
Xiang-Yu Long
Triticeae Research Institute, Sichuan Agricultural University, Wenjiang, Sichuan, 611130, China
Ya-Xi Liu
Triticeae Research Institute, Sichuan Agricultural University, Wenjiang, Sichuan, 611130, China
Helene Rocheleau
Eastern Cereal and Oilseed Research Centre, Agriculture and Agri-Food Canada, 960 Carling Ave, Ottawa, On, K1A 0C6, Canada
Therese Ouellet
Eastern Cereal and Oilseed Research Centre, Agriculture and Agri-Food Canada, 960 Carling Ave, Ottawa, On, K1A 0C6, Canada
Guo-Yue Chen
Triticeae Research Institute, Sichuan Agricultural University, Wenjiang, Sichuan, 611130, China
International Journal of Plant Breeding and Genetics
Year: 2011 | Volume: 5 | Issue: 3 | Page No.: 255-267







ABSTRACT
Using Quantitative real-time PCR to quantify gene expression, it is vital that one or more ideal internal control genes are used to normalize the samples to be compared. 10 traditional internal control genes were selected to evaluate the stability of expression in wheat leaves infected by strip rust, by qRT-PCR, including 18S Ribosomal RNA (18S), 26S Ribosomal RNA (26S), 60S Ribosomal protein L18a-1(60S), Actin, Cyclophilin (Cyp), Elongation factor 1 alpha (EF1A), Glyceraldehyde-3-phosphate dehydrogenase (GAPDH), Histone H3 (Histone),α-Tubulin and β-Tubulin. Expression stability of internal control genes was statistically analyzed by three different mathematical algorithms, GeNorm, NormFinder and BestKeeper. Three softwares gave similar results that 60S, Cyp and EF1A were the most stable whereas 18S and 26S did not favour normalization of qRT-PCR results in those samples. Cyp and EF1A have medium RNA transcription levels, and 18S and 26S have high RNA transcription levels. Using different normalization factors, expression levels of the target gene were obviously different. This study indicated that selection of the most stable internal control genes play an important role in gene expression and 60S, Cyp and EF1A would be ideal for normalization to target gene expression in wheat leaves infected by strip rust.
PDF Abstract XML References Citation
Received: November 08, 2010;
Accepted: March 20, 2011;
Published: May 21, 2011
How to cite this article
Xiang-Yu Long, Ya-Xi Liu, Helene Rocheleau, Therese Ouellet and Guo-Yue Chen, 2011. Identification and Validation of Internal Control Genes for Gene Expression in Wheat Leaves Infected by Strip Rust. International Journal of Plant Breeding and Genetics, 5: 255-267.
DOI: 10.3923/ijpbg.2011.255.267
URL: https://scialert.net/abstract/?doi=ijpbg.2011.255.267
DOI: 10.3923/ijpbg.2011.255.267
URL: https://scialert.net/abstract/?doi=ijpbg.2011.255.267
INTRODUCTION
Among the various methods available for generating gene expression profiling data, quantitative real-time polymerase chain reaction (qRT-PCR) is one of the most widely used technique and plays an important role in current biological research (Baarsch et al., 1995; Skovgaard et al., 2006; Kavousi et al., 2009; Cossio-Bayugar et al., 2008). The advantages of qRT-PCR include technical ease, low reagents cost, less hands-on time and high throughput, therefore making it a more efficient and reliable techniques (Wang and Brown, 1999; Bustin, 2000; Garcia-Vallejo et al., 2004). However, there are several parameters that need to be controlled during qRT-PCR, including initial sample amount, RNA recovery, RNA integrity, efficiency of cDNA synthesis and differences in the overall transcriptional activity of the tissues or cells analyzed (Andersen et al., 2004; Huggett et al., 2005; Chen et al., 2006). To obtain accurate results, the expression level of the targeted genes must be normalized using internal control genes, known as reference genes or as housekeeping genes. These internal control genes should be consistently expressed in different tissues or under different treatments (Thellin et al., 1999; Daud and Scott, 2008; Frericks and Esser, 2008; Mekki et al., 2006). Over the last few decades, the genes β-actin, glyceraldehyde-3-phosphate dehydrogenase (GAPDH), 18S or 26S ribosomal RNA (18S or 26S rRNA), ubiquitin C (UBC), α-tubulin (TUB), elongation factor-1 alpha and TATA-Box binding protein (TBP) had been validated as suitable internal control genes in several species for qRT-PCR (Bustin, 2000; Goidin et al., 2001; Kim and Kim, 2003; Lossos et al., 2003; Mitter et al., 2009).
Unfortunately, it is impossible to find an ideal and universal internal control gene for normalization in various samples (Thellin et al., 1999; Warrington et al., 2000; Bustin, 2002; Glare et al., 2002; Bas et al., 2004). This is because the expression level of an internal control gene might only be consistent in certain tissues but not others. For instance, GAPDH acted as a reliable reference gene in non-small cell lung cancer, whereas the hypoxanthine phosphoribosyl-transferase (HPRT) was consistently in aging rat liver (0 2002; Liu et al., 2005; Chen et al., 2006). The suitability of internal control genes for normalization are calculated and ranked by algorithms softwares such as geNorm, NormFinder, Bestkeeper and General Pattern Recognition (Vandesompele et al., 2002; Gabrielsson et al., 2005; Akilesh et al., 2003; Pfaffl et al., 2004; Andersen et al., 2004). The highest ranked internal control gene is then selected before doing qRT-PCR.Wheat strip rust, also known as yellow rust, caused by Puccinia striiformis westend. f.sp.tritici Eriks., is one of the most destructive disease of wheat worldwide (Chen, 2005). It infects wheat in early its stage of development, leading to reduce yield and quality of wheat (Roelfs et al., 1992). Some of rust resistance genes had been identified and characterized using differential display reverse transcriptase-PCR method (DDR), Suppression Subtractive Hybridization (SSH) and micro-array. These genes included Lr10, Lr21, Yr10 and other pathogenesis-related (PR) proteins (Wang et al., 2009; Hulbert et al., 2007; Coram et al., 2009; Feuillet et al., 2003; Huang et al., 2003). To understand the resistance mechanism of wheat to strip rust, it is necessary to study gene expression pattern to analysis the salvage pathway. In this study, 10 traditional internal control genes were selected (Table 1) and tested for expression stability in wheat when the leaves were infected by strip rust using geNorm, NormFinder and Bestkeeper software to analyze the data of expression, these 10 internal control genes were ranked from the most transcriptionally stable gene to the least transcriptionally stable gene.
MATERIALS AND METHODS
Plant materials and growth conditions: The hexaploid Chinese Spring (Triticum aestivunm L.), which is highly susceptible to strip rust, was used in this study. The seeds were germinated between two sheets of blotting paper and immersed in distilled water. When the roots of the seedlings were about 10 cm long, the seedlings were transferred to tanks with soil in the greenhouse. During the trefoil stage, the plants were then inoculated with CY32, the prevalent race of strip rust was supplied by Gansu Academy of Agricultural Sciences. After inoculation, the plants were kept at high humidity in the dark for 24 h and followed by a regular day-night cycle in a growth chamber. A control inoculation was made with sterile water. Wheat leaves were excised at 0, 12, 24, 36, 48, 72 and 96 h post-inoculation (hpi) and quickly frozen in liquid nitrogen and stored at -80°C prior to RNA extraction.
Primer design: Ten candidate reference genes were selected from the literature for qRT-PCR: Histone H3, 60S ribosomal protein L18a, Actin, Beta-tubulin, Alpha-tubulin, 18S rRNA, Cyclophilin, Elongation factor 1-alpha, 26S rRNA and GAPDH (Table 1). The primers of target gene(pathogenesis related proteins) were designed by Primer 5.0 (Premier Biosoft International, Palo Alto, CA) and DNAman 5.0 (Lynnon BioSoft, USA) according to manufacturer’s instructions.
| Table 1: | Details of traditional internal control genes |
 | |
RNA extraction and cDNA synthesis: After grinding of the samples in liquid nitrogen using mortar and pestle, the total RNA was isolated with Trizol reagent (TaKaRa Biotechnology, Dalian, China) according to the manufacturer’s instructions. The quality of total RNA was assessed by OD260/OD280 ratios and by electrophoresis on 1% agarose gels containing formaldehyde followed by ethidium bromide staining, and then quantified using NanoDrop® ND-1000 Spectrophotometer (Sitthiwong et al., 2007; Matsui et al., 2004). The RNA samples were stored at -80°C.
First-strand cDNA synthesis was performed using PrimeScript® RT reagent Kit (Perfect Real Time) (TaKaRa Biotechnology, Dalian, China) according to the manufacturer’s instructions. The obtained cDNA was diluted 20 fold and stored at -20°C prior to further use as qRT-PCR templates.
Real-time quantitative PCR: All the samples were analysed by qRT-PCR using MyIQ (Bio-Rad) and RealMasterMix (SYBR Green). The procedures began with template denaturation and hot start Taq activation at 95°C for 1 min, then 40 cycles of 95°C for 5 sec, 50-60°C for 30 sec and 72°C for 10 sec and the data of fluorescence was measured after the 72°C extension step. To confirm specificity of the products, the melting step was performed with slow heating, starting at 55°C and with a rate of 0.5°C per 10 sec, up to 95°C, with continuous measurement of fluorescence. The PCR efficiencies (E) and correlation coefficients (R2) for each primer pair were derived from a standard curve consisting of 5 samples each ten fold diluted. All reactions for each gene were run together with duplicate to reduce confounding variance; this included 15 template samples, standard samples and non-template control samples. Threshold cycle (Ct) values were calculated as the cycle when the fluorescence of the sample reached a threshold level called the baseline fluorescence. The mean Ct value was converted to a relative copy number value using the standard curve method.
Statistical analyses: Analysis of gene expression stability, three publicly available softwares GeNorm, Normfinder and Bestkeeper were used to rank the expression stability of internal control genes in all samples (Vandesompele et al., 2002; Gabrielsson et al., 2005; Akilesh et al., 2003; Pfaffl et al., 2004; Andersen et al., 2004). Analysis of mean Ct and pearson correlation coefficient were calculated by Minitab 15 software (http://www.minitab.com/). The expression data of PR were analyzed by BioRad iQ5 Optical System software version 2.0 and shown as the mean of three determinations for each samples.
RESULTS
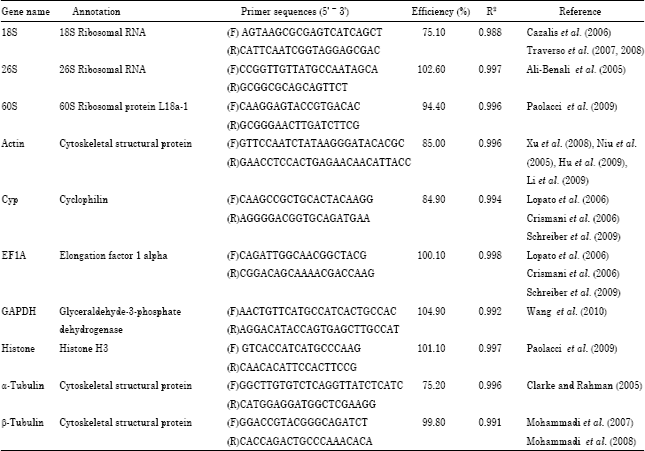
Variations of reference genes: To evaluate the expression levels of internal control genes, the RNA transcription levels of all the samples were measured based on the same baseline in qPCR. The raw expression levels of the ten internal control genes were shown in Fig. 1. Transcripts fell into several groups based on their expression strength, which was classified as: group I with high RNA transcription levels (mean Ct < 21.43), group II with medium RNA transcription levels (21.43 < mean Ct < 25.73) and group III with low RNA transcription levels (mean Ct > 25.73) (Fig. 1). 18S, 28S and GAPDH had the highest expression level with a low Ct value in group I. Cyp and EF1A both had a moderate expression level in group II. The other internal control genes belonged to the lowest expression level with a high Ct value in group III.
Stability of internal control gene within sample groups: To evaluate the stability of expression of ten internal control genes, the measured Ct values were converted to relative copy numbers which were used in geNorm, NormFinder and BestKeeper softwares to calculate expression stability and rank the candidate genes accordingly.
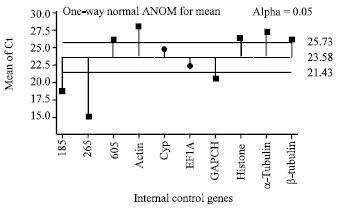
I-geNorm: The program geNorm calculated expression stability (M) based on the average pair-wise variation among all internal control genes (Vandesompele et al., 2002). All of the internal control genes showed high expression stability with M values of less than 1.3, which were below the default limit of 1.5 in the geNorm program (Fig. 2).
 | |
| Fig. 1: | Expression levels of internal control genes |
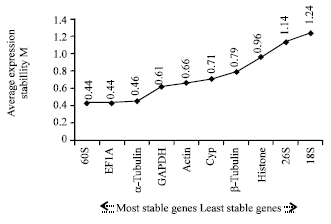
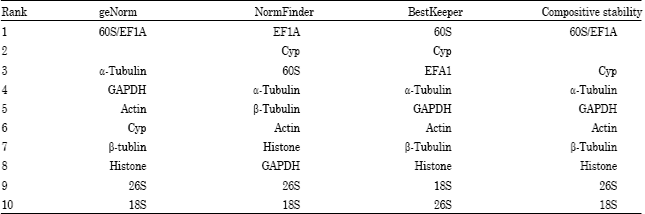
The ten internal control genes were ranked according to the stability measure M, from the most stable (lowest M value) to the least stable (highest M): 60S/EF1A, a-Tubulin, GAPDH, Actin, Cyp, β-Tubulin, Histone, 26S, 18S. This rank order data analysis indicated that 60S and EF1A were the most stable internal control genes, making them ideal for our quantitative analyses. Conversely, the 26S and 18S were the least stable internal control genes in our experiment.
II-NormFinder: NormFinder program was used to calculate an arbitrary stability value and standard error, based on the intragroup and intergroup variations of each gene in all samples (Andersen et al., 2004). The results of the NormFinder analysis were shown in Fig. 3. The EF1A was found to be the most stable (lowest stability value) internal control gene with a stability value of 0.21(0.206), following by Cyp with a stability value of 0.21 (0.214). The two least stable expressed internal control genes were 26S and 18S.
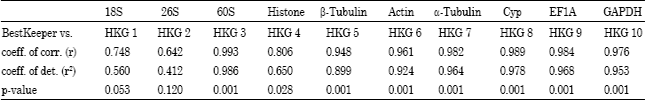
III-BestKeeper: BestKeeper calculated a Pearson correlation coefficient for each internal control gene pair and all highly correlated (and putatively stably expressed) internal control genes were then combined into an index value by calculating the geometric mean. If they were stably expressed internal control genes, their expression levels would be highly correlated (Pfaffl et al., 2004).
 | |
| Fig. 2: | Expression stability of internal control genes calculated by geNorm |
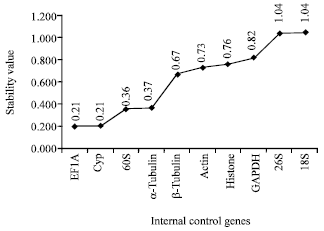 | |
| Fig. 3: | Expression stability of internal control genes calculated by NormFinder |
| Table 2: | Results from bestKeeper correlation analysis |
 | |
| Table 3: | Internal control genes ranking according to geNorm, NormFinder and BestKeeper |
 | |
The ten internal control genes were tested in our analysis of correlate one with another one and the results were shown in Table 2. With the highest correlation between the internal control genes, 60S (r = 0.993) was the most sable internal control gene. The second most internal control gene was Cyp (r = 0.989). The 18S and 26S had the lowest correlation coefficients with a R value were 0.748 and 0.642, respectively, making them the least stable internal control genes.
In this study, all three algorithmic methods gerNorm, NormFinder and BestKeeper have demonstrated that 60S and EF1A are the two most stable internal control genes, with Cyp being the third most stable internal control gene. On the other hand, 18S and 26S were both ranked as the two least reliable internal control genes, as shown in Table 3.
Expression levels of target gene influenced by the choice of internal control gene (s): The geNorm application evaluated the optimal number of internal control genes by using the Normalization Factor (NF) which was calculated by the stepwise inclusion of the following best-ranked gene. Afterwards, pair-wise variations (Vn/n +1) between two subsequent normalization factors were calculated, indicating the effect of including one additional gene for normalization (Vandesompele et al., 2002). It was showed that the V2/3 value was lower than the recommended cut-off value of 0.15. Thus, the geNorm suggested that the 60S and EF1A could be considered as internal control genes for normalization of all samples to analyze the expression of the target gene (Fig. 4). NormFinder calculated the accumulated standard deviation to select the best candidate internal control genes for normalization; the minimal value of the accumulated standard deviation is a great indicator of the optimal number of internal control genes. When the 2 internal control genes that showed greater stability were used for normalization, the accumulated standard deviation had a minimum value (Fig. 5) (Andersen et al., 2004). Therefore, the EF1A and Cyp were selected to normalize the samples to analyze target gene. The 60S and Cyp were the most stable internal control genes using BestKeeper and also selected to normalize the samples to analyze target gene.
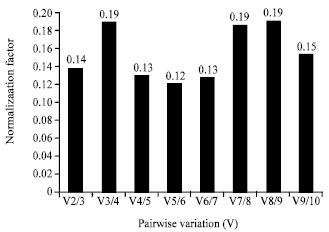 | |
| Fig. 4: | Determination of the optimal number of internal control genes for normalization by geNorm |
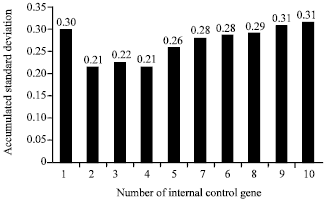 | |
| Fig. 5: | Determination of the optimal number of internal control genes for normalization by NormFinder |
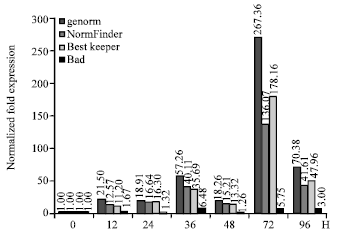 | |
| Fig. 6: | The target gene expression pattern using different normalization factors (NF) from 60S/EF1A, EF1A/Cyp, 60S/Cyp and 18S/26S |
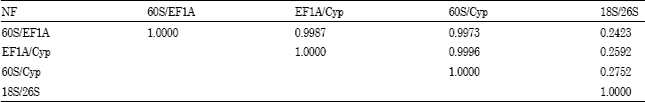
The normalization factors (NF) were calculated for the most stable internal control genes (60S/EF1A, EF1A/Cyp and 60S/Cyp) and least internal control genes (26S/18S). The normalization factors from 60S/EF1A, EF1A/Cyp and 60S/Cyp presented the relatively high correlation, especially between EF1A/Cyp and 60S/Cyp (Table 4). By contrast, the normalization factors from 26S/18S had the extremely low correlation coefficient with other three normalization factors (Table 4).
| Table 4: | Correlation of normalization factors (NF) between 60S/EF1A, EF1A/Cyp, 60S/Cyp and 18S/26S |
 | |
Finally, using the four different normalization factors as normalizers, these differences between the PR10 expression level after normalizing were shown in Fig. 6. The expression level of PR10 had obvious increase after inoculation using the normalization factors from the most stable internal control genes, especially in 72 h inoculation. However, using the least internal control genes to normalization, the expression level of PR10 slowly grown after inoculation and the highest expression level at 36 h inoculated not 72 h. The different normalization factors leaded to totally different result of the expression level.
DISCUSSION
Wheat strip-rust disease is a severe problem in agriculture worldwide and is also one of the leading causes for wheat yield losses in China (Wang et al., 2009). The disease-resistant mechanism is a highly complex phenomenon controlled and regulated by multiple intracellular signalling mechanisms (Bozkurt et al., 2007; Xu et al., 2010). In order to understand strip-rust disease-resistant mechanism, more and more researchers are focusing on analysis and applications of gene expression and regulation. Microarray and qRT-PCR are increasingly becoming the method of choice for high-throughput gene expression analysis (Hulbert et al., 2007; Coram et al., 2009). However, to obtain reliable results from both methods of analysis, accurate normalization of gene expression against one or more internal control genes is vital.
Many algorithmic softwares are employed to evaluate and select the best internal control genes. In this study, the expression data from the 10 internal control genes was analyzed to evaluate their expression stability by three algorithmic softwares: geNorm, NormFinder and BestKeeper. While geNorm and BestKeeper are two of the most popular softwares used for evaluating expression stability, they do not take into account the potential co-regulation of gene expression among the 10 internal control genes. For example, if two of the internal control genes have similar function and participate in the same pathway, then their expression levels would be dependent upon each other (Andersen et al., 2004). Thus, the validity of data generated by these two softwares could be not as reliable. NormFinder, however, is able to control the influence of co-regulation of internal control genes. It calculates the expression stability with intra-group and inter-group variation (Vandesompele et al., 2002). The three softwares mentioned above came to similar conclusion, including the most and least stable internal control genes.
In this study, 60S, EF1A and Cyp were the most appropriate internal control genes for normalizing the samples of wheat strip-rust. The three traditional internal control genes were EF1A, 60S, and Cyp, which present relative stability of expression (Gabrielsson et al., 2005; Silberberg et al., 2009). Although 18S rRNA and 26S were commonly used as internal control genes, this study has demonstrated that they are not suitable due to their instability (Tricarico et al., 2002). Some researchers reported that the 18S rRNA and 26S rRNA were not ideal as internal control genes, because both rRNA were highly compared to mRNA, therefore, their expression level are more easily fluctuated with various biological and drug factors (Mourelatos et al., 2001).
The geNorm and NormFinder also suggested that two internal control genes should be normalized for the samples of wheat strip-rust. Using different normalization factors to normalize, the results indicate that the expression of target gene can be influenced and produce different levels of gene expression. Selection of poor internal control genes could invalidate the normalization process and lead to the generation of misleading information, especially using only one unevaluated internal control gene for normalization (Glare et al., 2002; Haller et al., 2004). Therefore, it is absolutely necessary to establish expression stability of internal control gene for experimental samples before any reliable data could be obtained. To avoid this problem and get the accurate quantitative results, it is recommended that normalization should be carried out with more than one internal control genes that are not functionally related (Dheda et al., 2004; Neuvians et al., 2005; Khimani et al., 2005).
In conclusion, 10 traditional internal control genes were selected to analyse expression stability in samples which were inoculated by wheat strip-rust at different times. Among these internal control genes, 60S, EF1A and Cyp showed high expression stability in the tested samples. On the contrary, the frequently used 18S and 26S were unsuitable for quantifying target gene expression in the tested samples. geNorm, NormFinder and BestKeeper were suitable to perform an adequate normalization for the samples in experiments of gene expression.
ACKNOWLEDGMENT
This study was supported by the National Basic Research Program of China (973 Program 2010CB1344400 and 2009CB118304) and China Transgenic Research Program (2011ZX08002).
REFERENCES
- Akilesh, S., D.J. Shaffer and D. Roopenian, 2003. Customized molecular phenotyping by quantitative gene expression and pattern recognition analysis. Genome Res., 13: 1719-1727.
CrossRefDirect Link - Ali-Benali, M.A., R. Alary, P. Joudrier and M.F. Gautier, 2005. Comparative expression of five Lea genes during wheat seed development and in response to abiotic stresses by real-time quantitative RT-PCR. Biochim. Biophys. Acta Gene Struct. Expression, 1730: 56-65.
CrossRef - Andersen, C.L., J.L. Jensen and T.F. Orntoft, 2004. Normalization of real-time quantitative reverse transcription-PCR data: A model-based variance estimation approach to identify genes suited for normalization, applied to bladder and colon cancer data sets. Cancer Res. 64: 5245-5250.
CrossRefDirect Link - Baarsch, M.J., R.W. Scamurra, K. Burger, D.L. Foss, S.K. Maheswaran and M.P. Murtaugh, 1995. Inflammatory cytokine expression in swine experimentally infected with Actinobacillus pleuropneumoniae. Infect. Immunity, 63: 3587-3594.
PubMedDirect Link - Bas, A., G. Forsberg, S. Hammarstrom and M.L. Hammarstrom, 2004. Utility of the housekeeping genes 18S rRNA, β-Actin and Glyceraldehyde-3-Phosphate-Dehydrogenase for normalization in real-time quantitative reverse transcriptase-polymerase chain reaction analysis of gene expression in human T lymphocytes. Scand. J. Immunol., 59: 566-573.
CrossRefDirect Link - Bozkurt, O., T. Unver and M. Akkaya, 2007. Genes associated with resistance to wheat yellow rust disease identified by differential display analysis. Physiol. Mol. Plant Pathol., 71: 251-259.
CrossRef - Bustin, S.A., 2000. Absolute quantification of mRNA using real-time reverse transcription polymerase chain reaction assays. J. Mol. Endocrinol., 25: 169-193.
CrossRefPubMedDirect Link - Bustin, S.A., 2002. Quantification of mRNA using real-time reverse transcription PCR (RT-PCR): Trends and problems. J. Mol. Endocrinol., 29: 23-39.
PubMed - Cazalis, R., P. Pulido, T. Aussenac, J.M. Perez-Ruiz and F.J. Cejudo, 2006. Cloning and characterization of three thioredoxin h isoforms from wheat showing differential expression in seeds. J. Exp. Bot., 57: 2165-2172.
PubMedDirect Link - Cossio-Bayugar, R., E. Miranda-Miranda, A. Ortiz-Najera, S. Neri-Orantes and F. Olvera-Valencia, 2008. Cytochrome P-450 monooxygenase gene expression supports a multifactorial origin for acaricide resistance in Ripicephalus microplus. Res. J. Parasitol., 3: 59-66.
CrossRefDirect Link - Chen, J., D.A. Rider and R. Ruan, 2006. Identification of valid housekeeping genes and antioxidant enzyme gene expression change in the aging rat liver. J. Gerontol. A Biol. Sci. Med. Sci., 61: 20-27.
PubMedDirect Link - Chen, X.M., 2005. Epidemiology and control of stripe rust [Puccinia striiformis f. sp. tritici] on wheat. Can. J. Plant Pathol., 27: 314-337.
Direct Link - Clarke, B. and S. Rahman, 2005. A microarray analysis of wheat grain hardness. Theor. Applied Genet., 110: 1259-1267.
PubMedDirect Link - Coram, T.E., M.L. Settles and X. Chen, 2009. Large-scale analysis of antisense transcription in wheat using the Affymetrix GeneChip wheat genome array. BMC Genomics, 10: 253-253.
PubMedDirect Link - Crismani, W., U. Baumann, T. Sutton, N. Shirley and T. Webster et al., 2006. Microarray expression analysis of meiosis and microsporogenesis in hexaploid bread wheat. BMC Genomics, 7: 267-267.
CrossRefDirect Link - Daud, I.I. and M.E. Scott, 2008. Validation of reference genes in cervical cell samples from human papillomavirus-infected and uninfected women for quantitative reverse transcription-PCR assays. Clin. Vaccine Immunol., 15: 1369-1373.
PubMedDirect Link - Dheda, K., J.F. Huggett, S.A. Bustin, M.A. Johnson, G. Rook and A. Zumla, 2004. Validation of housekeeping genes for normalizing RNA expression in real-time PCR. Biotechniques, 37: 112-119.
Direct Link - Feuillet, C., S. Travella, N. Stein, L. Albar, A. Nublat and B. Keller, 2003. Map-based isolation of the leaf rust disease resistance gene Lr10 from the hexaploid wheat (Triticum aestivum L.) genome. Proc. Natl. Acad. Sci. USA., 100: 15253-15258.
Direct Link - Frericks, M. and C. Esser, 2008. A toolbox of novel murine house-keeping genes identified by meta-analysis of large scale gene expression profiles. Biochim. Biophys. Acta Gene Regul. Mech., 1779: 830-837.
CrossRef - Gabrielsson, B.G., L.E. Olofsson, A. Sjogren, M. Jernas and A. Elander et al., 2005. Evaluation of reference genes for studies of gene expression in human adipose tissue. Obesity Res., 13: 649-652.
CrossRefDirect Link - Garcia-Vallejo, J.J., B. van het Hof, J. Robben, J.A. van Wijk, I. van Die, D.H. Joziasse and W. van Dijk, 2004. Approach for defining endogenous reference genes in gene expression experiments. Anal. Biochem., 329: 293-299.
PubMedDirect Link - Glare, E., M. Divjak, M. Bailey and E.H. Walters, 2002. β-actin and GAPDH housekeeping gene expression in asthmatic airways is variable and not suitable for normalizing mRNA levels. Thorax, 57: 765-770.
CrossRefDirect Link - Goidin, D., A. Mamessier, M.J. Staquet, D. Schmitt and O. Berthier-Vergnes, 2001. Ribosomal 18s RNA prevails over glyceraldehyde-3-phosphate dehydrogenase and β-actin genes as internal standard for quantitative comparison of mRAN levels in invasive and noninvasive human melanoma cell subpopulations. Anal. Biochem., 295: 17-21.
CrossRef - Haller, F., B. Kulle, S. Schwager, B. Gunawan, A. von Heydebreck, H. Sultmann and L. Fuzesi, 2004. Equivalence test in quantitative reverse transcription polymerase chain reaction: Confirmation of reference genes suitable for normalization. Anal. Biochem., 335: 1-9.
PubMedDirect Link - Huang, L., S.A. Brooks, W. Li, J.P. Fellers, H.N. Trick and B.S. Gill, 2003. Map-based cloning of leaf rust resistance gene Lr21 from the large and polyploid genome of bread wheat. Genetics, 164: 655-664.
PubMedDirect Link - Huggett, J., K. Dheda, S. Bustin and A. Zumla, 2005. Real-time RT-PCR normalisation; strategies and considerations. Genes Immun., 6: 279-284.
CrossRefPubMedDirect Link - Hulbert, S.H., J. Bai, J.P. Fellers, M.P. Pacheco and R.L. Bowden, 2007. Gene expression patterns in near isogenic lines for wheat rust resistance gene Lr34/yr18. Phytopathology, 97: 1083-1093.
CrossRefPubMedDirect Link - Hu, X., Z. Zhang, Z. Fu, P. Xu, W. Song and W. Liu, 2009. Isolation and characterization of an ABC-transporter cDNA clone from wheat (Triticum aestivum L.). Mol. Biol., 43: 946-953.
CrossRefDirect Link - Kavousi, H.R., H. Marashi, J. Mozafari and A.R. Bagheri, 2009. Expression of phenylpropanoid pathway genes in chickpea defense against race 3 of Ascochyta rabiei. Plant Pathol. J., 8: 127-132.
CrossRefDirect Link - Khimani, A.H., A.M. Mhashilkar, A. Mikulskis, M. O’Malley and J. Liao et al., 2005. Housekeeping genes in cancer: Normalization of array data. Biotechniques, 38: 739-745.
PubMedDirect Link - Kim, S. and T. Kim, 2003. Selection of optimal internal controls for gene expression profiling of liver disease. Biotechniques., 35: 456-460.
PubMedDirect Link - Liu, D.W., S.T. Chen and H.P. Liu, 2005. Choice of endogenous control for gene expression in nonsmall cell lung cancer. Eur. Respir. J., 26: 1002-1008.
PubMedDirect Link - Li, Y.C., J.P. Ren, M.J. Cho, S.M. Zhou and Y.B. Kim et al., 2009. The level of expression of thioredoxin is linked to fundamental properties and applications of wheat seeds. Mol. Plant, 2: 430-441.
CrossRefDirect Link - Lopato, S., N. Bazanova, S. Morran, A.S. Milligan, N. Shirley and P. Langridge, 2006. Isolation of plant transcription factors using a modified yeast one-hybrid system. Plant Methods, 2: 3-17.
CrossRefDirect Link - Lossos, I.S., D.K. Czerwinski, M.A. Wechser and R. Levy, 2003. Optimization of quantitative real-time RT-PCR parameters for the study of lymphoid malignancies. Leukemia, 17: 789-795.
Direct Link - Matsui, T., P.K. Bhowmik and K. Yokozeki, 2004. Phenylalanine ammonia-lyase in moso bamboo shoot: Molecular cloning and gene expression during storage. Asian J. Plant Sci., 3: 315-319.
CrossRefDirect Link - Mekki, D.M., B. Liang-yong, W. Jin-yu, Y. Yan, Y. Yabo and H.H. Musa, 2006. Avian E2A expression level variation associated with phenotypic variation of two Chinese indigenous duck breeds, (Anas platyrhynchos). Int. J. Poult. Sci., 5: 1023-1028.
CrossRefDirect Link - Mitter, K., G. Kotoulas, A. Magoulas, V. Mulero and P. Sepulcre et al., 2009. Evaluation of candidate reference genes for QPCR during ontogenesis and of immune-relevant tissues of European seabass (Dicentrarchus labrax). Comp. Biochem. Physiol. B: Biochem. Mol. Biol., 153: 340-347.
CrossRef - Mohammadi, M., N.N.V. Kav and M.K. Deyholos, 2007. Transcriptional profiling of hexaploid wheat (Triticum aestivum L.) roots identifies novel, dehydration-responsive genes. Plant Cell Environ., 30: 630-645.
CrossRefDirect Link - Mohammadi, M., N.N.V. Kav and M.K. Deyholos, 2008. Transcript expression profile of water-limited roots of hexaploid wheat (Triticum aestivum ‘Opata’). Genome, 51: 357-367.
PubMedDirect Link - Mourelatos, Z., L. Abel, J. Yong, N. Kataoka and G. Dreyfuss, 2001. SMN interacts with a novel family of hnRNP and spliceosomal proteins. EMBO J., 20: 5443-5452.
CrossRefDirect Link - Niu, J.S., L.N. Zhang, D.F. Hong and Y.H. Wang, 2005. Cloning, characterization and expression of wheat EDR1 (enhanced disease resistance) gene. J. Plant Physiol. Mol. Biol., 31: 477-484.
PubMedDirect Link - Neuvians, T.P., I. Gashaw, C.G. Sauer, C. von Ostau and S. Kliesch et al., 2005. Standardization strategy for quantitative PCR in human seminoma and normal testis. J. Biotechnol., 117: 163-171.
CrossRef - Paolacci, A.R., O.A. Tanzarella, E. Porceddu and M. Ciaffi, 2009. Identification and validation of reference genes for quantitative RT-PCR normalization in wheat. BMC Mol. Biol., 10: 11-11.
Direct Link - Pfaffl, M.W., A. Tichopad, C. Prgomet and T.P. Neuvians, 2004. Determination of stable housekeeping genes, differentially regulated target genes and sample integrity: BestKeeper-Excel-based tool using pair-wise correlations. Biotechnol. Lett., 26: 509-515.
CrossRef - Schreiber, A.W., T. Sutton, R.A. Caldo, E. Kalashyan and B. Lovell et al., 2009. Comparative transcriptomics in the Triticeae. BMC Genomics, 10: 285-285.
CrossRefPubMedDirect Link - Silberberg, G., K., Baruch and R. Navon, 2009. Detection of stable reference genes for real-time PCR analysis in schizophrenia and bipolar disorder. Anal. Biochem., 391: 91-97.
CrossRef - Skovgaard, K., S.N. Grell, P.M. Heegaard, G. Jungersen, C.B. Pudrith and P.M. Coussens, 2006. Differential expression of genes encoding CD30L and P-selectin in cattle with Johne’s disease: Progress toward a diagnostic gene expression signature. Vet. Immunol. Immunopathol., 112: 210-224.
CrossRef - Traverso, J.A., F. Vignols, R. Cazalis, A.J. Serrato and P. Pulido et al., 2008. Immunocytochemical localization of Pisum sativum TRXs f and m in non-photosynthetic tissues. J. Exp. Bot., 59: 1267-1277.
CrossRef - Tricarico, C., P. Pinzani, S. Bianchi, M. Paglierani and V. Distante et al., 2002. Quantitative real time reverse transcription polymerase chain reaction: Normalization to rRNA or single housekeeping genes is inappropriate for hunman tissue biopsies. Anal. Biochem., 309: 293-300.
Direct Link - Vandesompele, J., K. De Preter, F. Pattyn, B. Poppe, N. van Roy, A. De Paepe and F. Speleman, 2002. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol., 3: 1-11.
CrossRefDirect Link - Warrington, J.A., A. Nair, M. Mahadevappa and M. Tsyganskaya, 2000. Comparison of human adult and fetal expression and identification of 535 houskeeping/maintenance genes. Physiol Genomics., 2: 143-147.
Direct Link - Wang, J.R., L. Wang, S. Gulden, H. Rocheleau AND M. Balcerzak, 2010. RNA profiling of fusarium head blight-resistant wheat addition lines containing the Thinopyrum elongatum chromosome 7E. Can. J. Plant Pathol., 32: 188-214.
CrossRefDirect Link - Wang, T. and M.J. Brown, 1999. mRNA quantification by real time TaqMan polymerase chain reaction: Validation and comparison with RNase protection. Anal. Biochem., 269: 198-201.
Direct Link - Wang, X., C. Tang, G. Zhang, Y. Li and C. Wang et al., 2009. cDNA-AFLP analysis reveals differential gene expression in compatible interaction of wheat challenged with Puccinia striiformis f. sp. tritici. BMC Genomics, 10: 289-289.
Direct Link - Xu, L., J. Jia, J. Lv, X. Liang, D. Han, L. Huang and Z. Kang, 2010. Characterization of the expression profile of a wheat aci-reductone-dioxygenase-like gene in response to stripe rust pathogen infection and abiotic stresses. Plant Physiol. Biochem., 48: 461-468.
CrossRef - Xu, H., X. Jiang, K. Zhan, X. Cheng, X. Chen, J.M. Pardo and D. Cui, 2008. Functional characterization of a wheat plasma membrane Na+/H+ antiporter in yeast. Arch. Biochem. Biophys., 473: 8-15.
CrossRef - Roelfs, A.P., R.P. Singh and E.E. Sari, 1992. Rust Diseases of Wheat: Concepts and Methods of Disease Management. CIMMIYT, Mexico, ISBN: 9789686127478, Pages: 81.
Direct Link - Sitthiwong, K., M. Toshiyuki, O. Nobuyuki and K. Yusuke, 2007. Cloning and sequencing of sucrose synthase cDNA from vegetable soybean and its expression during storage at 20�°C. Biotechnology, 6: 184-192.
CrossRefDirect Link - Thellin, O., W. Zorzi, B. Lakaye, B.D. Borman and B. Coumans et al., 1999. Housekeeping genes as internal standards: Use and limits. J. Biotechnol., 75: 291-295.
CrossRefDirect Link