0.85-1.6 and the results of fusion protein cleavage site analysis (FPCS) from the sequence of deduced amino acids of the F- gene; all isolated strains have the motif 112R-R-Q-R-R-F117, indicating that these strains were velogenic. Several genotypes were observed, including VIId, VIIa and II and most of the isolates proved to be of genotype VIId. The presence of multiple NDV strains further complicates attempts to prevent the spread of the disease among chickens. The nucleotide sequences of lineage VIId isolates genetically related to Turkey NDV isolates from Israel provide proof for potential epidemiologic linkage between outbreaks of NDV in these countries.
The examined isolates were genetically distant from genotype II (lentogenic vaccine strains used worldwide), at only 74.2-83.9% similarity, suggesting that the NDV vaccine used in this area may not be very efficient in preventing viral shedding, which will allow unobserved circulation of the virulent virus in the vaccinated bird population until evolution of an outbreak. Consequently, it may be a prerequisite to evaluate the effectiveness of the current vaccine used in Egypt against circulating NDV strains. The strain NDV-chicken-Egypt-Asy-11-2015 shared similarities with the Komarov strain, which suggests that this Komarov-like strain could have arisen from a vaccine strain either through direct exposure of chickens to the vaccine by routine vaccination or exposure to infectious Komarov-like viruses that might have been mutated from the vaccine strain and commonly circulate in rural poultry.
FUTURE RESEARCH
Our study concluded that the control of NDV by vaccination still faces new challenges using current commercial vaccines. Therefore, our future research will investigate the possibility of designing an effective vaccine based on our field strains from the predominant genotype VIId. This vaccine is supposed to provide improved protection against genotype-matched field strains and decrease viral shedding.
SIGNIFICANCE STATEMENT
This study reveals that the genotype VIId is the dominant genotype of NDV in Egypt, not genotype II, from which lentogenic vaccine strains originate. This genotype (VIId) is genetically distant from genotype II, with similarities from 74.2-83.9%. This difference can be beneficial for understanding that NDV vaccines used in this area may not be very effective in stopping viral shedding. Therefore, it is necessary to evaluate the effectiveness of the current vaccines used in the Upper Egypt provinces against circulating NDV strains. This study will help researchers to investigate the possibility of designing new vaccines based on circulating field isolates for better protection and minimization of viral shedding.
ACKNOWLEDGMENTS
We thank Professor Mustafa Al-Bakri Seif Al-Din for his contributions in this study.
REFERENCES
- Kapczynski, D.R., C.L. Afonso and P.J. Miller, 2013. Immune responses of poultry to Newcastle disease virus. Dev. Compa. Immunol., 41: 447-453.
CrossRefDirect Link - Ganar, K., M. Das, S. Sinha and S. Kumar, 2014. Newcastle disease virus: Current status and our understanding. Virus Res., 184: 71-81.
CrossRefDirect Link - Lamb, R.A., R.G. Paterson and T.S. Jardetzky, 2006. Paramyxovirus membrane fusion: Lessons from the F and HN atomic structures. Virology, 344: 30-37.
CrossRefDirect Link - Deng, R., Z. Wang, A.M. Mirza and R.M. Iorio, 1995. localization of a domain on the paramyxovirus attachment protein required for the promotion of cellular fusion by its homologous fusion protein spike. Virology, 209: 457-469.
CrossRefDirect Link - Melanson, V.R. and R.M. Iorio, 2004. Amino acid substitutions in the F-specific domain in the stalk of the newcastle disease virus HN protein modulate fusion and interfere with its interaction with the F protein. J. Virol., 78: 13053-13061.
CrossRefDirect Link - Pedersen, J.C., D.A. Senne, P.R. Woolcock, H. Kinde and D.J. King et al., 2004. Phylogenetic relationships among virulent Newcastle disease virus isolates from the 2002-2003 outbreak in California and other recent outbreaks in North America. J. Clin. Microbiol., 42: 2329-2334.
CrossRefDirect Link - Miller, P.J., E.L. Decanini and C.L. Afonso, 2010. Newcastle disease: Evolution of genotypes and the related diagnostic challenges. Infect. Genet Evol., 10: 26-35.
CrossRefDirect Link - Kim, S.H., S. Nayak, A. Paldurai, B. Nayak and A. Samuel et al., 2012. Complete genome sequence of a novel newcastle disease virus strain isolated from a chicken in West Africa. J. Virol., 86: 11394-11395.
CrossRefDirect Link - Diel, D.G., L.H.A. da Silva, H. Liu, Z. Wang, P.J. Miller and C.L. Afonso, 2012. Genetic diversity of Avian paramyxovirus type 1: Proposal for a unified nomenclature and classification system of Newcastle disease virus genotypes. Infect. Genet. Evol., 12: 1770-1779.
CrossRefDirect Link - Liu, X.F., H.Q. Wan, X.X. Ni, Y.T. Wu and W. B. Liu, 2003. Pathotypical and genotypical characterization of strains of Newcastle disease virus isolated from outbreaks in chicken and goose flocks in some regions of China during 1985-2001. Arch. Virol., 148: 1387-1403.
CrossRefDirect Link - Aldous, E.W., C.M. Fuller, J.K. Mynn and D.J. Alexander, 2004. A molecular epidemiological investigation of isolates of the variant avian paramyxovirus type 1 virus (PPMV-1) responsible for the 1978 to present panzootic in pigeons. Avian Pathol., 33: 258-269.
CrossRefDirect Link - Cattoli, G., A. Fusaro, I. Monne, S. Molia and A. Le Menach et al., 2010. Emergence of a new genetic lineage of Newcastle disease virus in West and Central Africa-implications for diagnosis and control. Vet. Microbiol., 142: 168-176.
CrossRefDirect Link - Khan, T.A., C.A. Rue, S.F. Rehmani, A. Ahmed, J.L. Wasilenko, P.J. Miller and C.L. Afonso, 2010. Phylogenetic and biological characterization of Newcastle disease virus isolates from Pakistan. J. Clin. Microbiol., 48: 1892-1894.
CrossRefDirect Link - Lee, Y.J., H.W. Sung, J.G. Choi, E.K. Lee, H. Yoon, J.H. Kim and C.S. Song, 2008. Protection of chickens from Newcastle disease with a recombinant baculovirus subunit vaccine expressing the fusion and hemagglutinin-neuraminidase proteins. J. Vet. Sci., 9: 301-308.
CrossRefDirect Link - Miller, P.J., D.J. King, C.L. Afonso and D.L. Suarez, 2007. Antigenic differences among Newcastle disease virus strains of different genotypes used in vaccine formulation affect viral shedding after a virulent challenge. Vaccine, 25: 7238-7246.
CrossRefDirect Link - Cho, S.H., H.J. Kwon, T.E. Kim, J.H. Kim and H.S. Yoo et al., 2008. Characterization of a recombinant Newcastle disease virus vaccine strain. Clin. Vaccine Immunol., 15: 1572-1579.
CrossRefDirect Link - Hines, N.L. and C.L. Miller, 2012. Avian paramyxovirus serotype-1: A review of disease distribution, clinical symptoms and laboratory diagnostics. Vet. Med. Int., Vol. 2012.
CrossRefDirect Link - Xiao, S., B. Nayak, A. Samuel, A. Paldurai and M. Kanabagattebasavarajappa et al., 2012. Generation by reverse genetics of an effective, stable, live-attenuated newcastle disease virus vaccine based on a currently circulating, highly virulent Indonesian strain. PLoS One, Vol. 7, No. 12.
CrossRefDirect Link - OIE., 2012. Newcastle Disease. In: Manual of Diagnostics Tests and Vaccines for Terrestrial Animals (Mammals, Birds and Bees), OIE., (Eds.)., World Organisation for Animal Health, France, pp: 555-573.
Direct Link - Iqbal, M., K. Mahboob, M. Zulifiqar, A. Haq, G. Nabi, and R. Tubssum, 2003. Production of hyperimmune serum against Newcastle Disease Virus (NDV) in rabbits. Pak. J. Vet. Res., 1: 22-25.
Direct Link - Pansota, F.M., F. Rizvi, A. Sharif, M.T. Javed, G. Muhammad, A. Khan and M.Z. Khan, 2013. Use of hyperimmune serum for passive immunization of chicks experimentally infected with newcastle disease virus. Pak. J. Agric. Sci., 50: 279-288.
Direct Link - Liang, R., D.J. Cao, J.Q. Li, J. Chen, X. Guo, F.F. Zhuang and M.X. Duan, 2002. Newcastle disease outbreaks in western China were caused by the genotypes VIIa and VIII. Vet. Microbiol., 87: 193-203.
CrossRefPubMedDirect Link - Berhanu, A., A. Ideris, A.R Omar and M.H. Bejo, 2010. Molecular characterization of partial fusion gene and C-terminus extension length of haemagglutinin-neuraminidase gene of recently isolated Newcastle disease virus isolates in Malaysia. Virol. J., Vol. 7, No. 183.
CrossRefDirect Link - Tamura, K., J. Dudley, M. Nei and S. Kumar, 2007. MEGA4: Molecular Evolutionary Genetics Analysis (MEGA) software version 4.0. Mol. Biol. Evol., 24: 1596-1599.
CrossRefPubMedDirect Link - Orsi, M.A., L. Doretto Junior, S.C.A. Camillo, D. Reischak and S.A.M. Ribeiro et al., 2010. Prevalence of Newcastle disease virus in broiler chickens (Gallus gallus) in Brazil. Braz. J. Microbiol., 41: 349-357.
CrossRefDirect Link - Aldous, E.W., J.K. Mynn, J. Banks and D.J. Alexander, 2003. A molecular epidemiological study of avian paramyxovirus type 1 (Newcastle disease virus) isolates by phylogenetic analysis of a partial nucleotide sequence of the fusion protein gene. Avian Pathol., 32: 239-257.
CrossRefDirect Link - Qin, Z.M., L.T. Tan, H.Y. Xu, B.C. Ma, Y.L. Wang, X.Y. Yuan and W.J. Liu, 2008. Pathotypical characterization and molecular epidemiology of Newcastle disease virus isolates from different hosts in China from 1996 to 2005. J. Clin. Microbiol., 46: 601-611.
CrossRefDirect Link - Diel, D.G., L. Susta, S.C. Garcia, M.L. Killian, C.C. Brown, P.J. Miller and C.L. Afonso, 2012. Complete genome and clinicopathological characterization of a virulent Newcastle disease virus isolate from South America. J. Clin. Microbiol., 50: 378-387.
CrossRefDirect Link - Hussein, H.A., M.M. Emara and M.A. Rohaim, 2014. Molecular characterization of Newcastle disease virus genotype VIID in Avian influenza H5N1 infected broiler flock in Egypt. Int. J. Virol., 10: 46-54.
CrossRefDirect Link - Radwan, M.M., S.F. Darwish, I.M. El-Sabagh, A.A. El-Sanousi and M.A. Shalaby, 2013. Isolation and molecular characterization of Newcastle disease virus genotypes II and VIId in Egypt between 2011 and 2012. Virus Genes, 47: 311-316.
CrossRefDirect Link - Alders, R.G., S. Inoue and J.C. Katongo, 1994. Prevalence and evaluation of hitchner B1 and V4 vaccines for the control of newcastle disease in village chickens in Zambia. Preventive Vet. Med., 21: 125-132.
CrossRefDirect Link - Manin, T.B., L.O. Shcherbakova, I.A. Bochkov, V.V. El'nikov, I.P. Pchelkina, S.K. Starov and V.V. Drygin, 2002. Characteristics of field isolates of Newcastle disease virus isolated in the course of outbreaks in the poultry plant in the Leningrad region in 2000. Vopr. Virusol., 47: 41-43, (In Russian).
PubMedDirect Link - Peroulis, I. and K. O'Riley, 2004. Detection of avian paramyxoviruses and influenza viruses amongst wild bird populations in Victoria. Aust. Vet. J., 82: 79-82.
CrossRefDirect Link - Singh, K., N. Jindal, S.L. Gupta, A.K. Gupta and D. Mittal, 2005. Detection of newcastle disease virus genome from the field outbreaks in poultry by reverse transcription-polymerase chain reaction. Int. J. Poult. Sci., 4: 472-475.
CrossRefDirect Link - Lima, F.S., E. Santin, A.C. Paulillo, L. Doretto Junior, V.M.B. de Moraes, N.M.Q. Gama and R.P. Schocken-Iturrino, 2004. Evaluation of different programs of Newcastle disease vaccination in Japanese quail (Coturnix coturnix japonica). Int. J. Poult. Sci., 3: 354-356.
CrossRefDirect Link - Carrasco, A.O.T., M.C. Seki, T.F. Raso, A.C. Paulillo and A.A. Pinto, 2008. Experimental infection of Newcastle disease virus in pigeon (Columbia livia): Humoral antibody response, contact transmission and viral genome shedding. Vet. Microbiol., 129: 89-96.
PubMed - Li, Z., T. Sergel, E. Razvi and T. Morrison, 1998. Effect of cleavage mutants on syncytium formation directed by the wild-type fusion protein of Newcastle disease virus. J. Virol., 72: 3789-3795.
PubMedDirect Link - Peeters, B.P., O.S. de Leeuw, G. Koch and A.L. Gielkens, 1999. Rescue of Newcastle disease virus from cloned cDNA: Evidence that cleavability of the fusion protein is a major determinant for virulence. J. Virol., 73: 5001-5009.
Direct Link - Panda, A., S. Elankumaran, S. Krishnamurthy, Z. Huang and S.K. Samal, 2004. Loss of N-linked glycosylation from the hemagglutinin- neuraminidase protein alters virulence of newcastle disease virus. J. Virol., 78: 4965-4975.
CrossRefDirect Link - Kattenbelt, J.A., J. Meers and A.R. Gould, 2006. Genome sequence of the thermostable Newcastle disease virus (strain I-2) reveals a possible phenotypic locus. Vet. Microbiol., 114: 134-141.
CrossRefDirect Link - Pearson, J.E., D.A. Senne, D.J. Alexander, W.D. Taylor, L.A. Peterson and P.H. Russel, 1987. Characterization of newcastle disease virus (Avian paramyxovirus-1) isolated from pigeons. Avian Dis., 31: 105-111.
PubMedDirect Link - Wakamatsu, N., D.J. King, B.S. Seal, S.K. Samal and C.C. Brown, 2006. The pathogenesis of Newcastle disease: A comparison of selected Newcastle disease virus wild-type strains and their infectious clones. Virology, 353: 333-343.
CrossRefDirect Link - Susta, L., P.J. Miller, C.L. Afonso and C.C. Brown, 2011. Clinicopathological characterization in poultry of three strains of Newcastle disease virus isolated from recent outbreaks. Vet. Pathol., 48: 349-360.
CrossRefDirect Link - Yusoff, K., M. Nesbit, H. McCartney, G. Meulemans and D.J. Alexander et al., 1989. Location of neutralizing epitopes on the fusion protein of Newcastle disease virus strain Beaudette C. J. Gen. Virol. 70: 3105-3109.
CrossRefDirect Link - Kapczynski, D.R. and D.J. King, 2005. Protection of chickens against overt clinical disease and determination of viral shedding following vaccination with commercially available Newcastle disease virus vaccines upon challenge with highly virulent virus from the California 2002 exotic Newcastle disease outbreak. Vaccine, 23: 3424-3433.
CrossRefDirect Link - Miller, P.J., E.L. Decanini and C.L. Afonso, 2010. Newcastle disease: Evolution of genotypes and the related diagnostic challenges. Infect. Genet Evol., 10: 26-35.
CrossRefDirect Link - Perozo, F., R. Marcano and C.L. Afonso, 2012. Biological and phylogenetic characterization of a genotype vii newcastle disease virus from Venezuela: Efficacy of field vaccination. J. Clin. Microbiol., 50: 1204-1208.
CrossRefDirect Link - Yu, L., Z. Wang, Y. Jiang, L. Chang and J. Kwank, 2001. Characterization of newly emerging Newcastle disease virus isolates from the People's Republic of China and Taiwan. J. Clin. Microbiol., 39: 3512-3519.
CrossRefPubMedDirect Link - Rui, Z., P. Juan, S. Jingliang, Z. Jixun and W. Xiaoting et al., 2010. Phylogenetic characterization of Newcastle disease virus isolated in the mainland of China during 2001-2009. Vet. Microbiol., 141: 246-257.
CrossRefDirect Link - Toyoda, T., B. Gotoh, T. Sakaguchi, H. Kida and Y. Nagai, 1988. Identification of amino acids relevant to three antigenic determinants on the fusion protein of newcastle disease virus that are involved in fusion inhibition and neutralization. Am. Soc. Microbiol., 62: 4427-4430.
PubMedDirect Link - Neyt, C., J. Geliebter, M. Slaoui, D. Morales, G. Meulemans and A. Burny, 1989. Mutations located on both F1 and F2 subunits of the Newcastle disease virus fusion protein confer resistance to neutralization with monoclonal antibodies. J. Virol., 63: 952-954.
Direct Link - Mohamed, M.H., S. Kumar, A. Paldurai, S.K. Samal, 2011. Sequence analysis of fusion protein gene of newcastle disease virus isolated from outbreaks in Egypt during 2006. Virol. J., 8: 1-237.
Direct Link - Nabila, O., S. Sultan, A.I. Ahmed, R.S. Ibrahim and M. Sabra, 2014. Isolation and pathotyping of newcastle disease viruses from field outbreaks among chickens in the Southern part of Egypt 2011-2012. Global Vet., 12: 237-243.
Direct Link - Miller, P.J., L.M. Kim, H.S. Ip and C.L. Afonso, 2009. Evolutionary dynamics of Newcastle disease virus. Virology, 391: 64-72.
CrossRefDirect Link - Chong, Y.L., A. Padhi, P.J. Hudson and M. Poss, 2010. The effect of vaccination on the evolution and population dynamics of avian paramyxovirus-1. PLoS Pathog., Vol. 6, No. 4.
CrossRefDirect Link