
Huiwon No
Department of Biomedical Science, Daegu Catholic University, Gyeongsan-Si, Republic of Korea
Jin-Kyung Kim
Department of Biomedical Science, Daegu Catholic University, Gyeongsan-Si, Republic of Korea
LiveDNA: 82.35300
International Journal of Pharmacology
Year: 2021 | Volume: 17 | Issue: 1 | Page No.: 48-56







ABSTRACT
Background and Objective: Rugchalcones are isolated from the flowers of Rosa rugose. The pharmacological activities of rugchalcones and their derivatives are rarely reported. In the present study, we investigated the ability of Rugchalcone Derivative 4 (RD4) to regulate the inflammatory response in the RAW264.7 murine macrophage cell line. Material and Methods: To determine the anti-inflammatory effects of RD4, the lipopolysaccharide (LPS)-induced synthesis of inflammatory mediators in RAW264.7 cells were measured by Enzyme-Linked Immunosorbent Assay (ELISA) and Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR). The anti-inflammatory mechanism of RD4 was investigated via western blot analysis of the activation of Nuclear Factor (NF)-κB and expression of Heme Oxygenase (HO)-1. Results: RD4 inhibited not only the synthesis of Nitric Oxide (NO) but also the expression of inducible NO synthase by LPS in the RAW264.7 murine macrophage cell line. RD4 inhibited the release of pro-inflammatory cytokines induced by LPS in RAW264.7 cells, including Interleukin (IL)-1β, IL-6 and tumour necrosis factor-α. The underlying anti-inflammatory mechanism of RD4 was correlated with the down-regulation of NF-κB and induction of HO-1. Conclusion: The data collectively indicate that RD4 inhibited the synthesis of several inflammatory mediators via regulation of NF-κB and HO-1 and represents a potential treatment for various inflammatory diseases.
PDF Abstract XML References Citation
Copyright: © 2021. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Huiwon No and Jin-Kyung Kim, 2021. Anti-Inflammatory Activity of Rugchalcone Derivative on Lipopolysaccharides-Stimulated RAW264.7 Cells. International Journal of Pharmacology, 17: 48-56.
DOI: 10.3923/ijp.2021.48.56
URL: https://scialert.net/abstract/?doi=ijp.2021.48.56
DOI: 10.3923/ijp.2021.48.56
URL: https://scialert.net/abstract/?doi=ijp.2021.48.56
INTRODUCTION
Inflammation is a complex process involving various types of cells and substances produced in defence against external challenges such as infections, injuries or toxins that ultimately restore normal tissue structure and function. It is self-limiting and strongly associated with the down-regulation of pro-inflammatory protein expression, increased anti-inflammatory protein levels and reversal of vascular changes that facilitate early immune cell recruitment1,2. Abnormal inflammatory responses contribute to the aetiology of many inflammatory diseases, including bronchitis, gastritis, inflammatory bowel disease, multiple sclerosis and rheumatoid arthritis3-5. Therefore, the control of inflammation is essential for the prevention and treatment of various diseases.
Macrophages are representative cells that drive the inflammatory response and serve as an essential interface between innate and adaptive immunity6,7. Stimulation of macrophages by various agents such as lipopolysaccharide (LPS) culminates in the activation of two distinct downstream signalling pathways: the transcription factor Nuclear Factor (NF)-κB pathway and the Mitogen-Activated Protein Kinase (MAPK) pathway1,8. Expression of major inflammatory mediators including Nitric Oxide (NO) and inflammatory cytokines are driven by these two pathways1,8-10.
NF-κB is an essential transcription factor in the control of inflammatory responses1,8,10-12. NF-κB regulates the expression of genes that encode inflammatory cytokines, adhesion molecules, chemokines, growth factors and inducible enzymes such as inducible NO Synthase (iNOS)11,13. NF-κB activity is generally blocked in the cytoplasm via association with an inhibitor of NF-κB (Iκ-B). Upon activation, Iκ-B undergoes phosphorylation and degradation, followed by nuclear translocation and binding with DNA, resulting in induction of transcription of various inflammatory mediators11,13.
Heme Oxygenase (HO)-1, an antioxidant enzyme involved in the stress response, catalyzes the degradation of heme, biliverdin and carbon monoxide and also plays an important role in inflammation14. Recent clinical studies reported the application of pharmacological compounds to up-regulate HO-1 in specific anti-inflammatory treatments14,15. Indeed, a variety of drugs, including statins, 5-amino salicylic acid and polyphenols are used in standard therapies to provide anti-inflammatory protection via induction of HO-114-17.
Rugchalcones are 2-aroyl benzofuran derivatives isolated from the flowers of Rosa rugosa with anti-tobacco mosaic virus activity18. However, no studies have reported the physiological and pharmacological activity of rugchalcones and their derivatives. The previous studies demonstrated the efficient synthesis of rugchalcone A, rugchalcone B and their derivatives19. Besides, their anti-inflammatory potential was screened via inhibition of NO production in RAW264.7 murine macrophages19. The study confirmed that the Rugchalcone Derivative 4 (RD4) synthesized by us was likely to show anti-inflammatory activity19.
To confirm the physiological and pharmacological activity of bioactive natural products and analogues, the anti-inflammatory effects of RD4 and the underlying mechanisms for RAW264.7 cells were investigated in this study.
MATERIALS AND METHODS
Study area: This study was conducted from January-November, 2020 at the Department of Biomedical Science, Immunology Laboratory, Daegu Catholic University, Korea.
Chemicals and Reagents: RD4 (Fig. 1a) was synthesized as previously described19. All reagents including LPS were obtained from Sigma-Aldrich (St. Louis, MO) unless otherwise indicated. Dulbecco’s Modified Eagle’s Medium (DMEM), Fetal Bovine Serum (FBS), penicillin and streptomycin were obtained from Hyclone (Logan, UT, USA). ELISA kits for assaying interleukin (IL)-1β, IL-6 and Tumor Necrosis Factor (TNF)-α were purchased from eBioscience (San Diego, CA, USA). Antibodies targeted against iNOS, HO-1, Iκ-Bα, phospho-NF-κB p65, phospho-ERK, phospho-JNK, phospho-p38, JNK, ERK and p38 were procured from Cell Signaling Technology (Danvers, MA, USA).
Cell culture and cell viability assay: RAW264.7 murine macrophages were obtained from the Korean Cell Bank (Seoul, Korea) and cultured in DMEM containing 10% FBS, 100 U mL–1 penicillin and 100 μg mL–1 streptomycin. The cells were incubated in an atmosphere of 5% CO2 at 37°C and sub-cultured every 3 days. The effects of RD4 on cell viability were tested using the CellTiter 96® AQueous One Solution Assay for cell proliferation (Promega, Madison, WI) according to the manufacturer's instructions.
Measurement of nitrites and cytokines: RAW264.7 cells were plated at a density of 2×105 cells in a 48-well cell culture plate with 500 μL of culture medium and incubated for 12 hrs. They were then treated with RD4 in a medium containing LPS (500 ng mL–1) and incubated for another 18 hrs. The number of nitrites generated was measured using the Griess reagent system (Promega). The amount of IL-1β, IL-6 and TNF-α in the cell-culture supernatant was measured using an ELISA kit (eBioscience, San Diego, CA) according to the manufacturer’s instructions.
Quantitative Real-Time reverse-transcription Polymerase Chain Reaction (qRT-PCR): Total RNA was isolated from RAW264.7 cells using TRIzol reagent (Invitrogen, Carlsbad, CA, USA). DNA was eliminated from total RNA using RNA Qualified RNase-Free DNase (Promega) and cDNA was synthesized by GoScriptTM Reverse Transcription System (Promega). The qRT-PCR assay was carried out with a LightCycler (Roche Diagnostics, Basel, Switzerland) using the LightCycler FastStart DNA Master SYBR Green I (Roche Diagnostics). All the experiments were repeated twice in triplicate each time. Transcripts of Glyceraldehyde-3-Phosphate Dehydrogenase (GAPDH) as a housekeeping gene were quantified as endogenous RNAs of reference to normalize each sample. Relative quantities were estimated by the -ΔΔCt method.
The primers used in this study corresponded to mouse:
| • | mouse iNOS: F 5'-CCT CCT CCA CCC TAC CAA GT-3', R 5'-CAC CCA AAG TGC TTC AGT CA-3' |
| • | mouse IL-1β: F 5'-TTC TCCACA GCC ACA ATG AG-3', R 5'-ACG GAC CCC AAA AGA TGA AG-3' |
| • | mouse IL-6: F 5'-CCT CTG GTC TTCTGG AGT ACC-3', R 5'-ACT CCT TCT GTG ACT CCA GC-3' |
| • | mouse TNF-α: F 5'-TGT CTC AGC CTC TTC TCA TT-3' R 5'-AGA TGA TCT GAG TGT GAG GG-3' |
| • | mouse HO-1: F 5'-CCT CAC TGG CAG ATC ATC-3’ R 5'-CCT CGT GGA GAC GCT TTA CAT A -3', and |
| • | mouse GAPDH: F 5'-TCT TGC TCA GTG TCC TTG C-3', R 5'-CTT TGT CAA GCT CAT TTC CTG G-3' |
Western blot analysis: The cells were then rinsed with ice-cold PBS, followed by the addition of 100 μL of a PRO-PREPTM Protein Extraction Solution (Seongnam, Korea) to each sample. Proteins were denatured in Sodium Dodecyl Sulfate (SDS). The samples were subjected to SDS Polyacrylamide Gel Electrophoresis (PAGE), followed by transfer to a nitrocellulose membrane. Non-specific binding was inhibited by exposing the cell membranes to TBST (50 mM Tris-HCl, pH 7.5, 150 mM NaCl and 0.1% Tween-20) containing 5% non-fat milk for 1 h at room temperature. The membranes were then incubated overnight with the indicated primary antibodies at 4°C. The membranes were then washed with phosphate-buffered saline containing 0.1% Tween 20) and incubated for 1 h with anti-rabbit IgG or anti-mouse IgG conjugated with Horseradish Peroxidase (HRP). After washing, the protein bands were then visualized using enhanced chemiluminescence reagent (Thermo Fisher Scientific) and DAVINCH-Chemi CAS-400 SM (Davinch-k, Seoul, Korea). Band densities were measured using Total Lab software (Davinch-k).
Statistical analysis: Values were expressed as the Mean±SEM of the results of at least three experiments. The values were then evaluated via One-Way Analysis of Variance (ANOVA) with Bonferroni multiple comparison post hoc tests using Prism 6.0 software (GraphPad Software, San Diego, CA, USA). Differences with p<0.05 were considered statistically significant.
RESULTS
Effect of rugchalcone derivative on cell viability: Selection of the appropriate nontoxic range of RD4 is important for further experiment. Therefore, the cytotoxicity of RD4 in RAW264.7 macrophages was evaluated by measuring the optical density at 490 nm using the MTS assay for 24 hrs. As shown in Fig. 1b, no cytotoxicity of RD4 in RAW264.7 cells occurred up to 50 μM.
Effect of RD4 on NO production and iNOS expression in LPS-stimulated RAW264.7 cells: First, the effect of RD4 on NO synthesis in LPS-stimulated RAW264.7 cells was evaluated, because overproduction of NO by iNOS has been implicated in the pathology of several inflammatory disorders. As shown in Fig. 2a, the concentration of NO was decreased by RD4 addition in LPS-stimulated RAW264.7 cells.
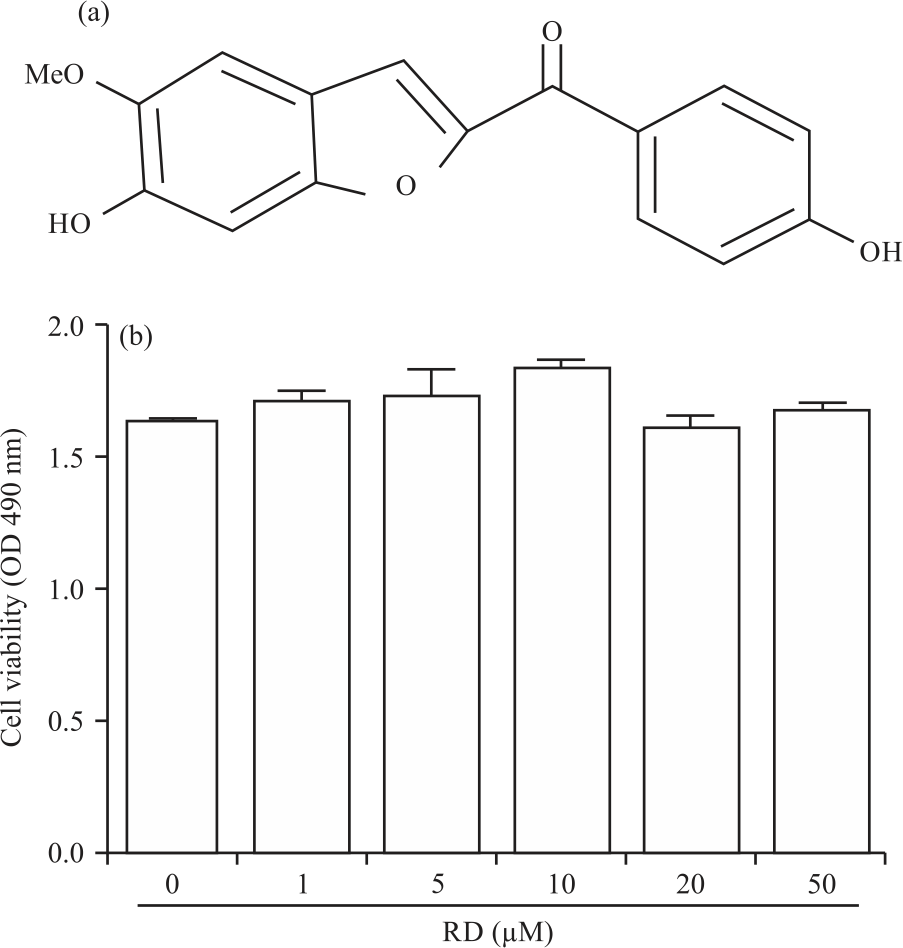 |
| Fig. 1(a-b): | Effect of RD4 on cell viability in RAW264.7 cells (a) Chemical structure of RD4 and (b) RAW264.7 cells were treated with the indicated concentrations of RD4 for 24 hrs and proliferation was determined. The results are expressed as Mean±SEM of three independent experiments |
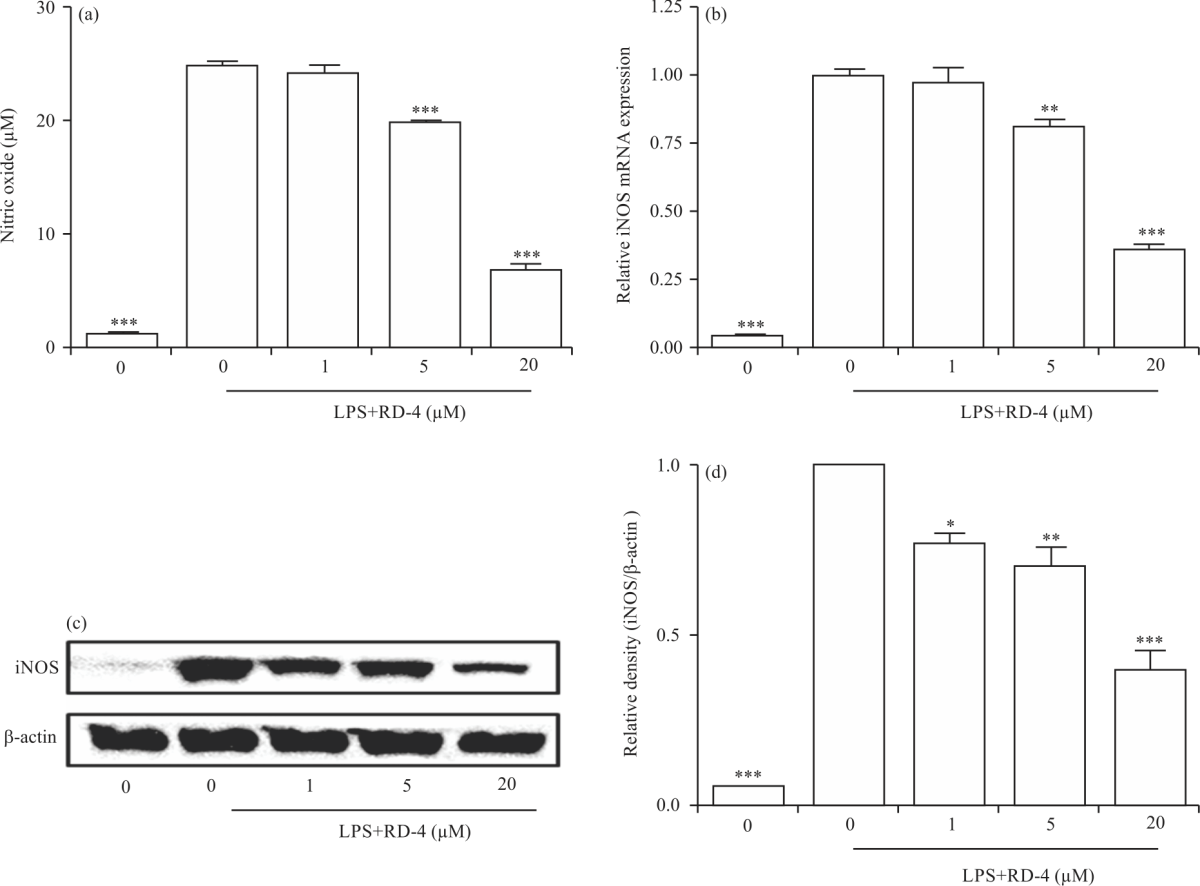 |
| Fig. 2(a-d): | Effects of RD4 on LPS-induced NO synthesis and iNOS expression RAW264.7 cells were treated with the indicated concentration of RD4 in the presence of 500 ng mL–1 of LPS or with LPS alone for 24 hrs and (a) NO release, (b) iNOS mRNA levels were determined, (c) Thirty micrograms of protein obtained from each cell lysate were resolved by 10% SDS-PAGE for iNOS determination. The β-actin expression serves as a loading control and (d) Densitometric analysis of iNOS protein expression. The results are reported as Mean±SEM of three independent experiments. Statistical significance is based on the difference when compared with LPS-stimulated cells (*p<0.05, **p<0.01, ***p<0.001) |
The nitrite concentrations in LPS-stimulated RAW264.7 cells and exposed to 20 μM RD4 were 25.4±0.1 and 7.8±1.4 μM, respectively (Fig. 2a).
We further tested whether NO inhibition by RD4 was caused by decreased expression of iNOS mRNA and reduced protein expression of iNOS in RD4-treated cells compared with LPS-treated cells. The relative levels of iNOS mRNA averaged 0.94±0.07 in LPS-treated RAW264.7 cells and decreased to 0.32±0.05 in the presence of 20 μM RD4 (Fig. 2b). Besides, concentration-dependent inhibition was also observed in iNOS protein levels (Fig. 2c, d). As shown in 2d, the relative protein expression level of iNOS was 1 in RAW264.7 cells stimulated with LPS but decreased to 0.70±0.05 and 0.40±0.05 for 5 and 20 μM RD4 concentrations, respectively.
Effect of RD4 on pro-inflammatory cytokine expression in LPS-stimulated RAW264.7 cells: IL-1β, IL-6 and TNF-α are pro-inflammatory cytokines, which mediate chronic inflammation. The attenuation of these cellular cytokines is essential to avoid further inflammatory response. Therefore, the level of cytokine synthesis in LPS-stimulated RAW264.7 cells was evaluated via ELISA. Without the LPS stimulation, the concentrations of IL-1β, IL-6 and TNF-α in the cell culture medium, by ELISA were 28.1±3.1 pg mL–1, 0.02±0.01 and 0.17±0.06 ng mL–1, respectively (Fig. 3a-c). Exposure of the cells to LPS significantly increased the production of IL-1β, IL-6 and TNF-α, when compared with an untreated control group (106.3±6.3 pg mL–1, 2.79±0.12 and 10.1±0.15 ng mL–1, respectively). However, the RD4 treatment suppressed the synthesis of these cytokines significantly in a dose-dependent manner. At the highest concentration of RD4 tested (20 μM), IL-1β, IL-6 and TNF-α production decreased to 59.3±3.1 pg mL–1, 0.11±0.02 and 8.5±0.12 ng mL–1, respectively (Fig. 3c).
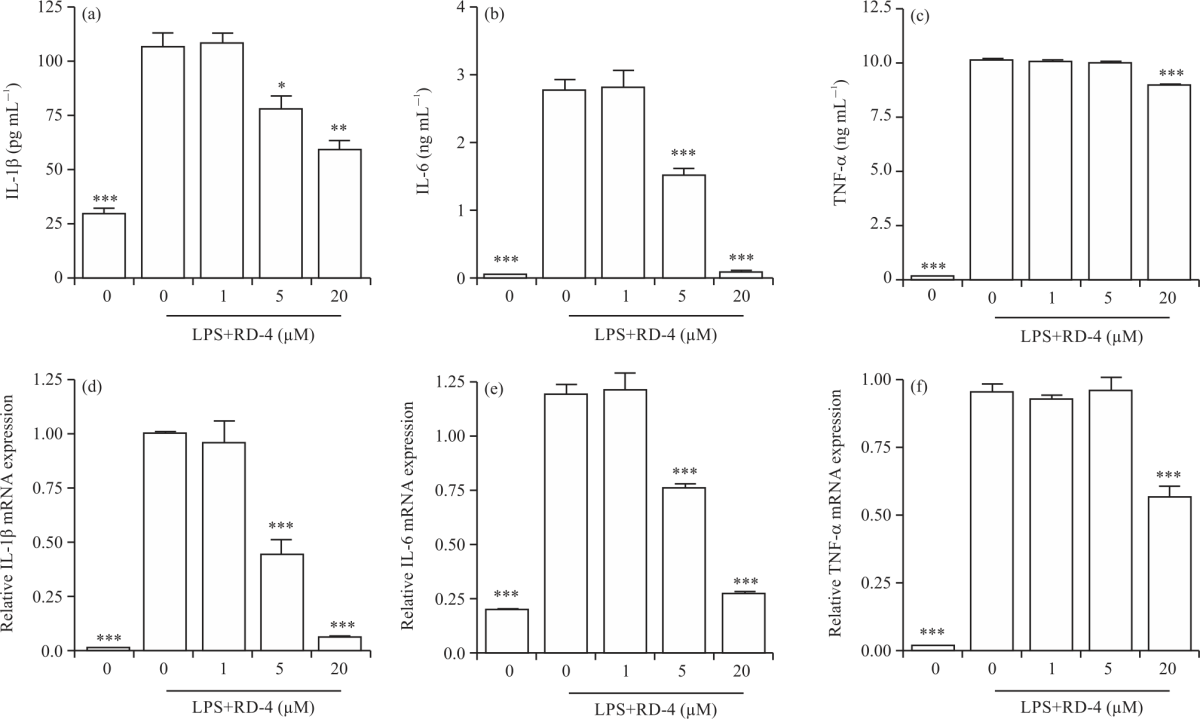 |
| Fig. 3(a-f): | Effects of RD4 on LPS-induced pro-inflammatory cytokine production in murine macrophages RAW264.7 cells were treated with indicated concentrations of RD4 with LPS (500 ng mL–1) or with LPS alone for 24 hrs, (a-c) The cell culture media were then collected and the amounts of IL-1β, IL-6 and TNF-α released were measured and (d-f) The mRNA levels of IL-1β, IL-6 and TNF-α were measured via qRT-PCR. The results are expressed as Mean±SEM of three independent experiments. Statistical significance is based on the difference when compared with LPS-stimulated cells (*p<0.05, **p<0.01, ***p<0.001) |
Among the three types of pro-inflammatory cytokines, the IL-6 reduction by RD4 treatment was the most comparable to that of the untreated control group (Fig. 3b).
To assess the effect of RD4 on mRNA expression of pro-inflammatory cytokines in LPS-stimulated macrophages, we measured their gene expression levels using qRT-PCR. Only LPS-stimulated treatment was chosen as the control (with an mRNA fold induction of 1). Untreated cells did not induce mRNA expression of the three pro-inflammatory cytokines, whereas LPS induced their expression (Fig. 3d-f). Treatment of cells with RD4 inhibited the mRNA expression of IL-1β and IL-6 in a dose-dependent manner with a significant inhibitory effect at 5 and 20 μM concentrations (Fig. 3d-e). Similar to the ELISA results, the level of TNF-α mRNA level induced by LPS significantly decreased only in 20 μM RD4 (Fig. 3f). These findings suggest that RD4 strongly inhibits various inflammatory mediators including NO and pro-inflammatory cytokines.
RD4 blocks the NF-κB pathway in LPS-stimulated RAW264.7 cells: Since NF-κB is a key transcription factor regulating the level of pro-inflammatory mediators in activated macrophages, we next investigated whether RD4 abrogated the NF-κB signalling pathway, which is implicated in the transcriptional regulation of inflammatory mediators in LPS-stimulated RAW264.7 cells. The cells were treated with LPS with or without RD4. The phosphorylated levels of NF-κB p65 subunit and cytosolic levels of I κ-Bα were analyzed by western blot. LPS stimulation induced NF-κB p65 phosphorylation and I κ-Bα degradation in RAW264.7 cells (Fig. 4a-b), however, RD4 treatment attenuated the phosphorylation of NF-κB p65 and I κ-Bα degradation in LPS-stimulated RAW264.7 cells (Fig. 4a-b).
Next, we investigated the role of RD4 in the LPS-induced activation of the MAPK signalling pathway. We assessed the effect of RD4 on LPS-induced phosphorylation of p38, JNK and ERK using three different phospho-specific antibodies. Results indicated that p38, JNK and ERK showed slight phosphorylation in cells of the untreated control group (Fig. 4c-d). LPS treatment significantly increased the activation of MAPKs by strengthening the phosphorylation of p38, JNK and ERK.
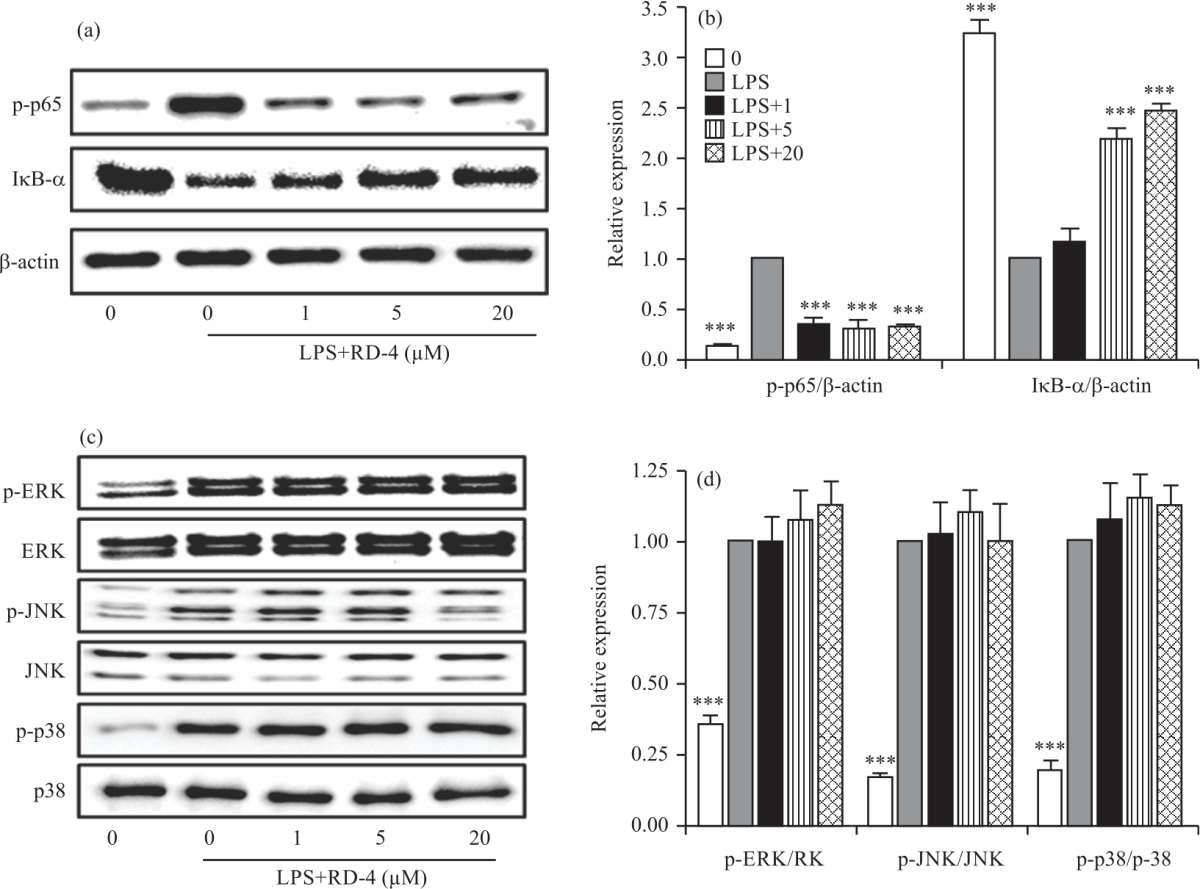 |
| Fig. 4a-d: | Effects of RD4 on LPS-induced NF-κB activation and MAPK phosphorylation in RAW264.7 cells RAW264.7 cells were treated with the indicated concentration of RD4 in the presence of 500 ng mL–1 of LPS or with LPS alone for 30 min. Western blot analysis was performed to determine (a) NF-κB, (B) MAPK activation as described in Materials and Methods and (c-d) Data analysis was performed using total lab software by measuring the integrated band densities following background subtraction. The results are reported as Mean±SEM of three independent experiments. Statistical significance is based on the difference when compared with LPS-stimulated cells (***p<0.001) |
 |
| Fig. 5(a-c): | Effects of RD4 on HO-1 induction in murine macrophages RAW264.7 cells were treated with indicated concentrations of RD4 for 8 hrs and (a) HO-1 mRNA (b) HO-1 protein levels were determined qRT-PCR and western blot analysis, respectively and (c) Data analysis was performed using total lab software by measuring the integrated band densities following background subtraction. The results are expressed as Mean±SEM of three independent experiments. Statistical significance is based on the difference when compared with 0 μM-treated cells (*p<0.05, ***p<0.001) |
In contrast to NF-κB p65 phosphorylation and Iκ-Bα degradation, no significant changes were found in the phosphorylation and total protein levels of MAPKs in RD4-treated cells compared with LPS-treated cells (Fig. 4c-d). These results indicated the potential role of NF-κB in the suppression of inflammatory mediators including NO and pro-inflammatory cytokines by RD4.
Effect of RD4 on HO-1 expression in RAW264.7 cells: We finally investigated whether the RD4 stimulated the induction of HO-1 in RAW264.7 macrophages because HO-1 plays an important role in the suppression of inflammation. Cells were challenged with RD4 and the levels of HO-1 mRNA were quantified by qRT-PCR. A dramatic and significant induction of HO-1 mRNA occurred in cells treated with 5 and 20 μM of RD4 (Fig. 5a). Next, we evaluated the protein levels of HO-1. As expected, RD4 dramatically induced HO-1 protein expression in cells treated with 5 and 20 μM concentrations (Fig. 5b-c). The relative protein expression level of HO-1 was 1 in RAW264.7 cells stimulated with LPS but increased to 10.9±0.67 for 20 μM RD4 concentrations. Altogether, these results suggest that HO-1 mediates the suppression of inflammatory responses.
DISCUSSION
The major findings of the present study are: (1) RD4 inhibited the expression of inflammatory mediators such as NO and pro-inflammatory cytokines, (2) RD4 suppressed NF-κB signalling pathways and (3) RD4 induced HO-1 expression. Thus, it can be concluded that RD4, a novel rugchalcone derivative, can significantly attenuate the inflammatory response in LPS-stimulated RAW264.7 cells.
Natural products, including phytochemicals, have long contributed to the development of modern therapeutics20,21. In particular, many studies have shown that certain phytochemicals can regulate the expression of various inflammatory mediators such as iNOS, pro-inflammatory cytokines and transcription factors such as NF-κB8,10,22,23. We believe ongoing efforts will provide new insights into the anti-inflammatory activity of phytochemicals and eventually result in the development of new types of anti-inflammatory agents. As part of this belief, we continue to synthesize phytochemicals and their derivatives and verify their anti-inflammatory activity. One of our previous studies demonstrated an efficient strategy to synthesize rugchalcone and its derivatives as well as their ability to inhibit NO production19. Based on this previous study, we investigated the anti-inflammatory activity and the detailed mechanism of RD4 in the present study.
NO acts as a cytotoxic agent in the pathophysiology of specific inflammatory diseases. The synthesis of NO by iNOS in activated macrophages may contribute to the pathology of various acute and chronic inflammatory diseases24,25. It is well known that the expression of iNOS plays a crucial role in stimulated macrophages, via the synthesis of NO24,25. Besides, pro-inflammatory cytokines, including IL-1β, IL-6 and TNF-α, play an essential role in promoting and exacerbating the inflammatory response3-5,7. Our data demonstrated that RD4 significantly inhibited the production of NO in LPS-stimulated RAW264.7 cells at non-cytotoxic concentrations. The reduced expression of iNOS mRNA and protein by RD4 treatment may result in the inhibition of LPS-induced NO synthesis by RD4. Besides, RD4 significantly inhibited the LPS-induced synthesis of IL-1β, IL-6 and TNF-α in LPS-stimulated macrophages, in a dose-dependent manner. These results indicate that RD4 regulates inflammation by inhibiting the synthesis of NO or pro-inflammatory cytokines.
NF-κB is one of the most widely known transcription factors in many cellular systems, where inflammatory stimuli contribute to NF-κB activation, followed by increased release of inflammatory mediators and cytokines8,10,12. In the absence of inflammatory stimuli, Iκ-Bα maintains NF-κB in the cytoplasm by concealing the nuclear localization sequences on NF-κB subunits. The phosphorylation of Iκ-B triggers proteasomal degradation of Iκ-B, increasing the nuclear localization of NF-κB, which activates various pro-inflammatory genes8,10,12. Our results revealed that exposure of RAW264.7 cells to LPS induced the degradation of Iκ-Bα and phosphorylation of NF-κB. However, RD4 treatment markedly reduced the phosphorylation of NF-κB and induced Iκ-Bα degradation. These findings suggest that RD4 treatment interferes with NF-κB activation by overcoming phospho-NF-κB levels and degradation of Iκ-B in LPS-stimulated RAW264.7 cells.
HO-1-knockout mice demonstrated the essential role of HO-1 in the anti-inflammatory effect. Compared with wild-type mice, HO-1-knockout mice were characterized by progressive chronic inflammation26. Increased proinflammatory cytokine levels were observed in peritoneal macrophages derived from HO-1-knockout mice compared with wild-type mice26. Similar phenomena have been found in humans, characterized by a pro-inflammatory state even in the case of human HO-1 deficiency27. Many recent studies suggest that naturally occurring phytochemicals play an anti-inflammatory role mediated via HO-1 expression16,17,28. Hence, we investigated whether RD4 induced HO-1 expression. Meanwhile, drugs that could strongly induce HO-1 might be beneficial for patients with inflammatory diseases. Taken together, the significantly increased HO-1 expression induced by RD4 treatment might contribute to the suppression of LPS-induced inflammatory responses.
CONCLUSION
In conclusion, we report for the first time that RD4 has anti-inflammatory activity in LPS-induced RAW264.7 macrophages and its possible anti-inflammatory mechanism. These effects were accompanied by blocking NF-κB activation and up regulating HO-1 expression in RAW264.7 cells. These findings shed a light on the possibility of the clinical application of RD4 in the treatment of inflammation-related diseases.
SIGNIFICANCE STATEMENT
This study showed that RD4 effectively inhibits the LPS-induced synthesis of inflammatory mediators including NO and pro-inflammatory cytokines via suppression of NF-κB in RAW264.7 macrophages without causing cytotoxicity. Besides, the induction of HO-1 might contribute to the anti-inflammatory activity of RD4. These results suggest that RD4 is a potential therapeutic agent against inflammatory disease.
ACKNOWLEDGMENT
This study was supported by research grants from Daegu Catholic University in 2020.
REFERENCES
- Medzhitov, R., 2008. Origin and physiological roles of inflammation. Nature, 454: 428-435.
CrossRefDirect Link - Stewart, A.G. and P.M. Beart, 2016. Inflammation: Maladies, models, mechanisms and molecules. Br. J. Pharmacol., 173: 631-634.
CrossRefDirect Link - Gatta, V., G. Mengod, M. Reale and A.M. Tata, 2020. Possible correlation between cholinergic system alterations and neuro/inflammation in multiple sclerosis. Biomedicines, Vol. 8.
CrossRefDirect Link - Samoilă, I., S. Dinescu and M. Costache, 2020. Interplay between cellular and molecular mechanisms underlying inflammatory bowel diseases development-A focus on ucerative colitis. Cells, Vol. 9.
CrossRefDirect Link - Sharma, V., R.K. Tiwari, S.S. Shukla and R.K. Pandey, 2020. Current and future molecular mechanism in inflammation and arthritis. J. Pharmacopuncture, 23: 54-61.
CrossRefDirect Link - Fujiwara, N. and K. Kobayashi, 2005. Macrophages in inflammation. Curr. Drug Targets-Inflam. Allergy, 4: 281-286.
Direct Link - Watanabe, S., M. Alexander, A.V. Misharin and G.R.S. Budinger, 2019. The role of macrophages in the resolution of inflammation. J. Clin. Invest., 129: 2619-2628.
CrossRefDirect Link - Gao, H., N. Kang, C. Hu, Z. Zhang, Q. Xu, Y. Liu and S. Yang, 2020. Ginsenoside Rb1 exerts anti-inflammatory effects in vitro and in vivo by modulating toll-like receptor 4 dimerization and NF-κB/MAPKs signaling pathways. Phytomedicine, Vol. 69.
CrossRefDirect Link - Yeung, Y.T., F. Aziz, A. Guerrero-Castilla and S. Arguelles, 2018. Signaling pathways in inflammation and anti-inflammatory therapies. Curr. Pharm. Des., 24: 1449-1484.
CrossRefDirect Link - Alam, M.B., M.K. Ju, Y.G. Kwon and S.H. Lee, 2019. Protopine attenuates inflammation stimulated by carrageenan and LPS via the MAPK/NF-κB pathway. Food Chem. Toxicol., Vol. 131.
CrossRefDirect Link - Mitchell, J.P. and R.J. Carmody, 2018. NF-κB and the transcriptional control of inflammation. Int. Rev. Cell Mol. Biol., 335: 41-84.
CrossRefDirect Link - Mulero, M.C., T. Huxford and G. Ghosh, 2019. NF-κB, IκB, and IKK: Integral components of immune system signaling. Adv. Exp. Med. Biol., 1172: 207-226.
CrossRefDirect Link - Nam, N.H., 2006. Naturally occurring NF-κB inhibitors. Mini Rev. Med. Chem., 6: 945-951.
CrossRefDirect Link - Drummond, G.S., J. Baum, M. Greenberg, D. Lewis and N.G. Abraham, 2019. HO-1 overexpression and underexpression: Clinical implications. Arch. Biochem. Biophys., Vol. 673.
CrossRefDirect Link - Motterlini, R. and R. Foresti, 2014. Heme oxygenase-1 as a target for drug discovery. Antioxid. Redox Signal., 20: 1810-1826.
CrossRefDirect Link - Kaulmann, A. and T. Bohn, 2016. Bioactivity of polyphenols: Preventive and adjuvant strategies toward reducing inflammatory bowel diseases-promises, perspectives and pitfalls. Oxid. Med. Cell. Longev., Vol. 2016.
CrossRefDirect Link - El-Achkar, G.A., M.F. Mrad, C.A. Mouawad, B. Badran and A.A. Jaffa et al., 2019. Heme oxygenase-1—Dependent anti-inflammatory effects of atorvastatin in zymosan-injected subcutaneous air pouch in mice. PLoS ONE, Vol. 14.
CrossRefDirect Link - Du, G., J.M. Han, W.S. Kong, W. Zhao and H.Y. Yang et al., 2013. Chalcones from the flowers of Rosa rugosa and their anti-tobacco mosaic virus activities. Bull. Korean Chem. Soc., 34: 1263-1265.
CrossRefDirect Link - Seo, Y.H., K. Damodar, J.K. Kim and J.G. Jun, 2016. Synthesis and biological evaluation of 2-aroylbenzofurans, rugchalcones A, B and their derivatives as potent anti -inflammatory agents. Bioorg. Med. Chem. Lett., 26: 1521-1524.
CrossRefDirect Link - Mathur, S. and C. Hoskins, 2017. Drug development: Lessons from nature. Biomed. Rep., 6: 612-614.
CrossRefDirect Link - Thomford, N.E., D.A. Senthebane, A. Rowe, D. Munro, P. Seele, A. Maroyi and K. Dzobo, 2018. Natural products for drug discovery in the 21st century: Innovations for novel drug discovery. Int. J. Mol. Sci., Vol. 19, No. 6.
CrossRefDirect Link - Farzaei, M.H., A.K. Singh, R. Kumar, C.R. Croley and A.K. Pandey et al., 2019. Targeting inflammation by flavonoids: Novel therapeutic strategy for metabolic disorders. Int. J. Mol. Sci., Vol. 20.
CrossRefDirect Link - Yao, L.J., J. Jalil, A. Attiq, C.C. Hui and N.A. Zakaria, 2019. The medicinal uses, toxicities and anti-inflammatory activity of Polyalthia species (Annonaceae). J. Ethnopharmacol., 229: 303-325.
CrossRefDirect Link - Sobrevia, L., L. Ooi, S. Ryan and J.R. Steinert, 2016. Nitric oxide: A regulator of cellular function in health and disease. Oxid. Med. Cell. Longev., Vol. 2016.
CrossRefDirect Link - Minhas, R., Y. Bansal and G. Bansal, 2020. Inducible nitric oxide synthase inhibitors: A comprehensive update. Med. Res. Rev., 40: 823-855.
CrossRefDirect Link - Kapturczak, M.H., C. Wasserfall, T. Brusko, M. Campbell-Thompson, T.M. Ellis, M.A. Atkinson and A. Agarwal, 2004. Heme oxygenase-1 modulates early inflammatory responses: Evidence from the heme oxygenase-1-deficient mouse. Am. J. Pathol., 165: 1045-1053.
CrossRefDirect Link - Radhakrishnan, N., S.P. Yadav, A. Sachdeva, P.K. Pruthi and S. Sawhney et al., 2011. Human heme oxygenase-1 deficiency presenting with hemolysis, nephritis, and asplenia. J. Pediatr. Hematol. Oncol., 33: 74-78.
CrossRefDirect Link - Leung, W.S., M.L. Yang, S.S. Lee, C.W. Kuo and Y.C. Ho et al., 2017. Protective effect of zerumbone reduces lipopolysaccharide-induced acute lung injury via antioxidative enzymes and Nrf2/HO-1 pathway. Int. Immunopharmacol., 46: 194-200.
CrossRefDirect Link