
Talha Jawaid
Department of Pharmacology, College of Medicine, Al-Imam Mohammad Ibn Saud Islamic University, Othman Ibn Affan Street, 13317 Riyadh, Kingdom of Saudi Arabia
LiveDNA: 91.24416
Mehnaz Kamal
Department of Pharmaceutical Chemistry, College of Pharmacy, Prince Sattam bin Abdulaziz University, 11942 Al-Kharj, Kingdom of Saudi Arabia
LiveDNA: 91.11064
Lubna Azmi
Department of Chemistry, University of Lucknow, Lucknow, India
LiveDNA: 91.10358
Osama A. Alkhamees
Department of Pharmacology, College of Medicine, Al-Imam Mohammad Ibn Saud Islamic University, Othman Ibn Affan Street, 13317 Riyadh, Kingdom of Saudi Arabia
Saud M. Alsanad
Department of Pharmacology, College of Medicine, Al-Imam Mohammad Ibn Saud Islamic University, Othman Ibn Affan Street, 13317 Riyadh, Kingdom of Saudi Arabia
International Journal of Pharmacology
Year: 2020 | Volume: 16 | Issue: 3 | Page No.: 244-256







ABSTRACT
Background and Objective: Bambusa arundinaceae leaves are widely used for treating numerous diseases in Indian traditional medicine. The current study explored the neuroprotective effect of Bambusa arundinaceae leaves ethanolic extract (EEBA) on memory impairment induced by streptozotocin (STZ) in mice. Materials and Methods: The neuroprotective effect of EEBA (100 and 200 mg kg–1 b.wt., p.o.) was assessed in the Morris Water Maze (MWM) test, Pole Climbing Test (PCT) and in the Elevated Plus Maze (EPM) test in comparison with standard piracetam (100 mg kg–1 b.wt., i.p.). The activity of acetylcholinesterase (AChE), malondialdehyde (MDA) and reduced glutathione (GSH) were also measured in various mice brain regions. Gene expression was also performed by RT-PCR and western blot test. Results: Treatment with EEBA 200 mg kg–1 showed a significant effect in all behavioral tests that demonstrated neuroprotective activity. EEBA treatment significantly reduced the AChE and MDA levels in mice brain regions, along with a rise in GSH level. RT-PCR results showed Bax and Bak mRNA were down-regulated, while Bcl-2 mRNA and protein were up-regulated in EEBA (200 mg kg–1) group. NR1, NR2B and GAP-43 proteins lead to the reduction of brain cell damage. EEBA 200 mg kg–1 showed a significant effect by shielding against STZ induced brain damage by interacting with these proteins. Conclusion: The effect of EEBA (200 mg kg–1 b.wt.) on behavioral and biochemical parameters was comparable with that observed in piracetam treated rats. These findings indicated that EEBA may exert a neuroprotective effect that may be accredited to inhibiting AChE and regulating the protein expression in the brain as well as its antioxidant mechanisms.
PDF Abstract XML References Citation
Copyright: © 2020. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Talha Jawaid, Mehnaz Kamal, Lubna Azmi, Osama A. Alkhamees and Saud M. Alsanad, 2020. Neuroprotective Effect of Bambusa arundinaceae Leaves Extract on Learning and Memory Impairment in Mice: Impact on NR2B, NR1 and GAP Pathways. International Journal of Pharmacology, 16: 244-256.
DOI: 10.3923/ijp.2020.244.256
URL: https://scialert.net/abstract/?doi=ijp.2020.244.256
DOI: 10.3923/ijp.2020.244.256
URL: https://scialert.net/abstract/?doi=ijp.2020.244.256
INTRODUCTION
One of the most common neurodegenerative disorders is Alzheimer's Disease (AD) which causes the loss of memory. The AD most frequently affects people1,2 over the age of 65. AD's pathophysiology is multifarious and involves excessive accumulation of neurofibrillary tangles and amyloid-β plaques in the brain3. Increasing evidence indicates that the loss of cholinergic neurons of basal forebrain with reduced acetylcholine (ACh) levels is accountable for the main signs of AD4. ACh is considered to be a major learning and memory-related neurotransmitter4,5. Therefore, for the symptomatic treatment of AD, acetylcholinesterase (AchE) inhibitors that decrease ACh degradation were used in clinical use. Nonetheless, restricted effectiveness, peripheral cholinergic side effects, low bioavailability and most significantly, no significant effect are limitations on the performance of currently available AChE inhibitors on disease progression. Thus, there is a serious medical need to look for new treatment strategies to address various cognitive disorders.
Streptozotocin (STZ), which was originally known as an antibiotic, is a glucosamine-nitrosourea derivative. It is toxic to β-cells of the pancreas is normally transported by Glucose Transporter 2 (GLUT2) and is widely used in animals to cause experimental diabetes in animals6. Administration of STZ by routes such as intracerebroventricular or intraperitoneal injection results in decreased comprehension and increased total tau protein, aggregated Aβ fragments and Aβ deposits. Such improvements are followed by a decline in the α/β ratio of Glycogen Synthase Kinase (GSK-3) in the brain. It has been shown that the administration of STZ in the brain of a mouse induces oxidative stress, neuroinflammation and biochemical changes, which is regarded as a reliable experimental model of neurodegenerative disease early pathophysiological changes6.
STZ has been reported to have an impact on animal memory acquisition, consolidation and recall7. Memory loss caused by STZ is linked with a significant rise in levels of AChE and MDA and decreased GSH8.
Bambusa arundinaceae (F. Gramineae) is a popular medicinal plant locally known as bamboo or bans and popularly known for their industrial applications. Bamboo shoots are eaten in various forms because they are low in fats and high in potassium, carbohydrates, dietary fibers, vitamins and active constituents9. A bamboo culm consists of an internode and a node, which is solid and gives the plant structural integrity. One or more buds are present on the node which produces side branches. The burnt root is applied to bleeding gums; ringworms and tender joints10. Seeds are acrid in taste, used as a laxative in strangury and urinary discharges. Stems are used in diabetes11. The bark is used for rashes in the skin. The leaf is an antileprotic, emmenagogue and febrifuge and antitussive10. It also has estrogenic12, analgesic and antipyretic activities13.
The present study was therefore designed to investigate the neuroprotective effect of Bambusa arundinaceae leaves ethanolic extract (EEBA) on memory impairment induced by STZ in mice in the MWM test, PCT and in the EPM test. The activity of AChE and oxidative stress parameters viz., MDA and GSH were also assessed in the cortex, cerebellum and hippocampus of mice brain. Gene expression was also performed by RT-PCR and western blot test.
MATERIALS AND METHODS
The study was carried out at Hygia Institute of Pharmaceutical Education and Research, Lucknow, India from July, 2014-2015.
Collection of plant: Bambusa arundinacea leaves were collected from the local garden in Lucknow, India. They were authenticated by taxonomists from National Botanical Research Institute, Lucknow, India.
Preparation of ethanolic extract of Bambusa arundinaceae leaves (EEBA): Bambusa arundinaceae leaves were dried in shade, crushed and sieved through a 40-mesh sieve. The powdered leaves underwent Soxhlet extraction at 50°C for 6 h with ethanol. After evaporation of the ethanol, the resulting crude extract was successively washed with chloroform, petroleum ether and ethyl acetate, completely dried and kept in a desiccator. The crude extract for its antiamnesic activities has been subjected to phytochemical investigation14 and pharmacological screening.
Chemicals: Streptozotocin, Triton X-100, sodium hydroxide, sodium chloride, acetylthiocholine iodide, Bovine Serum Albumin (BSA), Folin-Ciocalteu’s hydrochloric acid, 5,5-dithiobis (2-nitro-benzoic acid) (DTNB), 2-thiobarbituric acid (TBA) and trichloroacetic acid (TCA) were purchased from Sigma-Aldrich (St. Louis, USA). Trizol, Revert Aid H Minus first strand cDNA synthesis kit and Maxima SYBR green were acquired from Thermo Fischer Scientific (Mumbai, India). Piracetam (Nootropil®) was purchased from UCB India Pvt. Ltd. (Mumbai, India).
Animals: The experiments were conducted in Swiss albino mice (20-25 g). Each group consisted of six mice (n = 6). Animals were purchased from Central Drug Research Institute's Laboratory Animal Services Division, Lucknow, India. They were kept in polyacrylic cages (22.5×37.5 cm) and kept under standard environmental conditions: Room Temperature (RT), 24-27°C and humidity, 60-65% with 12 h light: 12 dark cycles. The water and food were available ad libitum. The study was approved by the Institutional Animal Ethics Committee (IAEC) in accordance with the Committee for the Purpose of Control and Supervision of Experiments on Animals (1088/07/CPCSEA) regulation and ethical standards were strictly followed during all procedures.
Experimental design: Intracerebral (i.c.) STZ induced memory impairment in mice: STZ administered centrally in subdiabetogenic dose produces memory impairment in rodents8. In freshly prepared artificial CSF (aCSF), STZ was dissolved and administered gradually by i.c. route (0.5 mg kg–1, 10 μL). The mice were anesthetized with chloral hydrate (300 mg kg–1, i.p.) and the scalp had a midline sagittal incision. A 27-gauge hypodermic needle attached to a 100 μL Hamilton syringe was injected (2.5 mm depth) into the brain vertically through the skull. The injection site was on a line traced through the anterior base of the ears 2 mm from either side of the midline15.
Grouping of the mice: STZ induced memory impairment model was validated in mice using piracetam, a nootropic agent. STZ was given to animals for 1-3 days through i.c. route. EEBA (100 and 200 mg kg–1, i.p.) was given for 14 days after the 1st dose of STZ in mice. EEBA was evenly suspended in 1% carboxymethyl cellulose (CMC) dissolved in water and given perorally (p.o.). The group administered with CSF (i.c.) on days 1 and 3 and CMC (10 mL kg–1, p.o.) for 14 days served as a vehicle group. The behavioral testing was initiated from the 14th day after 1st dose of STZ. The standard drug was also administered for 14 days and behavioral tests were performed from 14th day onwards (Fig. 1). Animals were distributed into six groups each containing 6 animals (Table 1). STZ (0.5 mg kg–1, i.c.) was administered 5 min before the day 1 trial (session 1) was started to induce memory impairment16,17.
Acute toxicity study: The acute toxicity tests were carried out in compliance with the Organization for Economic Cooperation and Development (OECD) guidelines 425 for the study of chemical substances for acute oral toxicity18. Mice (n = 6) were treated with varying EEBA doses through the peroral route (50, 250, 500, 1000 and 2000 mg kg–1), though the control group provided saline (10 mL kg–1, i.p.). For any gross effect, all the groups were examined until 6 h and the mortality rate after 24 h of treatment was observed.
Evaluation of memory in mice
Morris Water Maze (MWM) test: The MWM is comprised of a circular pool (45 cm diameter, 26 cm height), filled with water to a depth of 20 cm at 26±1°C and made dense white color. The pool was composed of four hypothetical quadrants.
| Table 1: | Grouping of the mice |
 | |
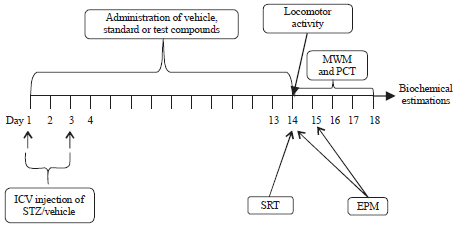 | |
| Fig. 1: | Administration of vehicle, standard and test drugs |
EPM: Elevated plus maze, ICV: Intracerebroventricular, MWM: Morris water maze, PCT: Pole climbing test, SRT: Social recognition test, STZ: Streptozotocin | |
An escape platform was positioned 1 cm under the surface of the water. Four different starting points were set around the pool perimeter for the mice. In a pseudorandom sequence, all four starting points were used once on each of the five training days. The water maze was placed in a large room with a variety of visual signs of the extra maze. The trail started by putting the animal in the water facing the pool wall at one of the starting points. If the animals failed to escape within 120 sec on the platform, the researcher gently placed them there and allowed them to stay for 30 sec. On five consecutive days, each animal has been exposed to a daily session of four trials. Escape Latency Time (ELT) was noted as an index of learning to locate the concealed platform in water maze19,20.
Pole Climbing Test (PCT): The effect of EEBA on STZ induced memory impairment was assessed in Swiss albino mice. The mice were then divided into six groups of six mice in each group and the behavior study in the pole climbing apparatus was carried out. The passive avoidance test is a broadly accepted simple and fast way of testing memory. A "step-through" apparatus consisting of an illuminated and dark compartment (each 20.3×15.9×21.3 cm) adjoining each other was used to determine the passive avoidance response. Floors were made of stainless steel rods of 3.175 mm, set apart by 8 mm. The test was conducted at the same time daily for 2 consecutive days. Every animal was put in the illuminated compartment facing away from the dark compartment on the first day (learning trial). Once the animal enters into the dark compartment absolutely, it experiences an electric shock (1 mA, 3 sec) through the grid floor made up of stainless steel. The time taken for the animal to reach the dark compartment was automatically recorded and defined as step-through latency. The same test procedure was followed on the 2nd day (testing trial). The test was terminated when the animal did not enter the dark compartment within 30 sec and the latency of 30 sec was reported by Joshi and Parle17.
Elevated Plus Maze (EPM) test: The elevated plus maze acted as the paradigm of exteroceptive behavior (where the sensation occurred outside of the body) for evaluating mice’s learning and memory. Two covered arms (16×5×12 cm) and two open arms (16×5 cm) formed the apparatus. The arms extended from a central platform (5×5 cm) and the maze was raised from the floor to a height of 25 cm. Each animal was positioned at the end of the open arm on the 1st day, facing off the central platform. Transfer Latency (TL) was taken as the time it took for an animal to shift into one of the covered arms with all its four legs. On the first day, TL was documented. If within 90 sec the animal does not enter one of the covered arms, it was gently pushed into one of the two covered arms and the TL was assigned as 90 sec. The animal was permitted to explore the maze for 10 sec and then returned to its home cage. Memory retention was observed 24 h after the 1st day trial i.e., on the 2nd day21.
Locomotor activity: After an acclimatization time of 15 min, animals were independently put in the activity cage for 10 min and their activity was observed. The photocell counts were calculated and locomotor activity increase or decrease was determined.
Biochemical estimation of oxidative stress markers: On the 5th day, after the behavioral studies had been completed, mice were sacrificed with an overdose of diethyl ether anesthesia. The brain was separated from each mouse, placed on an ice-cold plate and then dissected into the hippocampus and cerebral cortex.
Preparation of brain homogenate: Samples of brain tissue (hippocampus and cerebral cortex) were washed and homogenized with ice-cold 0.1 M phosphate buffer of pH 7.4, 10 times and used to calculate AChE, MDA and GSH levels22,23.
Estimation of AChE activity: The brain homogenate (500 μL) was combined with 1% Triton X-100 and centrifuged for 60 min at 10,000×g at 4°C. The supernatant was collected and used for the measurement of AChE. The mixture included 0.4 mL of supernatant, 20 μL of acetylthiocholine iodide, 2.4 mL phosphate buffer (pH 8) and 100 μL of DTNB for doing assay. The absorbance shift was measured with a spectrophotometer for 10 min at an interval of 2 min at 412 nm. AChE activity was expressed as μmol min–1 (mg protein)22,24.
Estimation of MDA level: About 0.5 mL of tissue homogenate, 0.5 mL distilled water and 1.0 mL 10% TCA was added, mixed properly and centrifuged for 10 min at 3,000 g. To 0.2 mL supernatant, 0.1 mL TBA was added. The total solution was placed in a water bath at 80°C for 40 min and then cooled at RT. The clear supernatant absorbance was measured in a spectrophotometer at 532 nm. The amount of MDA has been expressed as nmol mg–1 protein22,24.
| Table 2: | RT-PCR study of various proteins |
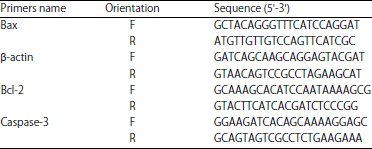 | |
Estimation of GSH level: The amount of GSH was determined by its reaction to DTNB (Ellman’s reagent) to obtain a yellow chromophore whose absorbance was spectrophotometrically determined. The brain homogenate was combined with an identical amount of 10% TCA and centrifuge at 200×g at 4°C for 10 min. The supernatant had been used in estimating GSH. To 0.1 mL of the processed tissue sample, 0.5 mL of DTNB, 2 mL of phosphate buffer (pH 8.4) and 0.4 mL of double-distilled water were added and vigorously shaken on vortex. The absorbance was taken within 15 min, at 412 nm. The amount of GSH has been expressed as nmol mg–1 protein22,24.
RT-PCR (reverse transcription-polymerase chain reaction) examination: Total RNA was extracted from hippocampal tissue using the Trizol reagent, following instructions from the manufacturer. A high-capacity cDNA Reverse Transcription Kit was used. The quantitative RT-PCR was done using the Thermo Scientific Luminaris Color HiGreen qPCR Master Mix. Also, mRNA-specific primers for Bax, Caspase-3, Bcl-2 and β-actin as a housekeeping gene were done and according to Livak and Schmittgen25, relative gene expression was determined after data analysis (Table 2).
Western blot analysis: Brain samples were taken from the mice’s hippocampal tissue. Hippocampal tissue was homogenized with a lysis buffer (50 nM L–1 NaCl, 1% Triton X-100, 1 mM L–1 EDTA, 0.5% SDS, 20 mM L–1 Tris HCl and 0.5% sodium deoxycholate; pH 7.5) and centrifuged for 20 min at 15,000×g. Protein samples (50 μg) per lane were performed on a polyacrylamide gel, transferred to a Polyvinylidene Difluoride (PVDF) membrane (Millipore, Billerica) and blocked for 2 h with 5% milk solution (non-fat dry milk in Phosphate Buffered Saline (PBST). The membrane was incubated overnight with the following specific antibodies at 4°C: phospho-Akt (1:1000; Cell Signaling Technology), rabbit polyclonal anti-ChAT (1:1000; Cell Signaling Technology, Boston, MA, United States), Akt (1:1000; Cell Signaling Technology), phosphoCREB (1:1000 dilution), CREB (1:1000 dilution), ERK1/2 (1:1000 dilution), phospho-ERK1/2 (Thr202/Thr204) (1:1000, dilution), BDNF (1:1000, dilution), NR1 (1:1000, dilution), NR2B (1:1000, dilution), GAP-43 (1:1000, dilution) and mouse monoclonal anti-β-actin (1:10000; dilution). Upon five times washing with Tris Buffered Saline with Tween (TBST), the membranes were then incubated for 1 h at RT with the respective conjugated anti-rabbit IgG (1:10000; dilution). Immunoreactive proteins were quantified using enhanced chemiluminescence (ECL) kit and the relative density of the protein bands was scanned using a LAS 4000 Fujifilm imaging system and analyzed using Quantity-One software through densitometric evaluation.
Estimation of protein: Protein for AchE has been estimated in all brain regions as described earlier by Wang and Smith26 and protein for MDA and GSH by Lowry et al.27 method. Standard BSA (1 mg mL–1) was used.
Statistical analysis: Results were expressed as mean±SEM. The statistical significance of difference among the different groups was determined by one-way ANOVA followed by Bonferroni’s post hoc test using GraphPad Prism 5 software (GraphPad Inc. (California, USA) for MWM test, EPM test, PCT, AChE, MDA and GSH estimation. The significance level for Bonferroni’s multiple comparison test was set to 0.05 for 3 or more groups and p<0.05 was considered statistically significant. Statistical analysis was done by one-way ANOVA followed by Dunnett's multiple comparison assay for RT-PCR analysis and Western blot analysis.
RESULTS
Phytochemical screening: The phytochemical screening results showed the presence of flavonoids, phytosterol, alkaloids, phenolic compounds, reducing sugar, glycosides, proteins and amino acids.
Assessment of acute toxicity: The EEBA showed a wide safety margin as no gross behavioral changes were observed and death was recorded up to 2000 mg kg–1 b.wt., p.o. up to 48 h.
Effect of EEBA on STZ induced memory impairment in the MWM test: As illustrated in Fig. 2, control and aCSF groups showed a significant reduction (p<0.05) in latency time from 3rd session onward compare to session 1. Though, no significant decrease in latency time (p>0.05) was found during all the 5 sessions in STZ treated animals.
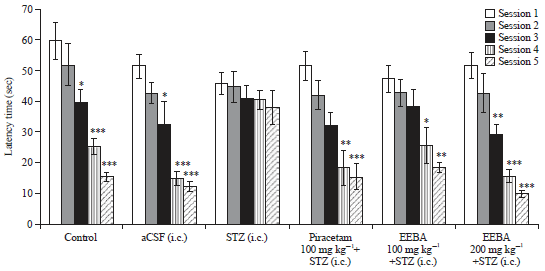 | |
| Fig. 2: | Effect of EEBA on STZ induced memory impairment in MWM test |
Data are expressed as mean escape latency time (s)±SEM (n = 6), *Significant difference (*p<0.05, **p<0.01 and ***p<0.001) in compare to the session 1 | |
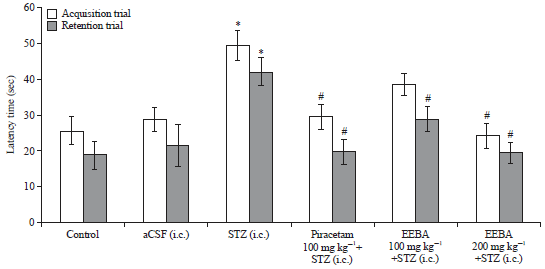 | |
| Fig. 3: | Effect of EEBA on STZ induced memory impairment in the EPM test |
Data are expressed as mean latency time (s)±SEM (n = 6), *Significant difference (*p<0.05) compare to the respective trial of control and aCSF groups and #Significant difference (#p<0.05) in comparison to the STZ group | |
EEBA (100 and 200 mg kg–1) showed dose-dependent amelioration of STZ induced memory impairment as revealed by significant reduction (p<0.05) in latency time in the MWM test. Preventive treatment with piracetam in STZ injected mice showed a significant decrease in latency to reach the hidden platform in the MWM test. No significant change was found among the latency time of session 1 (p>0.05) and 2 (p>0.05) of all the groups.
Effect of EEBA on STZ induced memory impairment in the EPM test: EEBA (100 and 200 mg kg–1) effect was studied on the 14th and 15th day after administration of the first dose of STZ in mice on STZ induced memory impairment in the EPM test. As illustrated in Fig. 3, i.c. injection of STZ caused a significant increase (p<0.05) in latency time during acquisition and retention trials relative to control and aCSF groups suggesting memory impairment.
The standard piracetam avoided STZ induced memory impairment as displayed by a significant reduction (p<0.05) in latency time during acquisition as well as retention trials as compared to the STZ group. EEBA (200 mg kg–1) administration caused significant (p<0.01) reduction in acquisition and retention latencies in comparison to the STZ group. Though, EEBA (100 mg kg–1) reduced latency time significantly (p<0.05) in the retention trial only.
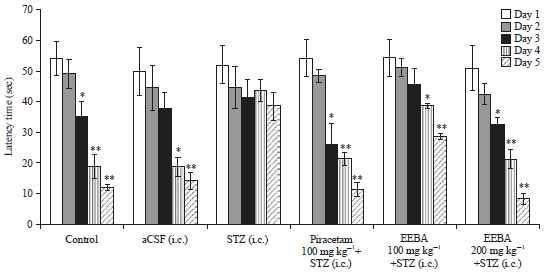 | |
| Fig. 4: | Effect of EEBA on STZ induced memory impairment in PCT |
Data are expressed as mean latency time (s)±SEM (n = 6), *Significant difference (*p<0.05) compare to day 1 of the respective group | |
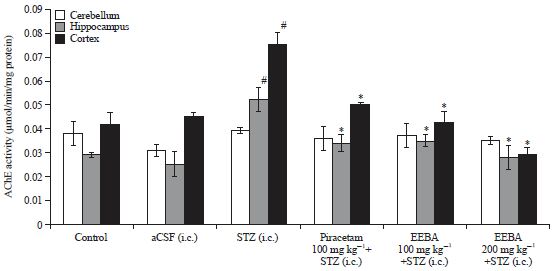 | |
| Fig. 5: | Effect of EEBA on AChE activity in STZ induced memory impaired mice brain |
Data are expressed as mean AChE activity μmol/min/mg protein) ±SEM (n = 6), #Significant increase (#p<0.05) compare to the control group and *Significant decrease (*p<0.05) compare to the STZ group | |
Effect of EEBA on STZ induced memory impairment in PCT: As illustrated in Fig. 4, the control group reported a significant reduction (p<0.05) in latency time from day 3 onwards when compared to day 1. i.c. administration of aCSF had no significant effect on PCT as shown by a significant decrease (p<0.05) in latency time on 4th and 5th day compared to day 1.
STZ treated animals took more time while the piracetam and EEBA (100 and 200 mg kg–1) treated groups showed significantly (p<0.05) less time to reach the pole in the pole climbing apparatus.
Effect of EEBA on locomotor activity: There was no significant change in the spontaneous locomotor activity between different groups was observed.
Effect of EEBA on AChE activity in STZ induced memory impaired mice brain: The AChE activity (μmol/min/mg protein) was assessed in brain regions of mice after the conduction of behavioral studies. STZ caused significant rise in AChE activity in hippocampus (p<0.05) and cortex (p<0.01) in comparison to control and aCSF groups. There was however no significant difference (p>0.05) in AChE activity of control and aCSF group (Fig. 5).
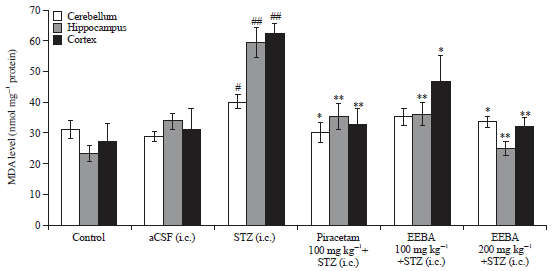 | |
| Fig. 6: | Effect of EEBA on MDA level in STZ induced memory impaired mice brain |
Data are expressed as mean MDA level (nmol mg–1 protein) ±SEM (n = 6), #Significant increase (#p<0.05 and ##p<0.01) compare to the control group and *Significant reduction (*p<0.05 and **p<0.01) compare to the STZ group | |
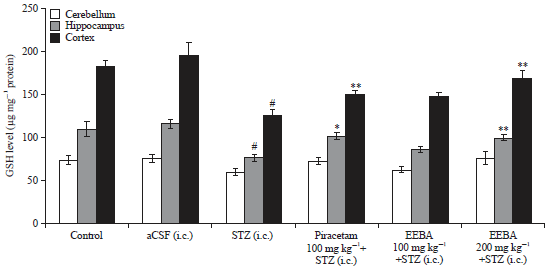 | |
| Fig. 7: | Effect of EEBA on GSH level in STZ induced memory impaired mice brain |
Data are expressed as mean GSH level (μg mg–1 protein) ±SEM (n = 6), #Significant reduction (#p<0.01) compare to the control group and *Significant increase (*p<0.05 and **p<0.01) compare to the STZ group | |
Piracetam significantly (p<0.05) prevented STZ induced elevation in AChE activity in cortex and hippocampus and cerebellum. Treatment with EEBA (100 and 200 mg kg–1) significantly (p<0.05) reduced AChE activity in hippocampus (p<0.05) and cortex (p<0.05) of STZ injected mice.
Effect of EEBA on MDA level in STZ induced memory impaired mice brain: As illustrated in Fig. 6, the MDA level significantly increased in cerebellum (p<0.05), cortex (p<0.01) and hippocampus (p<0.01) of STZ treated mice as compared to control and aCSF groups. Treatment with piracetam significantly reduced MDA level in cerebellum (p<0.05), cortex (p<0.01) and hippocampus (p<0.01) compare to STZ group.
EEBA (100 mg kg–1) treatment decreased STZ induced level of MDA in hippocampus (p<0.05) and cortex (p<0.01) significantly. Further, EEBA (200 mg kg–1) caused significant decrease (p<0.05) in level of MDA in STZ treated mice brain regions.
Effect of EEBA on GSH level in STZ induced memory impaired mice brain: A significant decrease in GSH level (μg mg–1 protein) was found in cortex (p<0.01) and hippocampus (p<0.01) of STZ treated mice in comparison to control and aCSF groups. Piracetam treatment significantly (p<0.05) stopped the STZ induced decrease in the level of GSH in the hippocampus (p<0.05) and cortex (p<0.01) of mice brain (Fig. 7).
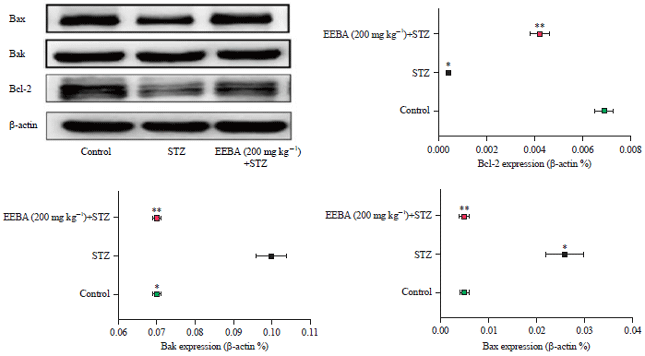 | |
| Fig. 8: | Protein expressions of Bcl-2, Bak and Bax in the hippocampus of the experimental mice in RT-PCR study |
Statistical analysis was done by one-way ANOVA followed by Dunnett's multiple comparison assay, *Denotes significance when compared to control group p<0.05), **Denotes significance when compared to STZ (p<0.05) | |
As illustrated in Fig. 7, further administration of EEBA (200 mg kg–1) significantly increased level of GSH in hippocampus (p<0.05) and cortex (p<0.01) in comparison to STZ group. Though, EEBA (100 mg kg–1) had no significant effect (p>0.05) on GSH level in mice brain regions. No significant change (p>0.05) was observed in the cerebellum following STZ or drug administration.
RT-PCR: Expressions of Bcl-2, Bak and Bax mRNA are demonstrated in Fig. 8 for excluding the variations due amount and nature of RNA, the results recorded were adjusted according to the expression of GAPDH. Hippocampal tissue in the STZ group was indicated by considerably amplified the levels of Bak and Bax and significantly declined the levels of Bcl-2. Their levels were appreciably inverted in EEBA (200 mg kg–1). EEBA 200 group showed the best results which were almost comparable to the piracetam group. In comparison to the piracetam group, EEBA (200 mg kg–1) demonstrated a more significant effect in the up-regulation of Bcl-2 protein and down-regulation of p53 protein. In mitochondrial pathway cell apoptosis chiefly involves the Bcl-2, Bak and Bax (both belong to the Bcl family) control the secretion of proapoptotic factors from mitochondria. In the present study, Bax and Bak mRNA (proapoptotic) were down-regulated, while Bcl-2 mRNA and protein which are antiapoptotic were up-regulated in EEBA (200 mg kg–1) group opposite to STZ group. Also, p53 regulates the Bcl-2 family proteins its expression in the given experiment was appreciably reduced in EEBA (200 mg kg–1) group opposite effect was recorded in the STZ group. These findings indicated that EEBA could exert its memory enhancement by interacting with these proteins.
Western blot analysis: This analysis was carried out for recording the neuroprotective activity of EEBA (200 mg kg–1). NR2B, NR1 and GAP-43 levels in STZ treated mice were significantly lower in the hippocampus than they were in the control group. In comparison, EEBA (200 mg kg–1) group had significantly higher NR2B, NR1 and GAP-43 protein levels in the hippocampus than the STZ group did. As shown in Fig. 9, protein expressions of NR1 and NR2B were augmented by almost two times and five times in the hippocampus of mice of the STZ group as compared to the mice of the control group, whereas their expressions were appreciably reduced in EEBA mice. The NR1 expression was amplified by almost six-folds in STZ treated mice while EEBA treated mice partially prevented this effect. Majorly two pathways are involved in cell apoptosis (death receptor pathway and mitochondrial pathways). Reported findings say that interaction of death receptor and its ligand e.g. , interaction of NR2B/NR1 are important for initiating apoptosis (extrinsic pathway).
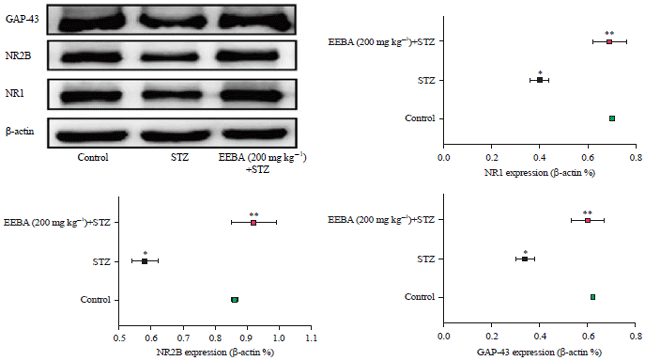 | |
| Fig. 9: | Expressions of NR1, NR2B and GAP-43 in the hippocampus of the experimental mice in the Western blot test |
Statistical analysis was done by one-way ANOVA followed by Dunnett's multiple comparison assay, *Denotes significance in comparison to control group (p<0.05), **Denotes significance in comparison to STZ (p<0.05) | |
Prior studies have shown that suppressing NR1, NR2B and GAP-43 proteins lead to a reduction of brain cell damage. EEBA 200 mg kg–1 showed a significant effect by shielding against STZ induced brain damage.
DISCUSSION
Bamboo is a multi-purpose plant recognized primarily for its industrial use but now known as a possible source of natural antioxidants and bioactive constituents. All parts of the bamboo plant have medicinal applications viz., culm, rhizome, leaves, stems, shoots, seeds and roots. Research has shown that bamboo is a rich source of antioxidants and frequent use of bamboo-based products can reduce the risk of chronic age-related diseases including Alzheimer's disease, cardiovascular diseases, Parkinson's disease, diabetes and cancer28.
The findings of the current study showed that EEBA prevented STZ induced memory impairment in all paradigms of behavioral, MWM test, EPM test and PCT in mice. Besides, the neuroprotective effect of EEBA was related to a significant decrease in AChE and MDA levels in mice brain regions, along with a rise in GSH level.
Intracerebroventricular (i.c.) injection of STZ is a model based on brain insulin resistance that mimics many of human and animal pathophysiological aspects such as memory impairment, changes in the metabolism of glucose, oxidative stress and the phosphorylation of tau protein29.
In the present study, i.c. injection of STZ on 1st and 3rd days caused memory impairment in mice before session 1 of the MWM test, EPM and PCT. There was no significant change in the latency time during all 5 sessions in the streptozotocin treated group. EEBA (100 and 200 mg kg–1) demonstrated antiamnesic activity, as shown in the MWM test by a significant reduction (p<0.05) in latency time. In EPM test, EEBA (200 mg kg–1) produced significant (p<0.01) reduction in acquisition and retention latencies. In PCT, EEBA (100 and 200 mg kg–1) treated groups showed significantly (p<0.05) less time to reach the pole in the pole climbing apparatus.
These findings agree with a large number of previous researches documenting memory impairing the impact of STZ in rodents30. In both the behavioral experiments, the commonly used nootropic agent piracetam ameliorated streptozotocin mediated memory impairment16,17.
Preventive administration of EEBA in mice for 5 days, dose-dependently improved STZ induced memory impairment. EEBA (100 and 200 mg kg–1) demonstrated antiamnesic activity, as shown in the MWM test by a significant reduction (p<0.05) in latency time. In EPM test, EEBA (200 mg kg–1) produced significant (p<0.01) reduction in acquisition and retention latencies. In PCT, EEBA (100 and 200 mg kg–1) treated groups showed significantly (p<0.05) less time to reach the pole in the pole climbing apparatus. These findings proposed the neuroprotective effect of EEBA in STZ induced memory impairment via a reduction in oxidative stress and phosphorylation of tau protein.
In any group, there was no significant change in spontaneous locomotor activity indicating that the behavioral effects are not correlated with any change in mice’s locomotor activity.
The AChE activity and oxidative stress parameters were examined in mice brain regions to understand the potential mechanism of action of the EEBA. STZ in this study caused a significant rise in AChE activity in the cerebellum, cortex and hippocampus of mice. EEBA administration has shown a dose-dependent inhibition of STZ mediated increased activity of AChE. AChE inhibition by EEBA may lead to an increased level of ACh in the brain that may be responsible for its neuroprotective effect. Standard piracetam also decreased the activity of AChE in brain regions of mice.
Several clinical and preclinical researches have confirmed that the pathogenesis of Alzheimer’s disease includes oxidative stress2. The current study measured GSH and MDA in brain regions of mice that acted as markers of lipid peroxidation and endogenous antioxidant, respectively. A rise in the MDA level shows the degeneration of the neurons. GSH is a natural antioxidant present in the body that plays an essential role in maintaining the intracellular redox state. In the STZ model, the levels of GSH and MDA were assessed once behavioral studies were completed. In contrast, to control values, STZ treated mice showed a significant decrease in GSH and a rise in MDA levels in brain regions, suggesting high oxidative stress. Standard piracetam significantly increased the level of GSH and reduced the MDA level in the brain of mice suggesting its antioxidant action, as earlier reported by Alikatte et al.16. The EEB administration reduced oxidative stress caused by STZ as presented by significantly raised GSH and decreased level of MDA in mice brain regions.
RT-PCR results displayed that after EEBA (200 mg kg–1) administration Bax, Bcl-2 and Bak mRNA expression levels were significantly effective than that in control and STZ group, Bax and Bak have a pro-apoptotic function in cells31, while Bcl-2 has the opposite function to Bax, in other words, the anti-apoptotic function32. The rate of apoptosis is inversely proportional to the degree of expression of Bcl-233. CNS pathological mechanism is very complex. In the present research, the apoptosis-related factors expressions were studied, including Bax, Bak and Bcl-2 in the hippocampus, while the precise mechanism yet to be explored.
NR2B and NR1 expression were studied using a western blot test. The findings of this research displayed that the administration of EEBA (200 mg kg–1) significantly increased both NR2B and NR1 expression, indicating that NMDA receptor signaling could be involved in the neuroprotective effect of EEBA on cognitive dysfunction. The NMDA receptor especially plays an essential role in synaptic plasticity, that has been involved in learning and memory34. For a better understanding of EEBA (200 mg kg–1) effect on cognitive dysfunctions in STZ treated group, the impact on synaptic plasticity was examined. GAP-43 is an intracellular growth protein that plays a crucial role in synaptic plasticity, regulating growth cone guidance and outgrowth of neurites35. The findings showed a significant reduction in levels of GAP-43 in hippocampal tissue following treatment with STZ. Nonetheless, treatment with EEBA (200 mg mg–1) improved the expression of GAP-43 considerably. Therefore, EEBA (200 mg kg–1) impact on cognitive functional recovery can be due to EEBA induced synaptic plasticity and neurogenesis in the hippocampus.
In present study, the EEBA extract significantly increased the GSH level in the brain and decreased the AchE enzyme and MDA levels. STZ causes memory impairment by decreasing GSH and increasing the AchE enzyme and MDA level in various mice brain regions. Bax and Bak mRNA (proapoptotic) were down-regulated, while Bcl-2 mRNA and protein (antiapoptotic) were up-regulated in EEBA (200 mg kg–1) group opposite to STZ group. Thus, the result obtained from the current study specify that EEBA extract showed dose-dependent activity and it is effective to treat dementia.
CONCLUSION
Oxidative stress, neuroinflammation and biochemical changes in the various brain regions induced by STZ can produce memory impairment linked with a significant rise in levels of AChE and MDA and decreased GSH. EEBA oral treatment produced a significant reduction in latency time in the MWM test, reduced acquisition and retention latencies in the EPM test and decreased the time to reach the pole in PCT. EEBA treatment significantly reduced the AChE and MDA levels in mice brain regions, along with a rise in GSH level.
Bax and Bak mRNA (proapoptotic) were down-regulated, while Bcl-2 mRNA and β-actin which are antiapoptotic were up-regulated in the EEBA group. EEBA showed a significant neuroprotective effect by suppressing NR1, NR2B and GAP-43 protein expression
SIGNIFICANCE STATEMENT
This work exposed the neuroprotective effect of EEBA on streptozotocin induced learning and memory impairment in mice. It showed a significant effect by decreasing the activity of AchE enzyme in various brain regions, increased acetylcholine level that helped to increase learning and memory. This study will help the researchers to uncover the importance of alternative plant-based remedy based on EEBA in the treatment of memory impairment that many researchers were not able to explore. Thus a novel approach that EEBA is effective and safe to treat a patient suffers from Alzheimer’s disease may be arrived at.
ACKNOWLEDGMENTS
The authors are thankful to Hygia Institute of Pharmaceutical Education and Research, Lucknow, India for providing the necessary facilities to carry out this research. Authors would also like to thank National Botanical Research Institute, Lucknow, India for plant authentication and Central Drug Research Institute, Lucknow, India for providing the animals.
REFERENCES
- Jewart, R.D., J. Green, C.J. Lu, J. Cellar and L.E. Tune, 2005. Cognitive, behavioral and physiological changes in Alzheimer disease patients as a function of incontinence medications. Am. J. Geriatr. Psychiatry, 13: 324-328.
CrossRefDirect Link - Kamat, P.K., 2015. Streptozotocin induced Alzheimer's disease like changes and the underlying neural degeneration and regeneration mechanism. Neural Regen. Res., 10: 1050-1052.
CrossRefPubMedDirect Link - Mullane, K. and M. Williams, 2013. Alzheimer's therapeutics: Continued clinical failures question the validity of the amyloid hypothesis—but what lies beyond? Biochem. Pharmacol., 85: 289-305.
CrossRefDirect Link - Wang, R., H. Yan and X.C. Tang, 2006. Progress in studies of huperzine A, a natural cholinesterase inhibitor from Chinese herbal medicine. Acta Pharmacol. Sin., 27: 1-26.
CrossRefDirect Link - Tota, S., K. Hanif, P.K. Kamat, A.K. Najmi and C. Nath, 2012. Role of central angiotensin receptors in scopolamine-induced impairment in memory, cerebral blood flow and cholinergic function. Psychopharmacology, 222: 185-202.
CrossRefPubMedDirect Link - Gao, C., Y. Liu, Y. Jiang, J. Ding and L. Li, 2014. Geniposide ameliorates learning memory deficits, reduces Tau phosphorylation and decreases apoptosis via GSK 3β pathway in streptozotocin-induced Alzheimer rat model. Brain Pathol., 24: 261-269.
CrossRefDirect Link - Agrawal, R., E. Tyagi, G. Saxena and C. Nath, 2009. Cholinergic influence on memory stages: A study on scopolamine amnesic mice. Indian J. Pharmacol., 41: 192-196.
CrossRefPubMedDirect Link - Saxena, G., S.P. Singh, R. Agrawal and C. Nath, 2008. Effect of donepezil and tacrine on oxidative stress in intracerebral streptozotocin-induced model of dementia in mice. Eur. J. Pharmacol., 581: 283-289.
CrossRefDirect Link - Choudhury, D., J.K. Sahu and G.D. Sharma, 2012. Value addition to bamboo shoots: A review. J. Food Sci. Technol., 49: 407-414.
CrossRefDirect Link - Rathod, J.D., N.L. Pathak, R.G. Patel, N.P. Jivani and N.M. Bhatt, 2011. Phytopharmacological properties of Bambusa arundinacea as a potential medicinal tree: An overview. J. Applied Pharmaceut. Sci., 1: 27-31.
Direct Link - Macharla, S.P., V. Goli, D. Santhosha and N.A. Ravinder, 2012. Antdiabetic activity of Bambusa arundinaceae stem extracts on alloxan induced diabetic rats. J. Chem. Biol. Phys. Sci. Sect. B: Biol. Sci., 2: 832-835.
Direct Link - Jawaid, T., A. Awasthi and M. Kamal, 2015. Estrogenic activity of a hydro-alcoholic extract of Bambusa arundinaceae leaves on female Wistar rats. J. Adv. Pharmaceut. Technol. Res., 6: 19-24.
CrossRefPubMedDirect Link - Jawaid, T., M. Kamal and J. Singh, 2017. Analgesic and antipyretic activities of ethanolic extract of Bambusa arundinacea leaves. Int. J. Pharmaceut. Sci. Res., 8: 2867-2872.
CrossRefDirect Link - Awasthi, H., S. Tota, K. Hanif, C. Nath and R. Shukla, 2010. Protective effect of curcumin against intracerebral streptozotocin induced impairment in memory and cerebral blood flow. Life Sci., 16: 87-94.
CrossRefPubMedDirect Link - Alikatte, K.L., B.R. Akondi, V.G. Yerragunta, P.R. Veerareddy and S. Palle, 2012. Antiamnesic activity of Syzygium cumini against scopolamine induced spatial memory impairments in rats. Brain Dev., 34: 844-851.
CrossRefPubMedDirect Link - Joshi, H. and M. Parle, 2006. Antiamnesic effects of Desmodium gangeticum in mice. Yakugaku Zasshi, 126: 795-804.
CrossRefPubMedDirect Link - Tota, S., P.K. Kamat, H. Awasthi, N. Singh, R. Raghubir, C. Nath and K. Hanif, 2009. Candesartan improves memory decline in mice: Involvement of AT1 receptors in memory deficit induced by intracerebral streptozotocin. Behav. Brain Res., 199: 235-240.
CrossRefPubMedDirect Link - Jawaid, T., A.K. Shakya, H.H. Siddiqui and M. Kamal, 2014. Evaluation of cucurbita maxima extract against scopolamine-induced amnesia in rats: Implication of tumour necrosis factor alpha. Z. Naturforsch., C: J. Biosci., 69: 407-417.
CrossRefPubMedDirect Link - Kulkarni, S.K., S.B. Kasture and S.A. Mengi, 2010. Efficacy study of Prunus amygdalus (almond) nuts in scopolamine-induced amnesia in rats. Indian J. Pharmacol., 42: 168-173.
CrossRefDirect Link - Tota, S., C. Nath, A.K. Najmi, R. Shukla and K. Hanif, 2012. Inhibition of central angiotensin converting enzyme ameliorates scopolamine induced memory impairment in mice: Role of cholinergic neurotransmission, cerebral blood flow and brain energy metabolism. Behav. Brain Res., 232: 66-76.
CrossRefDirect Link - Awasthi, H., D. Kaushal and H.H. Siddiqui, 2012. Chronic inhibition of central angiotensin-converting enzyme ameliorates colchicine-induced memory impairment in mice. Sci. Pharm., 80: 647-662.
CrossRefPubMedDirect Link - Livak, K.J. and T.D. Schmittgen, 2001. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method. Methods, 25: 402-408.
CrossRefDirect Link - Wang, C.S. and R.L. Smith, 1975. Lowry determination of protein in the presence of Triton X-100. Anal. Biochem., 63: 414-417.
CrossRefDirect Link - Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - Nirmala, C., M.S. Bisht, H.K. Bajwa and O. Santosh, 2018. Bamboo: A rich source of natural antioxidants and its applications in the food and pharmaceutical industry. Trends Food Sci. Technol., 77: 91-99.
CrossRefDirect Link - De Oliveira Santos, T., C.H.Y. Mazucanti, G.F. Xavier and A. da Silva Torrao, 2012. Early and late neurodegeneration and memory disruption after intracerebroventricular streptozotocin. Physiol. Behav., 107: 401-413.
CrossRefDirect Link - Zhang, Y., R. Ding, S. Wang, Z. Ren and L. Xu et al., 2018. Effect of intraperitoneal or intracerebroventricular injection of streptozotocin on learning and memory in mice. Exp. Therapeut. Med., 16: 2375-2380.
CrossRefPubMedDirect Link - Ersahin, M., D. Ozsavci, A. Sener, O.B. Ozakpinar and H.Z. Toklu et al., 2013. Obestatin alleviates subarachnoid haemorrhage-induced oxidative injury in rats via its anti-apoptotic and antioxidant effects. Brain Injury, 27: 1181-1189.
CrossRefDirect Link - Tso, M.K., E. Lass, J. Ai and R.L. MacDonald, 2015. Valproic acid Treatment after Experimental Subarachnoid Hemorrhage. In: Neurovascular Events after Subarachnoid Hemorrhage: Towards Experimental and Clinical Standardisation, Fandino, J., S. Marbacher, A.R. Fathi, C. Muroi and E. Keller (Eds.). Springer International Publishing, Switzerland, ISBN: 978-3-319-04981-6, pp: 81-85.
- Topkoru, B.C., O. Altay, K. Duris, P.R. Krafft, J. Yan and J.H. Zhang, 2013. Nasal administration of recombinant osteopontin attenuates early brain injury after subarachnoid hemorrhage. Stroke, 44: 3189-3194.
CrossRefDirect Link - Nakanishi, S., 1992. Molecular diversity of glutamate receptors and implications for brain function. Science, 258: 597-603.
CrossRefDirect Link - Aigner, L., S. Arber, J.P. Kapfhammer, T. Laux and C. Schneider et al., 1995. Overexpression of the neural growth-associated protein GAP-43 induces nerve sprouting in the adult nervous system of transgenic mice. Cell, 83: 269-278.
CrossRefDirect Link