
Toshiyuki Matsui
Not Available
Pankaj Kumar Bhowmik
Not Available
Kyosuke Yokozeki
Not Available
Asian Journal of Plant Sciences
Year: 2004 | Volume: 3 | Issue: 3 | Page No.: 315-319







ABSTRACT
In plants, phenylalanine ammonia-lyase (PAL; EC 4.3.1.5) catalyzes the conversion of L-phenylalanine to trans cinnamic acid in the first step of phenylpropanoid pathway. We previously isolated cDNA clones encoding PAL from asparagus spears and observed that the level of transcripts correlated well with increase enzyme activity and lignification. In order to identify factors regulating the postharvest deterioration of another important Asian vegetable bamboo (Phyllostachys edulis) shoot, we have now isolated PAL (pBA-PAL), cDNA clone from a cDNA library prepared from harvested bamboo shoot using reverse transcriptase-polymerase chain reaction. The partial cDNA clone encodes an mRNA of 527 bp and the derived amino acid sequence is highly homologous to PAL from rice, barely, maize and asparagus. Northern blot analysis showed increasing trend in the level of pBA-PAL mRNA reaching a peak after 3 d of storage at 20°C, which coincided well with enzyme activity. Our results demonstrate that during the postharvest storage of bamboo shoot PAL activity is regulated by increase level of pBA-PAL mRNA suggesting that the increase might be a response to the wounding associated with harvest.
PDF Abstract XML References Citation
How to cite this article
Toshiyuki Matsui, Pankaj Kumar Bhowmik and Kyosuke Yokozeki, 2004. Phenylalanine Ammonia-Lyase in Moso Bamboo Shoot: Molecular Cloning and Gene Expression During Storage. Asian Journal of Plant Sciences, 3: 315-319.
DOI: 10.3923/ajps.2004.315.319
URL: https://scialert.net/abstract/?doi=ajps.2004.315.319
DOI: 10.3923/ajps.2004.315.319
URL: https://scialert.net/abstract/?doi=ajps.2004.315.319
INTRODUCTION
Bamboo shoots are minor ingredient for “tsukemono” (pickled vegetables)[1]. Japanese use dried bamboo shoots mainly for Chinese style cooking[2]. Two kinds of bamboo shoots exist in Japan. Machiku shoots (large type), suitable for freezing because they suffer only minimal loss of texture and flavour are mainly supplied by Taiwan and Thailand, but Taiwanese supply has been decreasing[3]. Moso shoots (thick stemmed) is less appropriate for freezing due to many small hollow areas within the plants. Imports are limited pending new technology and China supplies most. If diced, they can be used for Chinese dishes such as spring rolls (harumaki) and steamed meat buns (nikuman). Currently it is limited to restaurants and processors. However, without appropriate handling, fiber formation or lignification, developed quickly, in harvested bamboo shoot and discoloration or browning occurred subsequently[4].
Lignin, the substance that lends fibers their toughness, is polymerized from cinnamyl alcohols derived from shikimic acid pathway[5]. PAL catalyzing the conversion of L-phenylalanine to trans-cinnamic acid in this pathway has been considered to play an important role as a key enzyme. Fluctuation in PAL activity has been shown to be a key element controlling the synthesis of lignin and in many cases, an increase in the amount of PAL mRNA has been shown to underlie the increase of PAL activity[6-9]. In elicited cultured pine cells, for instance, the induction of PAL activity occurs concurrently with increases in the activities of lignin-specific enzymes and is followed by the deposition of apparently genuine gymnosperm lignin in the cell walls[10,11]. PAL transcripts increased in response to fruit ripening and wounding in the mesocarp tissue of Cucurbita maxima[12]. In freshly harvested asparagus spears PAL activity increased upon excision and incubation[13,14]. Increased PAL activity was also observed in intact spears given an extended period of cold treatment. To our knowledge, there is no study on the role of PAL activity and gene expression on storage quality and shelf life of moso bamboo shoot. However, Chen et al.[4] reported a rapid increase of PAL activity in harvested bamboo shoot stored at relatively high temperature, 30°C.
This and very similar results from Kozukue et al.[15] point to the significance of PAL during storage of bamboo shoot but no PAL gene expression is detected during storage of shoot.
This study reports the cloning and characterization of a partial PAL cDNA from harvested bamboo shoots. By RNA blot analysis and in situ RNA hybridization the pattern of expression of PAL gene has also been reported in relation to PAL enzyme activity during storage.
MATERIALS AND METHODS
Plant material: Moso bamboo shoots were harvested from farmer’s field in Kagawa Prefecture, Japan in April, 2002. The shoots were immediately brought to the laboratory (Faculty of Agriculture, Kagawa University, Japan) and stored at 1 and 20°C in perforated plastic bags for 0, 1, 3, 6 and 9 d in darkness. After storage treatment shoots were cut into small pieces (3 mm in thickness), weighed and immediately stored at -80°C until needed for PAL enzyme analysis and RNA extraction.
Extraction and assay of PAL activity: Two g of shoots were homogenized at 2°C with a mortar and pestle in 10 ml of 0.1 M borate buffer, pH 8.8 and 1.0 g PVPP. A further 10 ml of the buffer was added to the homogenate which was then centrifuged at 14, 000 rpm for 20 min and the supernatant was used for enzymatic assays. Protein concentration was measured by Lowry[16] method. PAL activity was determined spectrophotometrically by measuring the increase in A290 due to the formation of trans-cinnamic acid. The reaction mixture consisted of 50 mM borate buffer, pH 8.8, 20 mM L-phenylalanine and the enzyme preparation in a total volume of 3 ml. A sample without L-phenylalanine was used as a blank. The activity was expressed as nmol trans-cinnamic acid formed per h per mg of protein.
RNA extraction and amplification of poly (A)+ RNA by RT-PCR: Total RNA was extracted according to the Hot Borate method of Wan and Wilkins[17]. The first strand cDNA was synthesized from 2 g of the total RNA by reverse transcriptase with Oligo-(dT) primer according to the instruction of SUPER SCRIPTTM Preamplification System for First Strand cDNA Synthesis (GIBCOBRL, Tokyo, Japan). PCR was performed in a total volume of 25 μl containing the first strand cDNA reaction products, 10xPCR Buffer, MgCl2, dNTP, First Start Taq DNA Polymerase (Roche) and primers. The primers (5'-ATYGAGGCTGCTGCYATTATG-3' as the upstream primer and 5'-ACATCTTGGTTGTGYTGCTC-3' as the downstream primer) were designed and synthesized on the basis of amino acid domains (IEAAAIM and AEQHNQD, respectively) conserved in various PAL genes. The Sal 1 and Not 1 restriction site sequences were also included at 5'-end of the sense and antisense primer, to facilitate cloning of PCR product. The PCR procedure started with 10 min at 95°C and was carried out 35 cycles of 30 s at 95°C, 30 s at 50°C and 30 s at 72°C and 10 min at 72°C with ASTEC Program Temperature Control System PC-700. The PCR products were confirmed by agarose gel electrophoresis.
Cloning and sequencing of cDNA, sequence data analysis, synthesis of RNA probe and Northern blot analysis were performed as described in our previous reports[14].
GenBank accession number: The nucleotide sequence data reported in this paper will appear in the DDBJ/EMBL/GenBank nucleotide sequence databases with the accession number AB 162422.
RESULTS
Changes in PAL activity: During storage at 20°C, PAL activity of bamboo shoots started to increase just after harvest reaching a peak at 3 d (Fig. 1) .
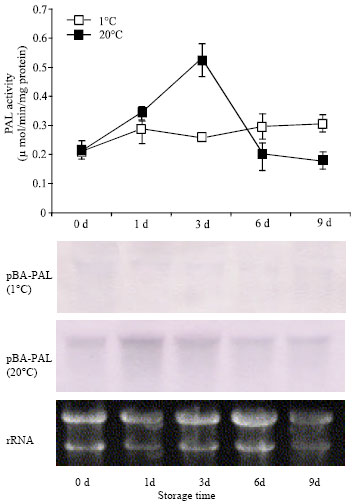 | |
| Fig. 1: | Changes in PAL activity and expression of pBA-PAL during storage of bamboo shoot at 1°C and 20°C. Each point represents the mean of three replicates and bars show SD about mean. Equal loading of RNA was confirmed by staining a gel with ethidium bromide |
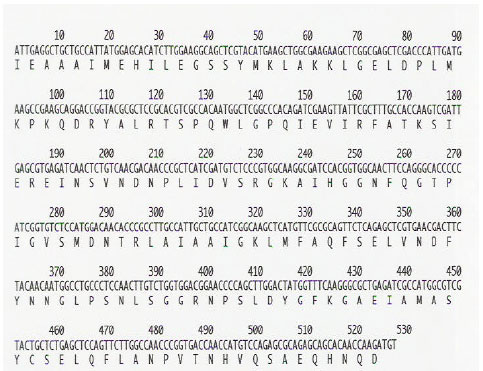 | |
| Fig. 2: | Nucleotide sequence and deduced amino acid sequence of the cDNA clone corresponding to pBA-PAL. The predicted amino acid sequence is given in single-letter code for each amino acid. The arrows indicate the positions of degenerated primers (→sense, ←antisense) used for RT-PCR. Numbering refers to total nucleotide residues on each line |
 | |
| Fig. 3: | Comparison of the deduced amino acid sequences from asparagus (AB102677), barley (Z49145), maize (L77912), pea (D10001), rice (X97946), tobacco (M844466) and tomato (M90692) by multi alignment. The amino acid residues are numbered at the beginning and end of the sequences on each line. Asterisks denote the amino acid residues those are identical. Dashes in the amino acid sequences represent gaps introduced to maximize alignment of the polypeptides |
 | |
| Fig. 4: | Phylogenetic tree of the alignment of pBA-PAL deduced amino acid sequence with other PAL genes in the database. Protein sequences were aligned using UPGMA and phylogenetic tree was constructed using GENETYX-MAC software |
| Table 1: | Percentage of nucleotide and deduced amino acid homology between PAL from bamboo shoot and other plants in the databases |
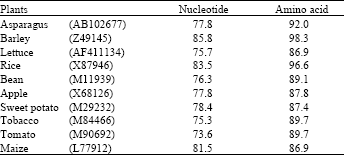 | |
| Bamboo PAL (AB162422) is calculated as 100 % | |
At 3 d the PAL activity was observed about three folds higher than the initial level. After that the activity sharply declined at 6 d and almost remain unchanged until 9 d. On the other hand, PAL activity of bamboo shoots stored at 1°C only slightly increased after 1 d and then almost remain unchanged. PAL activity at 20°C was significantly higher than that at 1°C.
Isolation and identification of cDNA clone: The cDNA pBA-PAL is a partial clone encoding a harvest-induced transcript from bamboo shoots. The encoded mRNA is 527 bp long (Fig. 2) and is highly homologous to the gene of other plant (Fig. 3). The pAS-PAL sequence is 85.8% identical to PAL from barley (X87946), 83.5% identical to PAL from rice (X87946) and 81.5% identical to maize (L77912) PAL, all of which are wound-induced gene (Table 1). Allowing for conservative amino acid substitutions, the similarities are 98.3, 96.6 and 86.9% for the barley, rice and maize sequences, respectively.
PAL gene expression: The cDNA pBA-PAL is a partial clone encoding a PAL transcript from moso bamboo shoot. The encoded mRNA is 527 bp long (Fig. 2) and is highly homologous to the PAL genes of other plant (Fig. 3). The expression of pBA-PAL was induced by wounding due to harvest. Maximum pBA-PAL expression appeared after 3 d of storage and coincided with the peak of enzyme activity (Fig. 1). A phylogenetic tree (Fig. 4) was generated from the alignment of the deduced amino acid sequences of pAS-PAL and other PAL gene in the database. The pBA-PAL (AB162422) and PAL from barley (Z49145) strongly clustered together in a subgroup belonged to the monocotyledon, having closest relationship with rice and maize gene.
DISCUSSION
During storage, bamboo shoots, similar to asparagus spears, undergo texture changes because of changes in PAL activity. We recently showed that induction of PAL activity in harvested asparagus is regulated by transcription of pAS-PAL in response to the wounding associated with harvest[14]. To understand the molecular basis of induction of PAL activity in harvested bamboo shoots during storage we have isolated a cDNA for PAL from harvested bamboo shoots and found that it is wound induced. The clone, pAS-PAL had high sequence similarity to rice, maize and barley cDNA that were also associated with wounding. The development of PAL activity in bamboo shoots was similar in a number of respects to asparagus spears and other wound induced tissue systems[2,14,18]. There was an increase in PAL activity in both top and bottom portions of the spears which continued for about 24 h. After that PAL activity started to decline. Although PAL activity in bamboo shoots reached the peak after 3 d of storage, it followed almost same pattern like asparagus spears.
Our Northern blot results also coincided well with Chen et al.[4] who reported enhanced PAL activity in harvested bamboo shoots stored at ambient temperature. The expression of pBA-PAL was also correlated well with the enzyme activity published by Kozukue et al.[15]. As in cut-injured sweet potatoes[19], the enhancing activity of PAL in harvested bamboo shoots is partly due to the PAL de novo synthesis especially for shoots stored at 20°C. Although we did not measure lignin content in this experiment but our report with asparagus[14] and the report of Chen et al.[4] clearly demonstrated that lignin production is induced to some extent by increased PAL activity. Alternatively, ethylene produced by the wounded tissue may induce PAL activity as observed in some other plant tissues[12,20].
Therefore, from our result it can be concluded that we have isolated a partial cDNA clone encoding PAL of moso bamboo shoot. Phylogenetic analysis of PAL sequences has revealed the existence of at least two major branches that contain characteristic conserved amino acid sequences, monocotyledon and dicotyledon. The pAS-PAL belonged to the subgroup wound-induced monocotyledon as it was highly homologous to barley PAL and was closely related to PAL from Maize. Using this cDNA clone, we could suggest that bamboo shoot having analogy with asparagus spears undergo texture changes during storage which is regulated by the increased PAL activity and the high induction of PAL mRNA.
REFERENCES
- Lawton, M.A. and C.J. Lamb, 1987. Transcriptional activation of plant defense genes by fungal elicitor, wounding and infection. Mol. Cell. Biol., 1: 335-341.
PubMed - Campbell, M.M. and B.E. Ellis, 1992. Fungal elicitor-mediated responses in pine cell cultures. I. Induction of phenylpropanoid metabolism. Planta, 186: 409-417.
CrossRefDirect Link - Kato, M., Y. Hayakawa, H. Hyodo, Y. Ikoma and M. Yano, 2000. Wound induced ethylene synthesis and formation of 1-aminocyclopropane-1-carboxylate (ACC) synthase. ACC oxidase, phenylalanine ammonia-lyase and peroxidase in wounded mesocarp tissue of Cucurbita maxima. Plant Cell Physiol., 41: 440-447.
Direct Link - Bhowmik, P.K., T. Matsui, H. Suzuki and Y. Kosugi, 2003. A phenylalanine ammonia-lyase gene from asparagus: cDNA cloning, sequence and expression in response to wounding. Asian J. Plant Sci., 2: 425-430.
CrossRefDirect Link - Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - Wan, C.Y. and T.A. Wilkins, 1994. A modified hot borate method significantly enhances the yield of high quality RNA from cotton (Gossypium hirsutum L.). Anal. Biochem., 223: 7-12.
Direct Link