
Seong-Hui Eo
Department of Biological Sciences, College of Natural Sciences, Kongju National University, Gongju 32588, Republic of Korea
Seon-Mi Yu
Department of Biological Sciences, College of Natural Sciences, Kongju National University, Gongju 32588, Republic of Korea
Yohan Han
Department of Biological Sciences, College of Natural Sciences, Kongju National University, Gongju 32588, Republic of Korea
Jung Han
Department of Biological Sciences, College of Natural Sciences, Kongju National University, Gongju 32588, Republic of Korea
Su Min Kim
Department of Biological Sciences, College of Natural Sciences, Kongju National University, Gongju 32588, Republic of Korea
Dan-Bi Kim
BIOCEN. Corporation Limited, 109, Deokjin-gu, jeonju 54853, Republic of Korea
Bong Kyun Jeon
BIOCEN. Corporation Limited, 109, Deokjin-gu, jeonju 54853, Republic of Korea
Won Kil Lee
Lee’s Biotech Corporation Limited, 125, Yuseong-gu, Daejeon 34141, Republic of Korea
Song Ja Kim
Department of Biological Sciences, College of Natural Sciences, Kongju National University, Gongju 32588, Republic of Korea
LiveDNA: 82.28838
International Journal of Pharmacology
Year: 2020 | Volume: 16 | Issue: 6 | Page No.: 479-491







ABSTRACT
Background and Objective: Thymoquinone (TQ) is one of the most potent constituents in Nigella Sativa and has many medicinal effects. The Rural Development Administration of Korea that is using genetic engineering to combine the properties of resveratrol and rice develops Iksan526. The present study was aimed to investigate the effects of thymoquinone (TQ) and Iksan526 callus extract (IS526) on A375 and B16F10 melanoma cell lines. Materials and Methods: A375 and B16F10 cells, cultured in vitro, were treated with TQ or IS526 and their effects on cell proliferation, tyrosinase activity, Reactive Oxygen Species generation (ROS), and expression levels of Cyclooxygenase (COX)-2, phosphorylated p38, and phosphorylated extracellular signal-regulated kinase (pERK) were evaluated with the MTT assay, tyrosinase activity assay, ROS assay, and western blot analysis, respectively. Results: In comparison with cells treated with either TQ or IS526, those treated with both TQ and IS526 showed a significant decrease in cell proliferation. TQ-induced cyclooxygenase-2 (COX-2) was inhibited by IS526 and TQ inhibited IS526-induced ROS production via MAPKs pathway in A375. Also, co-treatment with TQ and IS562 decreased tyrosinase expression via MAPKs pathway in A375. Combination with TQ and IS526 suppressed COX-2 and tyrosinase expression via the p38 kinase pathway in B16F10 cells. Conclusion: Taken together, results demonstrated that combination with TQ and IS526 can suppress cell growth and regulates COX-2, tyrosinase and ROS in melanoma cells, indicating its application potential for melanoma therapy.
PDF Abstract XML References Citation
Received: April 20, 2020;
Accepted: August 18, 2020;
Published: September 15, 2020
Copyright: © 2020. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Seong-Hui Eo, Seon-Mi Yu, Yohan Han, Jung Han, Su Min Kim, Dan-Bi Kim, Bong Kyun Jeon, Won Kil Lee and Song Ja Kim, 2020. Effects of Thymoquinone and Iksan 526 callus Extract on B16F10 and A375 Cell Lines. International Journal of Pharmacology, 16: 479-491.
DOI: 10.3923/ijp.2020.479.491
URL: https://scialert.net/abstract/?doi=ijp.2020.479.491
DOI: 10.3923/ijp.2020.479.491
URL: https://scialert.net/abstract/?doi=ijp.2020.479.491
INTRODUCTION
Malignant melanoma is the most aggressive and lethal form of skin cancer and accounts for less than 5% of all skin tumors; however, it is responsible for over 80% of skin cancer-related deaths1,2. Melanoma is curable through surgical removal during the early stages but is characterized by high metastatic potential, rapid growth and high lethality at advanced stages, making surgery an unlikely option in most cases. In addition to surgery, the major interventions for the treatment of malignant melanoma include chemotherapy, radiotherapy, and immunotherapy. These therapies are usually expensive and associated with severe adverse effects3-5. The World Health Organization report indicated 48,000 melanoma-related deaths and about 132,000 new cases of melanoma worldwide each year6. Furthermore, the incidence and mortality rates of melanoma have increased worldwide over the past decades, especially in young people. Ultraviolet (UV) radiation is a risk factor for the development of skin cancer, including melanoma7,8.
Melanin is a major factor that affects the skin, hair, and eye color and acts as a defense system against the harmful effects of UV-induced skin damage. However, abnormal melanin production may lead to several diseases and all types of skin cancers9. Melanogenesis is mediated by several enzymes such as tyrosinase, tyrosinase-related protein (TYRP)-1 and TYRP-2 and their transcription factors, Microphthalmia-associated Transcription Factor (MITF) and diverse signal transduction pathways10,11.
Cyclooxygenase (COX) is known as an important inflammatory mediator and serves as a rate-limiting enzyme in the conversion of arachidonic acid to prostaglandins12. There are two isozymes of COX, COX-1 and COX-2. In general, COX-1 is constitutively expressed and regarded as a housekeeping gene, whereas COX-2 is an inducible gene and its expression increases in response to various stimuli in different tissues, including the epidermis12,13. Previous studies have shown considerably elevated levels of COX-2 expression in various types of human cancers, including malignant melanoma. Therefore, COX-2 expression is an important tumor-promoting factor and several studies suggest the strong correlation between COX-2 expression and development and progression of disease14-16.
Reactive Oxygen Species (ROS) are small molecules that react with cellular components and cause structural and functional damage to cells and tissues. Cancer cells, including melanoma cells, exhibit high levels of ROS15,17,18. The generation of ROS in cells causes oxidative stress and is deemed as an adverse event for promoting tumorigenesis and tumor progression. Cutaneous ROS production may be induced by UVB radiation15,19. The increase in the ROS level is known to enhance UVB-induced skin carcinogenesis. Furthermore, ROS generation in cells may induce or mediate the activation of 73 mitogen-activated protein kinase (MAPK) signaling pathways such as p38 kinase, c-Jun N-74 terminal kinase (JNK) and extracellular signal-regulated kinase (ERK-1/-2) pathways. 75MAPK pathway activation is known to regulate COX-2 expression20-22.
Thymoquinone (TQ) is a component of the volatile oil extracted from the medical plant Nigella sativa. TQ is a promising compound for the treatment of various diseases such as cancer. It has been shown to inhibit cellular proliferation and induce apoptosis in many types of cancer cell lines such as breast, brain and pancreatic cells23,24. Several studies have documented the anti-inflammatory and antioxidant effects of TQ both in vitro and in vivo23-26. Iksan526 is resveratrol-enriched transgenic rice that overexpresses the stilbene synthase gene isolated from peanut (Arachis hypogaea var. Palkwang). This rice was developed by the Rural Development Administration of Korea. It is suggested that the resveratrol-enriched transgenic rice may exhibit biological activities similar to or higher than those of resveratrol27,28. Experimental data show that resveratrol-enriched transgenic rice may treat obesity in high-fat diet-fed mice and downregulate melanin synthesis in UVB-induced epidermal skin tissue of guinea pigs29-31.
Resveratrol (3,4 ,5-trihydroxy-trans-stilbene) is present in many berries, peanuts, grape, soy and red wine29-31. Previous studies have shown that resveratrol has been to possess a numerous spectrum of pharmacologic properties including anti-inflammatory, anti-oxidant and anti-tumorigenic effects against32. Also, resveratrol has been used for the treatment of various allergic disorders, such as dermatitis and bronchitis, as well as skin aging and other conditions33-35. However, there are several impediments to applying this promising agent as a treatment, such as its poor bioavailability and fast metabolism35.
In the last few decades, trends have been shifted to combination therapy from relying on monotherapies. This trend is enhanced efficacy compared to the mono-therapy approach because it is based on the fact that cancer cells have multiple pathways for survival including enhanced proliferation, angiogenesis stimulation, apoptosis evasion and invasion and metastases36,37. Therefore, the present study aimed to evaluate the effect of the novel combination of plant derived-natural products comprising TQ and Iksan526 callus extract against A375 and B16F10 melanoma cell line.
MATERIALS AND METHODS
Study area: The study was carried out at the Department of Biological Sciences, College of Natural Sciences, Kongju National University cell signaling network Lab, the Republic of Korea from June 2017 to December 2018.
Chemical reagents and antibodies: Thymoquinone (TQ), L-Dopa, Phenylmethanesulfonyl Fluoride (PMSF) were obtained from Sigma-Aldrich (St. Louis, MO, USA), Iksan526 callus extracts (IS526) purchased from BIOCEN (Jeonju, Republic of Korea).PD98059 (PD) was purchased from Calbiochem (San Diego, CA, USA) and, SB203580 (SB) was purchased from Biomol (Plymouth Meeting, PA, USA). The rabbit anti-pERK and pp38 were purchased from Cell Signaling Technology (Beverly, MA, USA). The rabbit anti-tyrosinase, mouse anti-GAPDH and goat anti-COX-2 were purchased from Santa Cruz Biotechnology Inc. (Santa Cruz, CA, USA). The anti-rabbit IgG was obtained from Sigma-Aldrich and anti-goat IgG obtained from Chemicon International (Billerica, MA, USA). The anti-mouse IgG was obtained from Enzo Life Sciences International (Farmingdale, NY, USA).
Cell culture: The A375 human melanoma cancer cell line and B16F10 mouse melanoma cells were purchased from the Korean Cell Line Bank (KCLB; Seoul, Republic of Korea). A375 and B16F10 cells were maintained in Dulbecco’s modified Eagle’s medium (DMEM; Gibco/Invitrogen, Carlsbad, CA) supplemented with 10% heat-inactivated Fetal bovine serum (FBS; Gibco/Invitrogen) and 50 μg mL–1 streptomycin (Sigma-Aldrich, St. Louis, MO, USA) and 50U mL–1 penicillin (Sigma-Aldrich) at 37 in a 5% CO2 and 95% air humidified incubator.
Treatment of A375 and B16F10 cells: Thymoquinone was dissolved in dimethyl sulfoxide (DMSO) and the final concentration of DMSO did not exceed 0.1% in the fresh medium. Iksan526 callus extracts were dissolved in distilled water. The cells were seeded into 35mm dishes at a density of 2×105 cells/well (B16F10) or 1.5×105 cells/well (A375) and incubated for 12 h to grow a monolayer. Cells were grown in 35 mm dish to ~75% confluence and treated with TQ (20 μM) and/or IS526 (40 nM) callus extracts for 24 h. PD 98059 (30 μM) and SB203580 (20 μM) was added 2 h before TQ and/or IS526 callus extracts to inhibit the phosphorylation of ERK-1/-2 and p38 kinase.
MTT assay: A375 and B16F10 cells proliferation of cultured cells were determined by MTT assay. Cells were seeded in 96-well plates at a density of 2×104 cells/well and cultured for 24 hrs before treating with TQ and/or IS526. Next, MTT reagent1 (5 mg mL–1 in PBS, 10 μL) was added to each well. Cells were incubated at 37°C for 4 h and 100 μL/well of MTT reagent 2 (10% SDS with 0.01N HCl) was added to dissolve the formazan crystals. After overnight incubation, the absorbance was measured at 600 nm with a spectrophotometer. The cell proliferation was calculated using the following formula:
![]()
where, T mean absorbance of TQ or IS526/C mean absorbance of control).
Western blot analysis: The western blotting assay was used for assessment of expressions of COX-2, pERK, pp38, tyrosinase and GAPDH in A375 and B16F10 cells. Cells were collected and washed with PBS. The total cell lysates were using cold radio immune precipitation assay buffer composed of 50mM Tris-HCl (pH 7.4), 150mM NaCl, 1% Nonidet P-40% and 0.1% Sodium Dodecyl Sulfate (SDS) and supplemented with protease and phosphatase inhibitors. Then the lysates were clarified by centrifugation (1,300 rpm, 10 min, 4°C) and collected. Subsequently, 30~μg of protein per sample were separated by 8% SDS-PAGE systems and transferred onto a nitrocellulose membrane (Whatman Schleicher and Schuell; Dachen, Germany). After blocking in 5% fat-free milk for 1h, the membranes were probed with diluted primary antibodies overnight at 4°C. The antibodies and dilution factors were as follows: COX-2 (1:1000), pERK (1:1000), pp38 (1:1000) and GAPDH (1:5000). Secondary antibodies conjugated with HRP were incubated for further 2hrs at room temperature. Proteins were detected by a chemiluminescence kit (BIOFACT CO., LTD, Daejeon, Republic of Korea) and bands were visualized using the LAS-4000 imaging system (FujiFilm, Tokyo, Japan). The bands were quantified by densitometric analysis using the ImageJ software package (National Institutes of Health, Bethesda, MD, USA).
Immunofluorescence staining: The expression of COX-2 and tyrosinase at the protein level in A375 was analyzed by Immunofluorescence microscopy. Cells were fixed with 3.5% paraformaldehyde in PBS for 15min at room temperature and permeabilized with 0.1% Triton X-100 in PBS for 15 min. The cells were then blocked with 5% skim milk to prevent non-specific reactions. Then, fixed cells were incubated with antibodies against tyrosinase (1:100) and COX-2 (1:100) for 1h at room temperature. Cells were washed three times with PBS and incubated with secondary antibodies (1:50) at room temperature for 1h. Fluorescence images were recorded using a BX51 fluorescence microscope (Olympus, Tokyo, Japan).
Cellular tyrosinase activity assay: A375 and B16F10 cells were treated with TQ and/or IS526 callus extracts for 24 hrs to determine tyrosinase activity. The treated cells were harvested, washed with PBS and lysed with lysis buffer (1% Triton X-100 in 0.1 M sodium phosphate buffer pH 6.8, containing protease inhibitor; PMSF) for 30 min. After centrifugation (13,000 rpm, 10 min), protein concentrations were determined using the BCA protein assay and adjustment of the concentration with lysis buffer. Cell lysate supernatants (each 100 μL) were placed in a well of a 96-well plate and mixed with 50 μL of L-DOPA (2 mg mL–1 in 0.1 M Na phosphate buffer) and 50 μL of lysis buffer. After incubation at 37°C for 1 h. Tyrosinase activity was determined by measuring sample absorbance at 475 nm. Cellular tyrosinase activities were determined as the ratio of tyrosinase content over total protein content.
ROS assay: The oxidation-sensitive dye dichlorodihydrofluorescein diacetate (DCFDA, Sigma-Aldrich) was used to monitor the production of intracellular ROS. Cells treatment with TQ and/or IS526 for 24 hrs and washed twice with PBS. The cells incubated at 37°C in DMEM without phenol red in the presence of 10 μM DCFDA for 30 min. Following treatment, the medium was removed, the chondrocytes were collected by centrifugation and fluorescence was measured on an Flx 8000 fluorometer (excitation, 485 nm/emission, 525 nm; Bio-Tek Instruments, Winooski, VT, USA).
Statistical analysis: Data were analyzed statistically by a one-way Analysis of Variance (ANOVA) followed by Tukey's post hoc test and all values are expressed as the Means±standard deviation (SD) of at least four independent experiments. Differences between groups were p values <0.05 were considered statistically significant.
RESULTS
Effect of the combined drugs on cell growth: To examine the possible combinatorial effect of TQ and IS526 against A375 and B16F10 cell proliferation, A375 and B16F10 cells were treated with TQ in the presence or absence of IS526. The results of the MTT assay showed that TQ and IS526 alone caused about 27% and 16% reduction in the proliferation of A375 cells after 24 hrs treatment, respectively. On the other hand, the combination of TQ and IS526 caused a reduction of 40% comparing with the control group in the proliferation of A375 cells. Also, a combination of TQ and IS526 caused a reduction of (18 and 29%) respectively compared with the TQ or IS526 (Fig. 1a). Moreover, when B16F10 cells were treated with the TQ or IS526 for 24 h, respectively, the proliferation was 65 and 75% compared to control-treated cells. Also, the combination of TQ and IS526 caused a reduction of 50% in the proliferation, a combination of TQ and IS526 caused a reduction of 23.08% and 33.33%, respectively compared with the TQ or IS526 (Fig. 1b).
TQ and IS526 regulates expression of Cyclooxygenase (COX) 2: The effects of TQ and/or IS526 on COX-2 expression are shown in Fig. 2. TQ induced the expression of COX-2, but cells treated with IS526 showed inhibited COX-2 expression as compared to control cells. IS526 reversed the increase in COX-2 expression induced by TQ and the difference was significant as compared with cells treated with TQ alone (Fig. 2a). However, the results showed that the treatment of IS526 or TQ suppressed the COX-2 expression in B16F10 cells. Also, the combination treatment induced a synergistic inhibitory effect on the COX-2 expression (Fig. 2b).
IS526 and TQ regulate COX-2 expression via ERK-1/-2 and p38 kinase pathways in A375 and B16F10 cells: To determine whether TQ and/or IS526 activate ERK-1/-2 and p38 kinase pathways in A375 and B16F10 cells, the expression levels of pERK and pp38 were detected by western blotting after 24 h treatment with different drugs. Treatment with TQ resulted in the activation of pERK, while IS526 treatment decreased pERK expression as compared with the control treatment. However, the pp38 protein level decreased, when treatment TQ or IS526 in both cells (Fig. 3). Also, IS526 inhibited the pERK expression induced by TQ and the pp38 protein level more decreased in A375. The difference was significant as compared with cells treated with TQ alone (Fig. 3a). When B16F10 cells were treated with combination TQ and IS526, it accelerated the reduction of pp38 and pERK expression compared with cells treated with either TQ or IS526 (Fig. 3b).
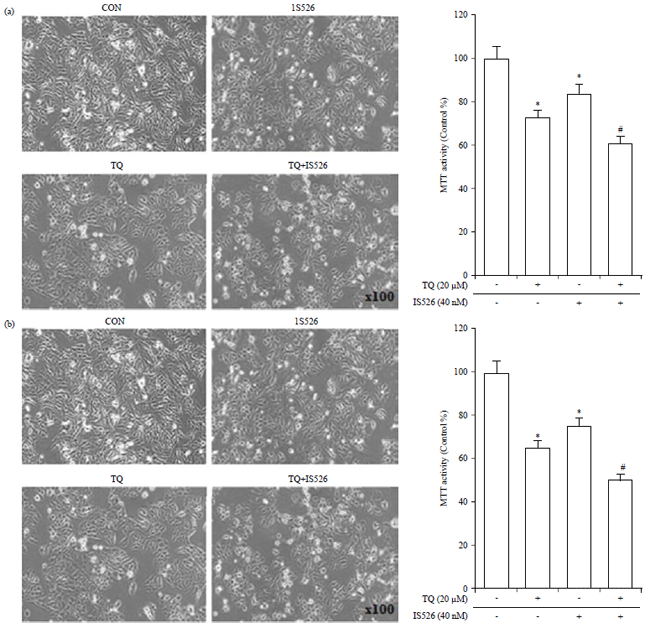 | |
| Fig. 1(a-b): | Synergistic growth inhibitory effect of the combination of thymoquinone (TQ) and IS526 |
| A375 and B16F10 cells were incubated with TQ, IS526, or both for 24 hrs, (a and b; left panel) Cells were viewed under a phase-contrast microscope and photographed (100× magnification), (a and b; right panel) Cell proliferation was measured by the MTT assay and the data represent growth inhibition as a percentage of control cells. The data represent a typical experiment, whereby similar results were obtained from three experiments, *p<0.05 as compared with untreated cells. #p<0.05 as compared with TQ or IS526 alone | |
Furthermore, investigated if the decreased cell proliferation and COX-2 expression during the combination treatment were induced through the activation of p38 kinase and ERK-1/-2. Therefore, to elucidate the association of these signaling systems with COX-2 expression, cell proliferation, tyrosinase activity due to TQ and IS526, the MAPKs signal transduction pathways were attenuated with their respective inhibitors, PD and SB. The inhibition of pp38 and pERK activity with SB or PD enhanced the suppressive effect of IS526 on TQ-induced COX-2 expression in A375 cells (Fig. 3a).
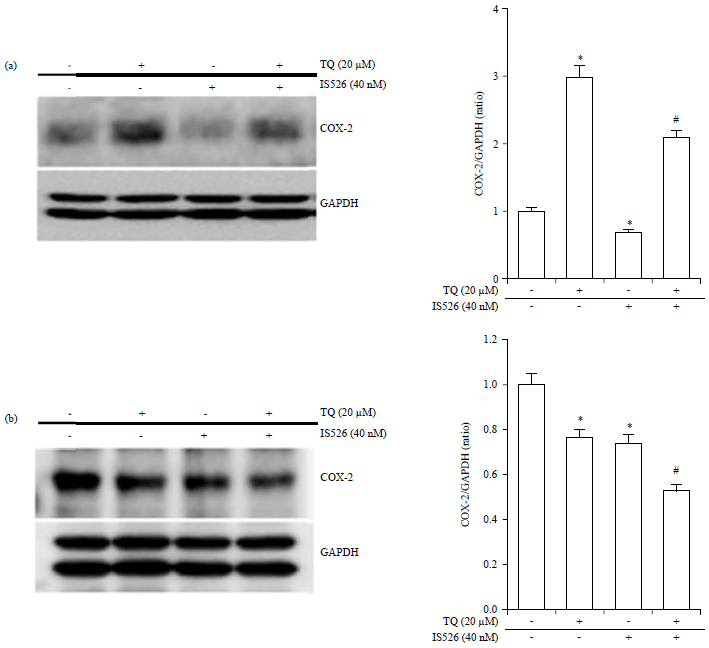 | |
| Fig. 2(a-b): | Effects of the combinatorial treatment of TQ and IS526 on the expression levels of COX-2 |
The cells were incubated with TQ, IS526, or both for 24 hrs, (a and b; left panel) The expression level of COX-2 and glyceraldehyde-3-phosphate (GAPDH) was examined by western blotting; GAPDH was used as a loading control, (a and b; right panel) The relative level of COX-2 was quantified by densitometric measurements (ImageJ). The data represent a typical experiment, whereby similar results were obtained from three experiments. *p<0.05 as compared with untreated cells, #p<0.05 as compared with TQ or IS526 alone | |
Also, inhibition of pp38 or pERK with SB and PD restored COX-2 expression in B16F10 cells (Fig. 3b, left panel). These results were confirmed by immunocytochemistry (Fig. 4a).
These results suggest that the inhibition of TQ-induced COX-2 expression by IS526 occurs via p38 kinase and ERK-1/-2 pathways in A375. And change of COX-2 expression in B16F10 were not a mediator MAPKs pathway.
Inhibitory effect of the combination of TQ and IS526 on tyrosinase activity via the MAPK pathway: The effects of TQ and IS526 alone or as a combination of tyrosinase, MITF protein levels are shown in Fig. 4. Several studies have demonstrated that A375 human melanoma cells express very low levels of MITF and tyrosinase38,39.
In comparison with control cells, those treated with TQ or IS526 showed inhibited expression of tyrosinase in A375 and B16F10. The co-treatment of two cell lines with TQ and IS526 resulted in a significant decrease in the expression of tyrosinase and MITF as compared to cells treated with either TQ or IS526, respectively (Fig. 3).
To investigate the effect of TQ and IS526 alone or as a combination of cellular tyrosinase activity in B16F10 cells, performed the tyrosinase activity assay. It was found that co-treatment of B16F10 cells with TQ and IS526 resulted in significant inhibition of tyrosinase activity.
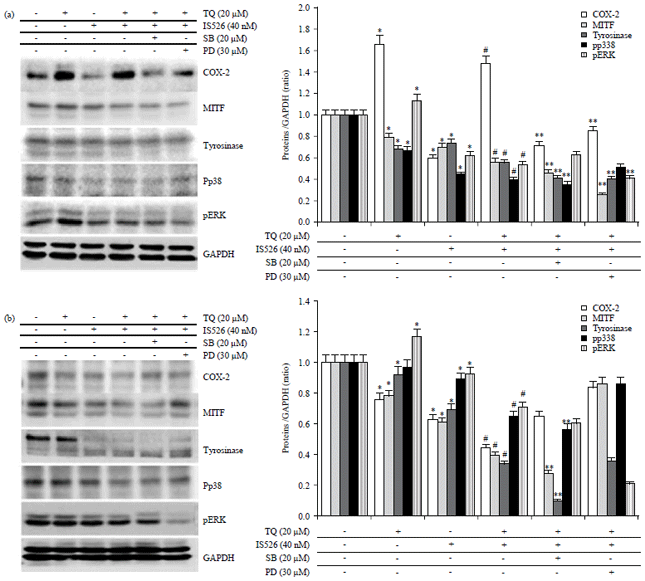 | |
| Fig. 3(a-b): | Effects of PD and SB on co-treatment with TQ and IS526 in COX-2, tyrosinase expression |
A375 cells were untreated or treated with TQ or IS526 for 24 hrs in the absence or presence of SB203580 (SB) or PD98059 (PD), (a and b; left panel) Expression of COX-2,tyrosinase, MITF, phosphorylated extracellular signal-regulated kinase (pERK), pp38 and glyceraldehyde-3-phosphate (GAPDH) was examined by western blotting; GAPDH was used as a loading control, (a and b, right panel) The relative level of proteins was quantified by densitometric measurements (ImageJ). The data represent a typical experiment, whereby similar results were obtained from three experiments, *p<0.05 as compared with untreated cells, #p<0.05 as compared with TQ or IS526 alone, **p<0.05 as compared with the combination of TQ and IS526 | |
These results indicate that TQ in combination with IS526 inhibited the tyrosinase activity and expression of tyrosinase in melanoma cells (Fig. 4b).
The treatment of melanoma cells with the p38 inhibition with SB accelerated the reduction in tyrosinase activity and tyrosinase expression induced by the combination of TQ and IS526. On the contrary, ERK-1/-2 inhibitor, PD, promoted tyrosinase activity and tyrosinase expression in the presence of TQ and IS526 on B16F10 and enhanced the suppressive effect of IS526 or TQ-induced tyrosinase expression in A375 cells (Fig. 3 and 4).
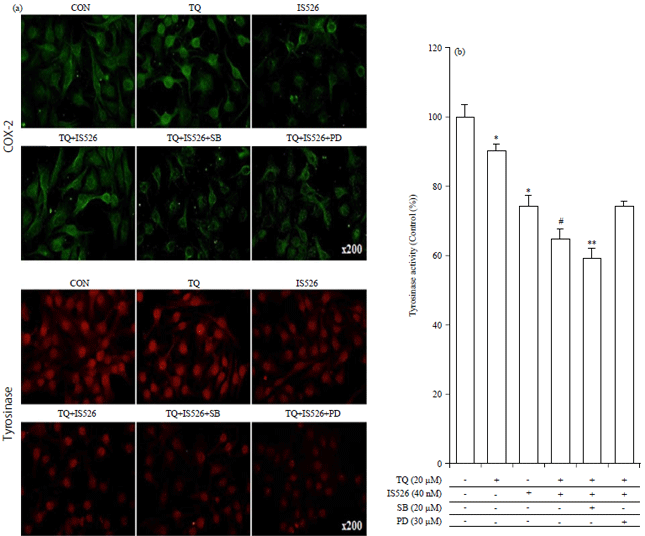 | |
| Fig. 4(a-b): | Effects of PD and SB on TQ and IS526 regulation of COX-2 and tyrosinase activity |
(a) Expression of tyrosinase and COX-2 was examined by Immunofluorescence staining, (b) Cellular tyrosinase activity was measured with an enzyme-linked immunosorbent assay (ELISA) plate reader. The data represent a typical experiment, whereby similar results were obtained from three experiments, *p<0.05 as compared with untreated cells, #p<0.05 as compared with TQ or IS526 alone, **p<0.05 as compared with the combination of TQ and IS526 | |
Taken together, the above results indicate that ERK-1/-2 and p38 kinase pathways are involved in the regulation of tyrosinase activity and expression in melanoma cells.
To examine the role of p38 or ERK in the reduce cell proliferation effect of combined use of the two agents, both cells were treated with SB and PD, that inhibit the p38 or ERK pathway (Fig. 5). Whereby the microscope (Fig. 5a) and MTT assay (Fig. 5b) indicated that SB or PD did not affect cell proliferation in both cells. These results disclosed that the inhibition of cell proliferation in melanoma cells (A375 and B16F10) by combination treatment of TQ and IS526 were not related to the MAPK pathway.
Inhibition of IS526-induced ROS generation by TQ via ERK-1/-2 and p38 pathways in A375 melanoma cells: The effects of TQ and/or IS526 on ROS generation in A375 cells were examined, as ROS is known to cause immediate inflammation in the epidermis and may trigger melanin production15,20,25.
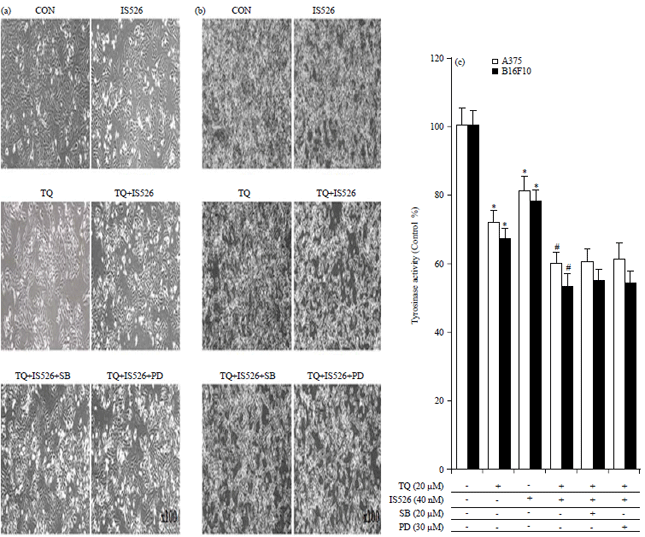 | |
| Fig. 5(a-c): | Effects of PD and SB on TQ and IS526-mediated reduction in cell proliferation |
A375 and B16F10 cells were untreated or treated with TQ or IS526 for 24 hrs in the absence or presence of SB203580 (SB) or PD98059 (PD). (a) A375 cells and (b) B16F10 cells were viewed under a phase-contrast microscope and photographed (100×magnification), (c) Cell proliferation was measured by the MTT assay and the data represent growth inhibition as a percentage of control cells. The data represent a typical experiment, whereby similar results were obtained from three experiments, *p<0.05 as compared with untreated cells, #p<0.05 as compared with TQ or IS526 alone, **p<0.05 as compared with the combination of TQ and IS526 | |
As shown in Fig. 6, TQ reduced the production of ROS, while IS526 increased the intracellular level of ROS. In comparison with IS526 treatment, co-treatment with TQ and IS526 resulted in a significant reduction in the ROS level. The combination of TQ and IS526 with SB or PD resulted in a slight increase in the intracellular ROS level.
These results indicate that TQ reversed the increase in ROS level induced by IS526 via ERK-1/-2 and p38 kinase pathways and the effect was significant as compared with that observed with IS526 treatment. No correlation was observed between ROS and COX-2 level, as these two activities in an opposite manner.
DISCUSSION
Recent studies have shown that the combination of compounds with less toxicity may improve treatment outcomes, as these treatment strategies may enhance the efficiency of the individual agents and prevent drug resistance in human cancer cells.
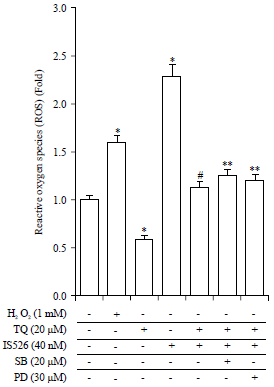 | |
| Fig. 6: | Effects of the combinatorial treatment of TQ and IS526 on ROS production |
A375 cells were exposed to TQ or IS526 for 24 hrs in the absence or presence of SB203580 (SB) or PD98059 (PD), ROS fluorescence was measured using an Flx8000 Bio-Tek fluorometer. Data are presented as the Means ± standard deviation (SD) from three independent experiments performed in triplicates, The data represent a typical experiment, whereby similar results were obtained from three experiments, *p<0.05 as compared with untreated cells, #p<0.05 as compared with TQ or IS526 alone, **p<0.05 as compared with the combination of TQ and IS526 | |
Chemicals from plants and foods may serve as attractive options for such combination therapy, as these are less toxic and more cost-effective as compared to synthetic chemicals37,40,41.
Previous studies showed that TQ and resveratrol combination significant cell inhibition and an increase in caspase-3 indicating cell apoptosis. Also, both drugs raised reactive oxygen species leading to a decrease of glutathione and a minor effect on lipid peroxidation42. The combination of TQ and resveratrol against also able to work synergistically in mice breast cancer36. However, the molecular mechanism underlying TQ and resveratrol combination effect of anti-cancer and anti-tyrosinase activity in melanoma cells has not been elucidated. The research aims to investigate the effect of TQ alone or in combination with IS526 melanoma cells.
The present study demonstrated the effect of TQ in combination with IS526 on A375 and B16F10 cells. In comparison with TQ or IS526 treatment, the combination treatment greatly inhibited the proliferation (Fig. 1), tyrosinase expression and activity (Fig. 3 and 4b) of cells.
The effectiveness of TQ and IS526 as single agents in melanoma prevention and treatment was previously identified. TQ was shown to inhibit cell growth and increase apoptosis in B16F10 and A375 cancer cell lines43,44. TQ induced apoptosis and DNA damage and increased the level of intracellular ROS in B16F10 cells44. Also, TQ exerted an inhibitory effect on the migration of A375 and B16F10 melanoma cells through the inhibition of NLRP3 inflammasome, resulting in a decrease in the proteolytic cleavage of caspase-143. Hatiboglu et al., reported that N. sativa oil and its components (thymoquinone and thymohydroquinone) exert inhibitory effects against mushroom tyrosinase and cellular tyrosinase activities45. Furthermore, N. sativa oil inhibited the expression of tyrosinase protein in B16F10 cells. Several studies have shown that TQ exerts anti-inflammatory effects against some diseases, including bronchial asthma24-26,46. Previous results showed that TQ effectively modulated COX-2 expression and prostaglandin E2 production via PI3K/protein kinase B (Akt) and p38 kinase pathways in the human breast cancer cell line MDA-MB-23124. Moreover, TQ-induced ROS generation regulated apoptosis and inflammation by modulating PI3K/Akt and p38kinase pathways and caused dedifferentiation in rabbit articular chondrocytes through ERK-1/-2 pathway25,47.
Iksan 526 is a resveratrol-enriched transgenic rice DJ526 generated by transferring the resveratrol biosynthesis gene, stilbene synthase. Chung et al. and Jeon et al. reported that the resveratrol-enriched rice accumulated 1.4-1.9 μg g–1 of resveratrol in its grain28,29 and the levels of resveratrol were higher in Iksan526 callus than in resveratrol-enriched rice48. Furthermore, resveratrol-enriched rice extract has been reported to decrease the expression of tyrosinase in B16 melanoma cells30 and control UVB-induced skin aging through the downregulation of oxidative stress-mediated aging, inflammation-induced skin aging and apoptosis-mediated skin aging and wrinkle formation33.
Similar effects on cell proliferation and tyrosinase activity were confirmed and expression in melanoma cells. Also, TQ and IS526 decreased COX-2 expression in B16F10 cells. However, the previous result showed a different pattern as compared with previously reported data on the COX-2 expression and ROS production in A375 cells. TQ increased COX-2 expression and decreased ROS generation, while IS526 induced ROS production and inhibited COX-2 expression (Fig. 3 and 6). These results suggest that TQ- and IS526-regulation of COX-2 expression was independent of ROS production. Furthermore, both drugs had no synergistic effect on COX-2 expression and ROS generation. However, IS526 inhibited TQ-induced COX-2 expression (Fig. 3) and TQ suppressed IS526-induced ROS generation in A375 cells (Fig. 6).
Mitogen-activated protein kinase pathways may function as tumor suppressors and act as the more common pro-oncogenic signals. These pathways control fundamental cellular processes such as growth, proliferation, differentiation, migration, stress responses, immune defense and apoptosis. There are three well-known MAPK pathways in mammalian cells, namely, JNK, ERK-1/-2 and p38 kinasepathway49,50. Numerous studies have demonstrated that the activation of the MAPK pathway results in the expression of COX-2 and that ROS production may trigger signaling pathways of MAPKs49-53.
In the present study, TQ induced the activation of pERK and inhibition of pp38, while IS526 decreased the pp38 level: The combination treatment changes the pp38 level and decreased pERK level as compared with TQ or IS526 treatment (Fig. 3). Treatment for p38 inhibition resulted in the suppression of COX-2 and tyrosinase expression in A375 and B16F10 cells but increased ROS production as compared to the effects observed after TQ or IS526 treatment in A375 cells (Fig. 3 and 6). Also, pERK inhibition decreased COX-2 level in A375 cells but restored tyrosinase activity and expression in B16F10 and ROS production in A375 cells (Fig. 3 and 4). ERK-1/-2 and p38 kinase had no effects on cell proliferation in both cell lines (Fig. 5).
CONCLUSION
In conclusion, results from the present study demonstrated that IS526 inhibits TQ-induced COX-2 production and TQ reduces IS526-induced ROS production through ERK-1/-2 and p38 kinase pathways. ERK-1/-2 and p38 kinase pathways regulate the inhibitory effect of the combination of TQ and IS526 on tyrosinase activity and expression. The combination of TQ and IS526 may synergistically inhibit the growth of melanoma cells.
The limitations of the study include the use of one mouse cell line and one human cell line for experimentation. Therefore, the present study deserves further investigations to be considered as a possible therapeutic option for melanoma, including in vivo animal models.
SIGNIFICANCE STATEMENT
This study discovered the combination of TQ and IS526 inhibits the growth of melanoma cells and regulates COX-2 and ROS. This effective plant-based natural combination is widely available and relatively uncostly. These results suggested that use the combination natural product maybe help in the prevention and treatment of melanoma.
ACKNOWLEDGMENTS
This work was supported by a grant from the Korea Foundation for The advancement of Science and Creativity (KOFAC) funded by the Korea Government (SBJ000025470).
This work was supported by grants from the National Research Foundation of Korea (NRF) funded by the Korean government (MSIP) (No. 2015R1C1A2A01055015, 2017R1D1A3B03033401 and 2020R1I1A3069699) and the Center for Women In Science, Engineering and Technology (WISET) Grant funded by the Ministry of Science and ICT (MSIT) under the Program for Returners into R and D.
This work was carried out with the support of the “Cooperative Research Program for Agriculture Science and Technology Development (Project No. PJ01118805)” Rural Development Administration and the Kongju National University, Republic of Korea.
REFERENCES
- Gata, V.A., C.I. Lisencu, C.I. Vlad, D. Piciu, A. Irimie and P. Achimas-Cadariu, 2017. Tumor infiltrating lymphocytes as a prognostic factor in malignant melanoma. Review of the literature. J. Buon., 22: 592-598.
Direct Link - Renzi, M., J. Schimmel, A. Decker and N. Lawrence, 2005. Management of skin cancer in the elderly. Dermatologic Clinics, 37: 279-286.
CrossRefDirect Link - Liang, Z., C.S. Yang, F. Gu and L.S. Zhang, 2017. A conditionally replicating adenovirus expressing IL-24 acts synergistically with temozolomide to enhance apoptosis in melanoma cells in vitro. Oncol. Lett., 13: 4185-4189.
CrossRefDirect Link - Riker, A.I., N. Zea and T. Trinh, 2010. The epidemiology, prevention, and detection of melanoma. Ochsner J., 10: 56-65.
Direct Link - Roh, M.R., P. Eliades, S. Gupta, J.M. Grant-Kels and H. Tsao, 2017. Cutaneous melanoma in women. Int. J. Women's Dermatol., 3: S11-S15.
CrossRefDirect Link - Narayanan, D.L., R.N. Saladi and J.L. Fox, 2010. Review: Ultraviolet radiation and skin cancer. Int. J. Dermatol., 49: 978-986.
CrossRefDirect Link - Apalla, Z., A. Lallas, E. Sotiriou, E. Lazaridou and D. Ioannides, 2017. Epidemiological trends in skin cancer. Dermatol. Pract. Concept, 7: 1-6.
Direct Link - Berwick, M., E. Erdei and J. Hay, 2009. Melanoma epidemiology and public health. Dermatologic Clinics, 27: 205-214.
CrossRefDirect Link - D'Mello, S.A.N., G.J. Finlay, B.C. Baguley and M.E. Askarian-Amiri, 2016. Signaling pathways in melanogenesis. Int. J. Mol. Sci., Vol. 17.
CrossRefDirect Link - Alam, B., V.K. Bajpai, J. Lee, P. Zhao and J.H. Byeon et al., 2017. Inhibition of melanogenesis by jineol from Scolopendra subspinipes mutilans via MAP-Kinase mediated MITF downregulation and the proteasomal degradation of tyrosinase. Sci. Rep.
CrossRefDirect Link - Niu, C. and H.A. Aisa, 2017. Upregulation of melanogenesis and tyrosinase activity: Potential agents for vitiligo. Molecules, Vol. 22.
CrossRefDirect Link - Zhou, P., J. Qin, Y. Li, G. Li and Y. Wang et al., 2017. Combination therapy of PKCζ and COX-2 inhibitors synergistically suppress melanoma metastasis. J. Exp. Clin. Cancer Res., Vol. 36.
CrossRefDirect Link - Buckman, S., 1998. COX-2 expression is induced by UVB exposure in human skin: implications for the development of skin cancer. Carcinogenesis, 19: 723-729.
CrossRefDirect Link - Becker, M.R., M.D. Siegelin, R. Rompel, A.H. Enk and T. Gaiser, 2009. COX-2 expression in malignant melanoma: a novel prognostic marker? Melanoma Res., 19: 8-16.
CrossRefDirect Link - Bickers, D.R. and M. Athar, 2006. Oxidative stress in the pathogenesis of skin disease. J. Invest. Dermatol., 126: 2565-2575.
CrossRefPubMedDirect Link - Rundhaug, J.E., C. Mikulec, A. Pavone and S.M. Fischer, 2007. A role for cyclooxygenase-2 in ultraviolet light-induced skin carcinogenesis. Mol. Carcinog., 46: 692-698.
CrossRefDirect Link - Liou, G.Y. and P. Storz, 2010. Reactive oxygen species in cancer. Free Radical Res., 44: 479-496.
CrossRefDirect Link - Yilmazer, A., 2017. Cancer cell lines involving cancer stem cell populations respond to oxidative stress. Biotechnol. Rep., 17: 24-30.
CrossRefDirect Link - de Jager, T.L., A.E. Cockrell and S.S. du Plessis, 2017. Ultraviolet Light Induced Generation of Reactive Oxygen Species. In: Ultraviolet Light in Human Health, Diseases and Environment, Ahmad, S.I. (Ed.), Springer, Cham, Switzerland, ISBN: 978-3-319-56016-8, pp: 15-23.
CrossRefDirect Link - Rutkowski, R., S.A. Pancewicz, K. Rutkowski and J. Rutkowska, 2007. [Reactive oxygen and nitrogen species in inflammatory process]. Pol. Merkur Lekarski, 23: 131-136.
Direct Link - Son, Y., Y.K. Cheong, N.H. Kim, H.T. Chung, D.G. Kang and H.O. Pae, 2011. Mitogen-activated protein kinases and reactive oxygen species: how can ROS activate MAPK pathways? J. Signal Trans., Vol. 2011.
CrossRef - Xia, Q., Q. Hu, H. Wang, H. Yang and F. Gao et al., 2015. Induction of COX-2-PGE2 synthesis by activation of the MAPK/ERK pathway contributes to neuronal death triggered by TDP-43-depleted microglia. Cell Death Dis., Vol. 6.
CrossRefDirect Link - Yu, S.M. and S.J. Kim, 2012. Thymoquinone (TQ) regulates cyclooxygenase-2 expression and prostaglandin E2 production through PI3kinase (PI3K)/p38 kinase pathway in human breast cancer cell line, MDA-MB-231. Anim. Cells Sys., 16: 274-279.
CrossRefDirect Link - Woo, C.C., A. Hsu, A.P. Kumar, G. Sethi and K.H.B. Tan, 2013. Thymoquinone inhibits tumor growth and induces apoptosis in a breast cancer xenograft mouse model: the role of p38 MAPK and ROS. PLoS One, Vol. 8.
CrossRefDirect Link - Dur, A., H. Kose, A. Kocyigit, O. Kocaman, M. Ismayilova and F.C. Sonmez, 2016. The anti-inflammatory and antioxidant effects of thymoquinone on ceruleine induced acute pancreatitis in rats. Bratisl Lek Listy, 117: 614-618.
CrossRefDirect Link - Yu, S.M. and S.J. Kim, 2015. The thymoquinone-induced production of reactive oxygen species promotes dedifferentiation through the ERK pathway and inflammation through the p38 and PI3K pathways in rabbit articular chondrocytes. Int. J. Mol. Med., 35: 325-332.
CrossRefDirect Link - Baek, S.H., W.C. Shin, H.S. Ryu, D.W. Lee and E. Moon et al., 2013. Creation of resveratrol-enriched rice for the treatment of metabolic syndrome and related diseases. PLoS ONE, Vol. 8.
CrossRefDirect Link - Chung, H.J., S.P. Sharma, H.J. Kim, S.H. Baek and S.T. Hong, 2016. The resveratrol-enriched rice DJ526 boosts motor coordination and physical strength. Sci. Rep., Vol. 6.
CrossRefDirect Link - Baek, S.H., H.J. Chung, H.K. Lee, R. D'Souza and Y. Jeon et al., 2014. Treatment of obesity with the resveratrol-enriched rice DJ-526. Sci. Rep., Vol. 4.
CrossRefDirect Link - Lee, T.H., J.O. Seo, M.H. Do, E. Ji, S.H. Baek and S.Y. Kim, 2014. Resveratrol-enriched rice down-regulates melanin synthesis in uvb-induced guinea pigs epidermal skin tissue Biomol. Ther., 22: 431-437.
CrossRefDirect Link - Lee, T.H., J.H. Kang, J.O. Seo, S.H. Baek and S.H. Moh et al., 2016. Anti-melanogenic potentials of nanoparticles from calli of resveratrol-enriched rice against UVB-induced hyperpigmentation in guinea pig skin. Biomol. Ther., 24: 85-93.
CrossRefDirect Link - Bishayee, A., 2009. Cancer prevention and treatment with resveratrol: From rodent studies to clinical trials. Cancer Prev. Res., 2: 409-418.
CrossRefPubMedDirect Link - Subedi, L., T.H. Lee, H.M. Wahedi, S.H. Baek and S.Y. Kim, 2017. Resveratrol-enriched rice attenuates UVB-ROS-induced skin aging via downregulation of inflammatory cascades. Oxid. Med. Cell. Longevity, 2017: 1-15.
CrossRefDirect Link - Subedi, L., S.H. Baek and S.Y. Kim, 2018. Genetically engineered resveratrol-enriched rice inhibits neuroinflammation in lipopolysaccharide-activated BV2 microglia via downregulating mitogen-activated protein kinase-nuclear factor kappa B signaling pathway. Oxid. Med. Cell. Longevity.
CrossRefDirect Link - Alobaedi, O.H., W.H. Talib and I.A. Basheti, 2017. Antitumor effect of thymoquinone combined with resveratrol on mice transplanted with breast cancer. Asian Pac. J. Trop. Med., 10: 400-408.
CrossRefDirect Link - Mokhtari, R.B., T.S. Homayouni, N. Baluch, E. Morgatskaya, S. Kumar, B. Das and H. Yeger, 2017. Combination therapy in combating cancer. Oncotarget, 6: 38022-38043.
CrossRefPubMedDirect Link - Vachtenheim, J., H. Novotna and G. Ghanem, 2011. Transcriptional repression of the microphthalmia gene in melanoma cells correlates with the unresponsiveness of target genes to ectopic microphthalmia-associated transcription factor. J. Invest. Dermatol., 117: 1505-1511.
CrossRefDirect Link - Housman, G., S. Byler, S. Heerboth, K. Lapinska, M. Longacre and N. Snyder and S. Sarkar, 2014. Drug resistance in cancer: an overview. Cancers, 6: 1769-1792.
CrossRefDirect Link - Vanneman, M. and G. Dranoff, 2012. Combining immunotherapy and targeted therapies in cancer treatment. Nat. Rev. Cancer, 12: 237-251.
CrossRefDirect Link - Ismail, N., Y. Abdel–Mottaleb, A.A.E. Ahmed and N.N. El-Maraghy, 2018. Novel combination of thymoquinone and resveratrol enhances anticancer effect on hepatocellular carcinoma cell line. Future J. Pharm. Sci., 4: 41-46.
CrossRefDirect Link - Ahmad, I., K.M. Muneer, I.A. Tamimi, M.E. Chang, M.O. Ata and N. Yusuf, 2013. Thymoquinone suppresses metastasis of melanoma cells by inhibition of NLRP3 inflammasome. Toxicol. Appl. Pharmacol., 270: 70-76.
CrossRefDirect Link - Hatiboglu, M.A., A. Kocyigit, E.M. Guler, K. Akdur, A. Nalli, E. Karatas and S. Tuzgen, 2018. Thymoquinone induces apoptosis in B16-F10 melanoma cell through inhibition of p-STAT3 and inhibits tumor growth in a murine intracerebral melanoma model. World Neurosurg., 114: e182-e190.
CrossRefDirect Link - Lee, S.Y., S.M. Lee, W.B. Heo, J.G. Kim and Y.H. Kim, 2011. Effect of Nigella sativa oil on melanogenesis. J. Soc. Cosmet. Scientists Korea, 37: 319-326.
CrossRefDirect Link - Khader, M. and P.M. Eckl, 2014. Thymoquinone: An emerging natural drug with a wide range of medical applications. Iran. J. Basic Med. Sci., 17: 950-957.
Direct Link - Yu, S.M. and S.J. Kim, 2013. Thymoquinone-induced reactive oxygen species causes apoptosis of chondrocytes via PI3K/Akt and p38kinase pathway. Exp. Biol. Med., 238: 811-820.
CrossRefDirect Link - Ku, K.L., P.S. Chang, Y.C. Cheng and C.Y. Lien, 2014. Production of stilbenoids from the callus of Arachis hypogaea: a novel source of the anticancer compound piceatannol. J. Agric. Food Chem., 53: 3877-3881.
CrossRefDirect Link - Subedi, L., T.H. Lee, H.M. Wahedi, S.H. Baek and S.Y. Kim, 2017. Resveratrol-enriched rice attenuates UVB-ROS-induced skin aging via downregulation of inflammatory cascades. Oxid. Med. Cell. Longevity, 2017: 1-15.
CrossRefDirect Link - Burotto, M., V.L. Chiou, J.M. Lee and E.C. Kohn, 2014. The MAPK pathway across different malignancies: A new perspective. Cancer, 120: 3446-3456.
CrossRefDirect Link - Dhillon, A.S., S. Hagan, O. Rath and W. Kolch, 2007. MAP kinase signalling pathways in cancer. Oncogene, 26: 3279-3290.
CrossRefPubMedDirect Link - Eo, S.H., H.S. Cho and S.J. Kim, 2014. Resveratrol regulates type II collagen and COX-2 expression via the ERK, p38 and Akt signaling pathways in rabbit articular chondrocytes. Exp. Therapeut. Med., 7: 640-648.
CrossRefDirect Link - Eo, S.H., D.W. Kim, S.Y. Choi, H.A. Kim and S.J. Kim, 2015. PEP-1-SIRT2 causes dedifferentiation and COX-2 expression via the MAPK pathways in rabbit articular chondrocytes. Exp. Cell Res., 339: 351-359.
CrossRefDirect Link