
Hyo Jeong Lee
College of Pharmacy and Research Institute of Pharmaceutical Sciences, Gyeongsang National University, Jinju-Si, Gyeongsangnam-Do 52828, Republic of Korea
Minji Bang
Department of Neuroscience, School of Medicine and Center for Neuroscience Research, Konkuk University, Seoul 05029, Republic of Korea
Chan Young Shin
Department of Neuroscience, School of Medicine and Center for Neuroscience Research, Konkuk University, Seoul 05029, Republic of Korea
Yong-Chul Kim
School of Life Sciences, Gwangju Institute of Science and Technology, Gwangju 61005, Republic of Korea
Sun-Young Han
College of Pharmacy and Research Institute of Pharmaceutical Sciences, Gyeongsang National University, Jinju-Si, Gyeongsangnam-Do 52828, Republic of Korea
LiveDNA: 82.34609
International Journal of Pharmacology
Year: 2020 | Volume: 16 | Issue: 8 | Page No.: 562-567







ABSTRACT
Background and Objective: Cellular senescence manifests as cell cycle arrest accompanied by phenotypic changes such as deregulated metabolism and specialized secretory activity. Senotherapeutics, which exploits cellular senescence for therapeutic purposes, is evolving with the advent of pharmacological interventions such as senolytics and senostatics. LDD-2269 is a benzylidene hydrazine derivative with lysine (K)-specific demethylase 4A activity and its effect on cellular senescence was examined in this study. Materials and Methods: Cellular senescence was measured using senescence-associated β-galactosidase assay. Western blot and quantitative RT-PCR were employed for the measurement of p21 and IL-8 expression. Results: Treatment with LDD-2269 induced senescence in U87MG glioblastoma cells and primary rat astrocytes. LDD-2269 upregulated the gene expression of p21 and IL-8, which is consistent with the senescent phenotype induction. LDD-2269 also augmented growth inhibition by temozolomide. Conclusion: The results indicate that LDD-2269 can be utilized in cancer senotherapeutics. The combination of LDD-2269 with other senolytics is a promising senotherapeutic strategy in cancer treatment.
PDF Abstract XML References Citation
Copyright: © 2020. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Hyo Jeong Lee, Minji Bang, Chan Young Shin, Yong-Chul Kim and Sun-Young Han, 2020. Effects of LDD-2269 on the Senescence of Glioblastoma Cells. International Journal of Pharmacology, 16: 562-567.
DOI: 10.3923/ijp.2020.562.567
URL: https://scialert.net/abstract/?doi=ijp.2020.562.567
DOI: 10.3923/ijp.2020.562.567
URL: https://scialert.net/abstract/?doi=ijp.2020.562.567
INTRODUCTION
Cellular senescence is characterized by permanent growth arrest accompanied by complex phenotypic changes1. Cell cycle arrest, deregulated metabolism, macromolecular damage and secretory phenotype are hallmarks of senescent cells2. Senescent cells not only contribute to natural aging but also cause age-related diseases3. Cancer is an age-related disease and cellular senescence is closely related in several ways. Senescence-Associated Secretory Phenotype (SASP), a hallmark of senescence, is known to function as both a tumor promoter and a suppressor2. Several anticancer agents, including kinase modulators, induce senescence in cancer cells4,5. Besides, in oncogene-induced senescence, oncogenes such as RAS function as cellular stress to induce premature senescence6.
Senolysis indicates selective elimination of senescent cells6. Novel treatment strategies to overcome current treatment problems can be developed using senolysis. Senescence of cancer cells is induced and senescent cells are eliminated by senolytic agents. Combination treatment with senescent-inducing agents and senolytic agents can be an effective approach for cancer therapy. By selectively clearing only senescent cells, cancer cells can be eliminated without affecting normal cells. Thus, it is important to understand the effect of anticancer agents on senescence.
LDD-2269 is a lysine (K)-specific demethylase 4A (KDM4A) inhibitor that inhibits the growth of colon cancer cells1. KDM4A is an enzyme that removes methyl residues from the trimethylated form of histone 3 protein at the lysine 9 and 36 residues (H3K9 and H3K36)7.
Histone alterations, a mechanism of epigenetic regulation, are associated with cellular senescence and histone demethylases are reported to be involved in replicative senescence8. For example, the KDM6B protein (also known as JMJD3), regulates cellular senescence and aging via signaling processes such as p53 and the INK box network9.
Given the relationship between histone methylation and senescence, the effect of the KDM4A inhibitor LDD-2269 on cellular senescence was examined in this study. Furthermore, the genes involved in cellular senescence were studied. The cytotoxic effects of LDD-2269 were measured to investigate its potential as an anti-cancer agent.
MATERIALS AND METHODS
Study area: The study was carried out at College of Pharmacy, Gyeongsang National University, School of Medicine Konkuk University and School of Life Science, Gwangju Institute of Science and Technology, Korea from March 20, 2017-February 13, 2020
Cell culture: The human U87MG glioblastoma cell line was purchased from the Korean Cell Line Bank (Seoul, Korea). The cells were cultured in MEM (Minimum Essential Medium, 11095-080, Gibco, Thermo Fisher Scientific, Waltham, MA, USA) supplemented with 10% fetal bovine serum and 1% penicillin/streptomycin at 37°C under humidified air containing 5% CO2. Rat primary astrocytes were isolated from the frontal cortices of a 2 days old Sprague-Dawley rat.
Cells were cultured in DMEM/F12 media supplemented with 10% FBS. The purity of astrocytes was confirmed by immunostaining with a Glial Fibrillary Acidic Protein (GFAP) antibody.
Immunoblot analysis: Cells were analyzed in SDS lysis buffer (12 mM Tris-Cl, pH 6.8, 5% glycerol and 0.4% SDS) and the protein concentrations were measured with the SMART BCA Protein Assay kit (iNtRON Biotechnology, Korea). The proteins were resolved by SDS-polyacrylamide gel electrophoresis, transferred to PVDF membranes (Millipore, Billerica, MA, USA) and incubated overnight with the appropriate antibodies. The antibody against p21 (rabbit monoclonal IgG; #2947, 1:1,000) was purchased from Cell Signaling Technology Inc. (Danvers, MA, USA) and β-actin antibody (mouse monoclonal IgG; #A5441, 1:5,000) from Sigma-Aldrich (St. Louis, MO, USA). Goat anti-rabbit IgG (#111-035-003; 1;5,000) and anti-mouse IgG (#115-035-033; 1:5,000) secondary antibodies were obtained from Jackson ImmunoResearch Laboratories, Inc., (West Grove, PA, USA).
Analysis of senescence-associated β-galactosidase (SA-βgal): Cells were plated in 6-well plates (1.3×106 cells/well) and incubated overnight at 37°C. The next day, cells were treated with LDD-2269 (0, 0.1, 1, 10 and 100 μM). After 24 hrs, the cells were fixed and stained for β-galactosidase expression using the cell senescence assay kit (Cell Signaling Technology Inc., Danvers, MA, USA) according to the manufacturer’s protocol. The cells were washed with PBS and fixed in 4% formaldehyde for 15 min at RT. Then, they have washed again with PBS and incubated with the SA-βgal staining solution at 37°C for 48 hrs. Images were acquired using a Zeiss Axio Observer Z1 inverted fluorescence microscope (Zeiss, Oberkochen, Germany). Five random fields of images were selected for counting β-galactosidase-positive cells.
Quantitative RT-PCR: Total RNA was extracted using the ReliaPrepTM RNA Cell Miniprep System (Promega, Madison, WI, USA) according to the manufacturer’s instructions. The first-strand cDNA was obtained from 1 μg of purified RNA using SuperScriptTM III Reverse Transcriptase (Invitrogen, Carlsbad, CA, USA). The qPCR was performed using BrightGreen qPCR Master Mix (Applied Biological Materials, Richmond, BC, Canada) at 95°C for 10 min followed by 60 cycles at 95°C for 15 sec and 55-60°C for 60 sec and CFX Manager software program (Bio-Rad Laboratories, Hercules, CA, USA). Data are represented as relative amounts of mRNA normalized to GAPDH control.
The primer sequences used in this study were as follows: p21 forward 5-CAGACCAGCATGACAGATTTC-3' and reverse 5-TTAGGGCTTCCTCTTGGAGA-3 (60°C), IL-8 forward 5'-ATGACTTCCAAGCTGGCCGTGGCT-3' and reverse 5'-TCTCAGCCCTCTTCAAAAACTTCTC-3' (60°C), GAPDH forward 5-TGAACGGGA AGCTCACTGG-3 and reverse 5-GGTCCACCACTGACACGTTG-3 (60°C).
Cell viability assay: Cell viability was assessed using a tetrazolium-based cell viability assay (EZ-Cytox, DaeilLab, Korea). About 2,000 cells/well were plated in a 96-well plate in 100 μL of the medium. The next day, cells were treated with compounds along with dimethyl sulfoxide (DMSO) as a negative control and further incubated for 72 hrs. Then, 15 μL of EZ-Cytox kit reagent was added to each well of the 96-well plate and incubated at 37 in a humidified CO2 incubator for 4 hrs. After incubation, the Optical Density (OD) was measured at a wavelength of 450 nm with a Victor X5 Multilabel Plate Reader (Perkin Elmer, Waltham, MA, USA).
RESULTS
LDD-2269 induces cellular senescence in glioblastoma cell lines: The benzylidene hydrazine derivative LDD-2269 was characterized as a KDM4A inhibitor in a previous report by our laboratory. In this study, LDD-2269 was tested for its senescence-inducing effects. The SA-βgal assay was carried out in the U87MG glioblastoma cell line. As shown in Fig. 1a, LDD-2269 induced SA-βgal activity. Cells exhibiting positive SA-βgal activity were counted and the percentage of SA-βgal-positive cells was calculated (Fig. 1a). A five-fold increase in SA-βgal-positive cells was observed with 100 μM of LDD-2269. Senescence was also induced in rat primary astrocytes by LDD-2269, confirming the results obtained with U87MG cells (Fig. 1b).
LDD-2269 induced senescence is mediated by p21 and IL-8: To confirm senescence induction, gene expression related to senescence was investigated. U87MG cells were treated with LDD-2269 and the expression levels of p21 and IL-8 were determined by western blot and qRT-PCR analyses. Western blot results for p21 after 48 hrs of treatment with LDD-2269 were shown in Fig. 2a. At a concentration of 100 μM, LDD-2269 highly upregulated p21 expression (Fig. 2a-b). The results of p21 qRT-PCR, the increment of p21 mRNA at 100 μM LDD-2269 treatment, are consistent with the western blot results. The mRNA level of IL-8 was also higher in the 100 μM LDD-2269 treatment group than in the control group (Fig. 2c).
Treatment with LDD-2269 augmented growth suppression mediated by temozolomide: The senescence-inducing effects of LDD-2269 on the growth of U87MG cells in the presence of temozolomide were investigated. Temozolomide is an alkylating agent used for the treatment of gliomas. Cells were treated with various concentrations of temozolomide with or without 50 μM of LDD-2269. The combination of temozolomide with LDD-2269 resulted in higher inhibition of cell growth than temozolomide alone (Fig. 3). At 500 μM of temozolomide, the percentage of inhibition of cell growth was 28.6% (% cell growth 71.4%). When cells were treated with both temozolomide and LDD-2269, 63.5% inhibition was observed. These results indicate the potential of LDD-2269 in combination with temozolomide to treat glioblastoma.
DISCUSSION
In this study, the LDD-2269 compound, previously reported as KDM4A inhibitor, induced cellular senescence in glioblastoma cells and primary rat astrocytes. Small molecules that modulate cellular senescence can be categorized into senolytics and senostatics10. While senolytics induce senescent cell clearance, senostatics inhibit senescence-associated bystander signaling.
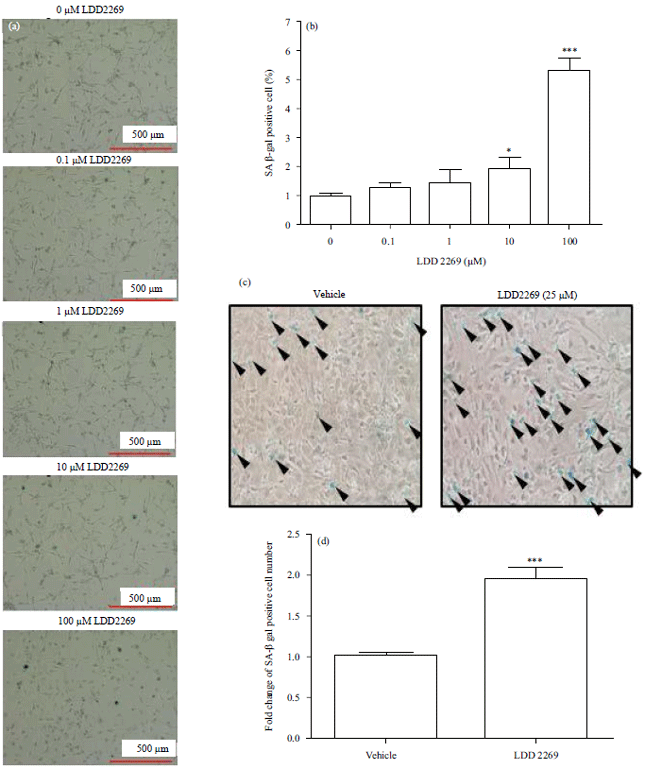 | |
| Fig. 1(a-d): | Induction of senescence by LDD-2269 |
Cells were treated with LDD-2269 at at the indicated concentration for 24 hrs. The SA-βgal assay was conducted and photographed. (a) Images of U87MG cells (b) Fold changes of SA-betagal-positive U87MG cells (c) Images of rat primary astrocytes (d) Fold changes of SA-βgal-positive rat primary astrocytes | |
Several senolytics have been identified for therapeutic purposes, which is termed as senotherapy. Navitoclax and ABT-737, Bcl-2 inhibitors with pro-apoptotic function, exhibit senolytic activity. The combination of dasatinib and quercetin is also a candidate for senotherapy6. Heat shock protein 90 (Hsp90) inhibitors, 17-AAG and geldanamycin, are pharmacological modulators effective in selectively eliminating senescent cells11.
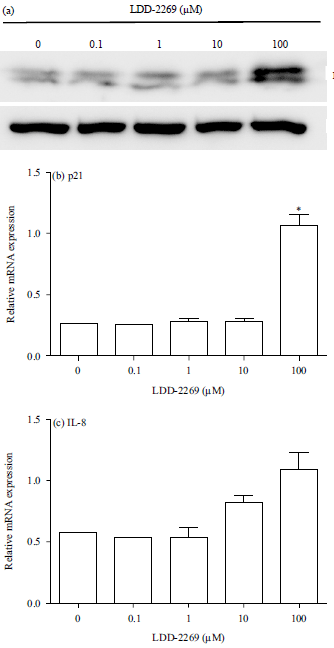 | |
| Fig. 2(a-c): | Effects of LDD-2269 on p21 and IL-8 levels |
(a) U87MG cells were treated with LDD-2269 at the indicated concentration for 48 hrs and cell lysates were subjected to immunoblot analyses, (b) U87MG cells were treated with the indicated concentration of LDD-2269 compound for 8 hrs and RNA extracts were subjected to qRT-PCR experiments using the p21 primers and (c) U87MG cells were treated with the indicated concentration of LDD-2269 for 24 hrs and qRT-PCR experiments were performed with RNA extracts using the IL-8 primers | |
LDD-2269 described in this study has the potential for use in combination with senolytics. Senescence induction by LDD-2269 (Fig. 1), followed by senescent cell clearance by senolytics, can be a powerful approach for the treatment of glioblastoma.
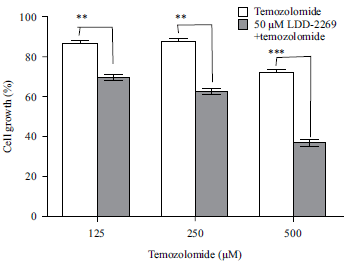 | |
| Fig. 3: | Effect of LDD-2269 on cell growth in the presence of temozolomide |
U87MG cells were treated with the indicated concentration of temozolomide with or without 50 μM of LDD-2269 for 72 hrs. Cell viability was measured using EZ-Cytox | |
The changes in the expression of p21 and IL-8 are shown in Fig. 2. The role of p21 protein, a CDK2 inhibitor and a master regulator of the cell cycle, in cellular senescence, is well known by Gorgoulis et al.2. LDD-2269 upregulated the expression of p21 and mRNA at 100 μM, explaining the senescent phenotype at the same concentration. IL-8, a chemokine and one of the SASP components, presumably recruits inflammatory cells6. The IL-8 mRNA level was also increased by LDD-2269, consistent with the senescent phenotype.
Our understanding of cellular senescence is expanding with extensive research effort and the development of chemical tools. Pharmacological modulators such as LDD-2269 can be a useful tool for senescence research. Furthermore, the combination of LDD-2269 with other senolytics is a promising senotherapeutic strategy in cancer treatment.
CONCLUSION
These results indicate that LDD-2269 can be utilized in cancer senotherapeutics. The combination of LDD-2269 with other senolytics is a promising senotherapeutic strategy in cancer treatment.
SIGNIFICANCE STATEMENT
This study discovers the senescence-inducing effects of KDM4A inhibitor, potentially implicating the importance of KDM4A in cellular senescence. This study will help the researchers to find a novel senotherapeutic approach in combination with other anticancer agents. Novel senotherapeutics can provide new paradigms for cancer pharmacotherapy.
ACKNOWLEDGMENT
This work was supported by a grant from the National Research Foundation (NRF, 2018R1A2B6002081) funded by the government of Korea (MEST).
REFERENCES
- Alexander, P.B., L. Yuan, P. Yang, T. Sun and R. Chen et al., 2015. EGF promotes mammalian cell growth by suppressing cellular senescence. Cell Res., 25: 135-138.
CrossRefDirect Link - Gorgoulis, V., P.D. Adams, A. Alimonti, D.C. Bennett and O. Bischof et al., 2019. Cellular senescence: Defining a path forward. Cell, 179: 813-827.
CrossRefDirect Link - Childs, B.G., M. Durik, D.J. Baker and J.M. van Deursen, 2015. Cellular senescence in aging and age-related disease: From mechanisms to therapy. Nat. Med., 21: 1424-1435.
CrossRefDirect Link - Petrova, N.V., A.K. Velichko, S.V. Razin and O.L. Kantidze, 2016. Small molecule compounds that induce cellular senescence. Aging Cell, 15: 999-1017.
CrossRefDirect Link - Lee, C., J. Baek and S.Y. Han, 2017. The role of kinase modulators in cellular senescence for use in cancer treatment. Molecules, Vol. 22.
CrossRefDirect Link - Childs, B.G., M. Gluscevic, D.J. Baker, R.M. Laberge, D. Marquess, J. Dananberg and J.M. van Deursen, 2017. Senescent cells: An emerging target for diseases of ageing. Nat. Rev. Drug Discov., 16: 718-735.
CrossRefDirect Link - Klose, R.J., K. Yamane, Y. Bae, D. Zhang and H. Erdjument-Bromage et al., 2006. The transcriptional repressor JHDM3A demethylates trimethyl histone H3 lysine 9 and lysine 36. Nat., 442: 312-316.
CrossRefDirect Link - Cheng, L.Q., Z.Q. Zhang, H.Z. Chen and D.P. Liu, 2017. Epigenetic regulation in cell senescence. J. Mol. Med., 95: 1257-1268.
CrossRefDirect Link - Salminen, A., K. Kaarniranta, M. Hiltunen and A. Kauppinen, 2014. Histone demethylase Jumonji D3 (JMJD3/KDM6B) at the nexus of epigenetic regulation of inflammation and the aging process. J. Mol. Med., 92: 1035-1043.
CrossRefDirect Link - Short, S., E. Fielder, S. Miwa and T. von Zglinicki, 2019. Senolytics and senostatics as adjuvant tumour therapy. EBioMedicine, 41: 683-692.
CrossRefDirect Link - Kim, E.C. and J.R. Kim, 2019. Senotherapeutics: Emerging strategy for healthy aging and age-related disease. BMB Rep., 52: 47-55.
CrossRefDirect Link