
Fanping Wang
School of Laboratory Medicine, Xinxiang Medical University, 453003 Xinxiang, China
Lulu Chen
School of Laboratory Medicine, Xinxiang Medical University, 453003 Xinxiang, China
Shanshan Zhu
School of Laboratory Medicine, Xinxiang Medical University, 453003 Xinxiang, China
Shanshan Wang
Third Affiliated Hospital of Xinxiang Medical University, Xinxiang, 453003 Henan, China
Chen Chen
School of Laboratory Medicine, Xinxiang Medical University, 453003 Xinxiang, China
Weibin Zhang
School of Laboratory Medicine, Xinxiang Medical University, 453003 Xinxiang, China
Xia Wang
School of Laboratory Medicine, Xinxiang Medical University, 453003 Xinxiang, China
Jingjing Zhang
School of Laboratory Medicine, Xinxiang Medical University, 453003 Xinxiang, China
Mingyong Wang
School of Laboratory Medicine, Xinxiang Medical University, 453003 Xinxiang, China
LiveDNA: 86.20838
International Journal of Pharmacology
Year: 2018 | Volume: 14 | Issue: 3 | Page No.: 369-376







ABSTRACT
Background and Objective:Sulforaphane (SFN) is a potent chemopreventive agent obtained from cruciferous vegetables. A series of trials have demonstrated that SFN can induce cell apoptosis in a variety of tumor cells. However, the actual mechanism underlying how SFN induces apoptotic effects in tumor cells remains unclear. Current study aimed to investigate the effects of SFN on growth and apoptosis of acute leukemia cells and its molecular cascades involving in preventing tumor growth. Materials and Methods: In this study, the cell counting Kit-8 assay was used to investigate the consequences of SFN treatment on proliferation of KG1a and K562 cells to determine the appropriate drug concentration. Furthermore, the apoptosis rate of KG1a and K562 cells was assessed by FACS. The expression of Bax, Bcl-2 and caspase-3 in both cells which were affected by sulforaphane was analyzed by quantitative reverse transcription-polymerase (PCR) and Western-blotting, respectively. Results: SFN could inhibit the proliferation of KG1a and K562 cells in a dose and time dependent manner. The numbers of apoptotic cells in SFN treated groups were more than those in control group. In addition, SFN could also increase protein and mRNA expressions of Bax and caspase-3 in a dose dependent manner, but decrease Bcl-2 expression as compared to control group. Conclusion: This study indicated that SFN inhibited tumor cell proliferation and promoted apoptosis in KG1a and K562 cells through modulation of Bax, Bcl-2 and caspase-3. Current work also shed light on SFN as a promising therapeutic agent for the treatment of leukemia.
PDF Abstract XML References Citation
Received: July 27, 2017;
Accepted: October 30, 2017;
Published: March 15, 2018
Copyright: © 2018. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Fanping Wang, Lulu Chen, Shanshan Zhu, Shanshan Wang, Chen Chen, Weibin Zhang, Xia Wang, Jingjing Zhang and Mingyong Wang, 2018. Sulforaphane Induces Apoptosis of Acute Human Leukemia Cells Through Modulation of Bax, Bcl-2 and Caspase-3. International Journal of Pharmacology, 14: 369-376.
DOI: 10.3923/ijp.2018.369.376
URL: https://scialert.net/abstract/?doi=ijp.2018.369.376
DOI: 10.3923/ijp.2018.369.376
URL: https://scialert.net/abstract/?doi=ijp.2018.369.376
INTRODUCTION
Acute myeloid leukemia (AML) is a malignant clonal disease of hematopoietic stem cells resulting from genetic alterations that lead to deregulation of proliferation, differentiation and cell death in hematopoietic progenitors1,2. Among all kinds of adult leukemia, the incidence of acute myeloid leukemia was the highest and the remission rate of chemotherapy was low3. Therefore, looking for a more effective drug for the treatment against leukemia is still the goal in clinical settings.
Epidemiological studies have suggested that a frequent intake of vegetables of cruciferous family could reduce the cancer risk and lead to a weaker metastasis of tumors, such as lung cancer, pancreatic cancer, prostate cancer and so on4,5. Sulforaphane (SFN) is an isothiocyanate (ITC) obtained from cruciferous vegetables and is particularly high in broccoli, brussels sprouts and cabbages6 and has been shown to inhibit the malignant growth in cancer cells of various origins with little or no toxicity toward normal cells7-9. SFN is a potent anticancer agent, however, the mechanism and molecular target of SFN remain unclear.
Apoptosis is a type of programmed cell death that is ideal for cancer treatment10,11. Plenty of chemotherapeutic drugs induce apoptosis of cancer cells. It has been shown that SFN has the pro-apoptotic effects on many human cancers such as colon cancer, liver cancer and cervical cancer and so on12-14. Shang et al.15 reported that SFN induced apoptosis of human leukemia HL-60 cells. The execution of apoptosis depends on the balance between pro-apoptotic protein Bax and anti-apoptotic Bcl-2 family members16. Caspases are also integral parts of the apoptotic pathway. Among them, caspase-3 is believed to be one of the most common members involving in the execution of apoptosis in various cell lineages17. These proteins control the mitochondrial outer membrane permeabilization (MOMP) and the release of mitochondrial inter-membrane proteins such as cytochrome c18.
The purpose of study is based on the hypothesis that SFN induces apoptosis of the leukemia cells through the modulation of Bax, Bcl-2 and caspase-3. In the present study, the potential molecular mechanisms of leukemia cell’s apoptosis by SFN were examined in vitro. The results confirmed SFN as a potent drug preventing tumor cell growth through activating BAX and caspase-3 apoptotic pathway.
MATERIALS AND METHODS
This study was carried out in 2014-2016. in the labs of School of Laboratory Medicine, Xinxiang Medical University, Xinxiang, China.
Reagents: R-Sulforaphane (SFN) was purchased from LKT Laboratories, Inc. (st. Paul, MN). The Cell Counting Kit-8 (CCK-8) was obtained from Dojindo Molecular Technologies Inc. (Kumamoto, Japan). The FITC Annexin V Apoptosis Detection Kit was provided by BD Pharmingen (San Diego, USA). RNA extraction kit, reverse transcription PCR reagent kit and SYBR Premix Ex Taq kit were purchased from Takara Company (Dalian, China) and from Shanghai Tiangen Company. Antibodies against Bax, Bcl-2, caspase-3 and GAPDH were obtained from Abcam Company (Cambridge, UK) and horseradish peroxidase (HRP)-conjugated secondary antibodies (Goat Anti-Rabbit IgG) were purchased from CW Biotech Company (Beijing, China).
Cells: The human acute myeloid leukemia KG1a and K562 cells were provided from the Cancer Institute of Southern Medical University (Guangzhou, China), maintained in RPMI-1640 medium, supplemented with 10% fetal bovine serum, 100 U mL–1 penicillin and 100 μg mL–1 streptomycin in a humidified atmosphere of 5% CO2 at 37°C.
Cell proliferation assay: Cell viability was determined with a cell counting Kit-8 (CCK-8). KG1a and K562 cells (3000/well) were seeded into 96-well plates and cultured in the medium with different concentrations of SFN (0, 2, 4, 6, 8, 10, 12 μmol L–1) for different times (24, 48 and 72 h) and subsequently10 μL CCK-8 reagent was added to each well for additional 4 h. Then, the absorbance of every well was measured at 450 nm wavelength with a microplate reader and the cell survival ratio was expressed as a percentage vs. control. Three triplicates were experimented to determine each data point.
Apoptosis assay: Cell apoptosis were determined with an Annexin V-FITC/PI Apoptosis Detection Kit. KG1a cells and K562 cells (4×105/well) were plated into 6-well plates. After incubation with SFN (0, 4, 8, 12 μmol L–1) for 48 h, the cells were washed with PBS twice and stained with an Annexin V-FITC/PI Apoptosis Detection Kit according to the manufacturer’s instructions. The apoptotic rate of KG1a and K562 cells were measured by FACS. Finally, the apoptotic rate was analyzed by the Cell Quest Analysis software.
Reverse transcription and qPCR analysis: Following treatments of KG1a and K562 cells with 0, 4, 8 and 12 μmol L–1 SFN for 48 h, RNA was isolated from both kinds of cells with TRIzol reagent. cDNA was synthesized with 1 μg of mRNA using a high capacity cDNA reverse transcription kit according to the manufacturer’s instructions. Subsequently, cDNA was amplified by real-time quantitative PCR (qPCR) with a SYBR premix Ex Taq kit (Takara, Dalian, China) using an ABI 7300 Sequence Detection System. The primers used in qPCR were listed as follows:

PCR conditions were as follows: one cycle at 95°C for 3 min, followed by 40 cycles at 95°C for 30s, 55°C for 30s and 72°C for 1 min. The specificity of the PCR products was test by dissociation curves. Each reaction was performed in triplicate. For each individual analysis, one of the samples was designated as the calibrator and given a relative value of 1.0. All quantities were expressed as n-fold relative to the calibrator.
Western blotting: Total proteins were extracted from 2-5×106 cells with an ice-cold SDS protein lysis buffer. Protein concentration was measured with the Micro BCA Protein Assay Reagent kit. Proteins were separated by 10% SDS-PAGE electrophoresis and transferred onto polyvinylidene difluoride (PVDF) membranes. The membranes were blocked with 5% non-fat milk in TBST buffer for 1 h and subsequently incubated with primary antibodies against caspase-3 (1:1,000), Bax (1:1,000), Bcl-2 (1:1,000), or GAPDH (1:10,000) overnight at 4°C. Following washing with TBST 3 times, membranes were incubated with the secondary antibodies conjugated with peroxidase at (1:5,000) for 1 h at room temperature. Afterwards, the membranes were washed with TBST 3 times. The antigen-antibody complexes were visualized using the ECL detection system. At last, protein bands were analyzed with the Image J soft-ware. Each sample was repeated in triplicate.
Statistical analysis: Statistical analysis was performed using statistical product and service solutions (SPSS) software (version 16.0, IBM Inc., New York) data presented were Mean±SD from three different experiments. Statistical significance between different groups was determined using Students’ t-test. A value of p<0.05 was considered statistically significant.
RESULTS
SFN inhibited proliferation in KG1a and K562 cells: The CCK-8 assay showed that SFN could inhibit the growth of leukemia cells, which was indicated by a declined cell viability trend in a dose and time dependent manner following treatments with different concentrations of SFN (Fig. 1a, b). Microscopic observation also showed that a significant decrease in the numbers of A549 and H460 cells following treatments with various concentrations of SFN for 48 h as compared to control group (Fig. 1c).
SFN induced apoptosis in KG1a and K562 cells: To further explore whether SFN could cause apoptosis of acute leukemia cells, KG1a and K562 cells were stained with Annexin V-FITC and PI and the apoptotic rate was analyzed by flow cytometry. The result showed that apoptotic rates of KG1a cells were 0, 3.12, 13.33 and 28.18% following treatments with SFN at 0, 4, 8 and 12 μmol L–1 for 48 h. Consistently, the apoptotic rates of K562 cells were 0, 2.81, 10.63 and 30.27% following treatments with 0, 4, 8 and 12 μmol L–1 SFN for 48 h (Fig. 2). These experiments indicated that the apoptotic rates of KG1a and K562 cells were gradually increased with the increase of SFN concentrations.
SFN regulated the expression of Bcl-2, Bax and caspase-3 in KG1a and K562 cells: The mRNA level of Bax was remarkably increased in KG1a and K562 cells when treated with SFN (Fig. 3a, b), while the mRNA expression of Bcl-2 was slightly inhibited by SFN in K562 cells, but not in KG1a cells (Fig. 3b, e). Additionally, it was also measured caspase-3 expression in KG1a and K562 cells following treatments with SFN. The mRNA expression level of caspase-3 was inhibited by SFN in both cells in a dose-dependent manner (Fig. 3c, f).
To further determine the apoptotic effects of SFN in the leukemia cell. The protein level of Bax, Bcl-2 and caspase-3 in KG1a and K562 cells was examined by Western blotting following treatments with SFN. Interestingly, it was found that SFN could markedly inhibit Bcl-2 protein expression and enhance Bax and caspase-3 protein expressions in a dose dependent manner in both kinds of cells, which did not completely correlate the mRNA expression data (Fig. 4).
Together, these results indicate that SFN can induce leukemia cells apoptosis, which may involve its effects on up-regulating of Bax and caspase-3 level expression and down-regulating Bcl-2 expression level.
DISCUSSION
In the present study, it is the first time to present evidence that SFN could inhibit proliferation of the acute myeloid leukemia KG1a and K562 cells in vitro.
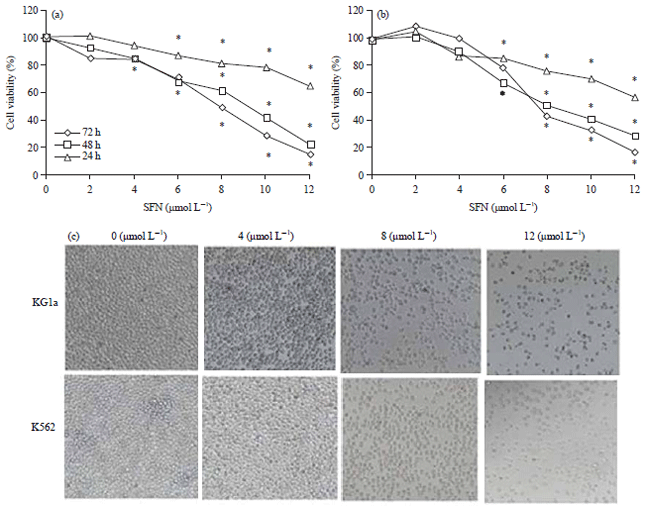 | |
| Fig. 1: | Inhibition effects of SFN (Sulforaphane) on cell viability were detected by CCK-8 assay on (a) KG1a, (B) K562 and (c) Inhibition effects of SFN on cell density were examined by optical microscope |
Data shown are the Means±SD from 3 independent experiments.*p<0.05 compared to the control | |
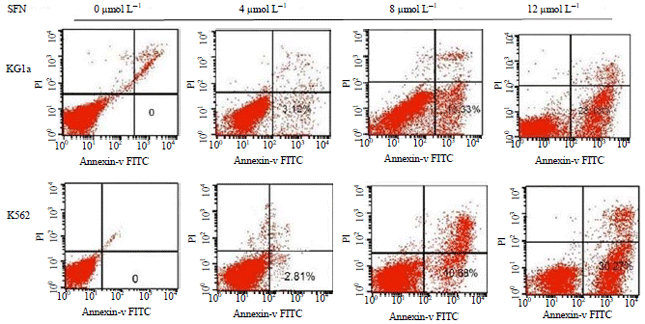 | |
| Fig. 2: | Apoptosis of KG1a and K562 cells treated with SFN for 48 h (by FACS) |
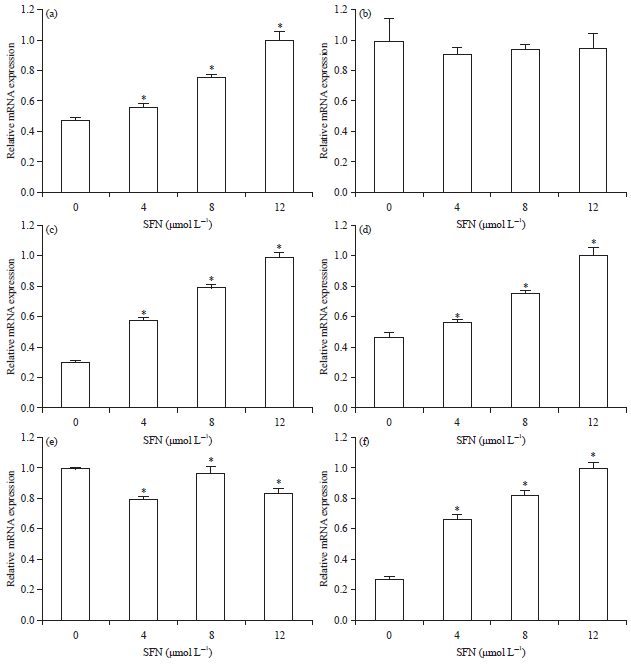 | |
| Fig. 3: | qPCR analysis of Bax, Bcl-2 and Caspase-3 expression in KG1a and K562 cells |
Data shown are the Means±SD from three independent experiments.*p<0.05 compared to the control | |
Most importantly present study showed that SFN regulated apoptosis of leukemia cells in a time and dose dependent manner. Sulforaphane is an isothiocyanate (ITC) obtained from cruciferous vegetables. A series of clinical trials have demonstrated that SFN can induce cell differentiation, lead to cell cycle arrest and promote cell apoptosis in a variety of human cancer types19,20.This study could give guidance for SFN as a new potential agent against leukemia.
Apoptosis is regulated by anti-apoptotic and pro-apoptotic effectors including Bcl-2 family. Bcl-2 and Bax are two representative members of Bcl-2 family. It is known that Bax promotes and Bcl-2 prevents cell apoptosis21,22. The expression of Bax and Bcl-2 in cell remains a homeostasis, which forms an apoptosis regulation system that controls cell proliferation and apoptosis. When Bax is over-expressed, it will form homodimer and induce cell apoptosis. In another way, the Bcl-2 expression increase causes Bax homodimer to dissociate and form heterodimers with Bcl-2, which is more stable than Bax homodimer, inducing suppression on cell apoptosis. Therefore, the ratio of Bax to Bcl-2 determines the occurrence of cell apoptosis, which is also the key factor of the prognosis of malignant tumor23.
In order to examine how SFN induced apoptosis in leukemia cell, the expression of Bax and Bcl-2 in KG1a and K562 cells following treatments with different concentrations of SFN was examined.
 | |
| Fig. 4: | Western blotting analysis of Bax, Bcl-2 and Caspase-3 expression in KG1a and K562 cells |
Western blotting date confirmed that the Bax protein expression was markedly increased and Bcl-2 protein expression was decreased in the SFN treated KG1a and K562 cells, while the expression of Bcl-2 mRNA was slightly inhibited in K562 cells but was not changed in KG1a cells following treatments with SFN. There was a slightly reduced Bcl-2 mRNA level in SFN treated K562 cells, however, there was no change of Bcl-2 transcription level in SFN treated KG1a cell lines. This discrepancy of Bcl-2 protein decrease with its unchanged mRNA levels in these cell lines need to be further studied in terms of protein synthesis and degradation. However, such phenomenon that the protein expression does not always correlate with gene transcription was documented in other studies as well24.
Caspases is a family of protease enzymes playing essential roles in programmed cell death25. In this study attention was focused on caspase-3, one of the most important factors in apoptosis in various cell types in which it cleaves most caspase-related substrates that are induced in the process of apoptosis26-28. Here, it was found that caspase-3 was increased in KG1a and K562 cells at both mRNA and protein levels in a dose-dependent manner following treatments with SFN.
In summary, this study showed that SFN inhibited cell proliferation and revealed that SFN up-regulated Bax and caspase-3 and down-regulated Bcl-2, which led to intrinsic apoptosis in KG1a and K562 human leukemia cells. Such results provided evidence that SFN could be a potent anticancer agent in myeloid leukemia cells. In addition, it was confirmed that SFN could induce apoptosis and inhibit proliferation in KG1a and K562 cells.
CONCLUSION
These results revealed that SFN could cause apoptosis in 2 kinds of leukemia KG1a and K562 cells, which correlated with upregulation of Bax and caspase-3 and down-regulation of Bcl-2. These findings suggested that SFN could be used as a potential anticancer agent against acute myeloid leukemia based on its effective pro-apoptosis activities.
SIGNIFICANCE STATEMENT
This study discovered that SFN a chemo-preventive agent could induce apoptosis of acute human leukemia cells through modulation of Bax, Bcl-2 and caspase-3, which could help uncover the mechanisms of inhibitive role of SFN in proliferation of leukemia cells. This data showed that SFN could be potentially used as a new drug for the treatment of leukemia in clinical setting.
ACKNOWLEDGMENTS
This study was financed by Program for New Century Excellent Talents in University of China (No.NCET-13-0990), Science and Technology of Education Department of Henan Province (No.15A320009 and No.13B320220), Program for National Undergraduate Innovation Training (No.201510472002) and Scientific Research Fund Project of Xinxiang Medical University (No. 2013QN119). The authors would like to thank Mrs Cabatic Maureen, Institute of Psychiatry and Neuroscience, Medical University of Vienna, Vienna, Austria, for her editing of this paper .
REFERENCES
- Pomares, H., C.M. Palmeri, D. Iglesias-Serret, C. Moncunill-Massaguer and J. Saura-Esteller et al., 2016. Targeting prohibitins induces apoptosis in acute myeloid leukemia cells. Oncotarget, 7: 64987-65000.
CrossRefDirect Link - Yang, X.Y., M.Y. Zhang, Q. Zhou, S.Y. Wu and Y. Zhao et al., 2016. High expression of S100A8 gene is associated with drug resistance to etoposide and poor prognosis in acute myeloid leukemia through influencing the apoptosis pathway. OncoTargets Therapy, 9: 4887-4899.
CrossRefDirect Link - Ferrara, F. and C.A. Schiffer, 2013. Acute myeloid leukaemia in adults. Lancet, 381: 484-495.
CrossRefDirect Link - Lam, T.K., L. Gallicchio, K. Lindsley, M. Shiels and E. Hammond et al., 2009. Cruciferous vegetable consumption and lung cancer risk: A systematic review. Cancer Epidemiol. Prevention Biomarkers, 18: 184-195.
CrossRefDirect Link - Srivastava, R.K., S.N. Tang, W. Zhu, D. Meeker and S. Shankar, 2011. Sulforaphane synergizes with quercetin to inhibit self-renewal capacity of pancreatic cancer stem cells. Frontiers Biosci. (Elite Edn.), 3: 515-528.
CrossRefDirect Link - Hahm, E.R. and S.V. Singh, 2010. Sulforaphane inhibits constitutive and interleukin-6-induced activation of signal transducer and activator of transcription 3 in prostate cancer cells. Cancer Prevent. Res., 3: 484-494.
CrossRefDirect Link - Clarke, J.D., R.H. Dashwood and E. Ho, 2008. Multi-targeted prevention of cancer by sulforaphane. Cancer Lett., 269: 291-304.
CrossRefDirect Link - Sangthong, S. and N. Weerapreeyakul, 2016. Simultaneous quantification of sulforaphene and sulforaphane by reverse phase HPLC and their content in Raphanus sativus L. var. caudatus Alef extracts. Food Chem., 201: 139-144.
CrossRefDirect Link - Herr, I. and M.W. Buchler, 2010. Dietary constituents of broccoli and other cruciferous vegetables: Implications for prevention and therapy of cancer. Cancer Treat. Rev., 36: 377-383.
CrossRefDirect Link - Heo, S.K., E.K. Noh, G.D. Gwon, J.Y. Kim and J.C. Jo et al., 2016. Radotinib inhibits acute myeloid leukemia cell proliferation via induction of mitochondrial-dependent apoptosis and CDK inhibitors. Eur. J. Pharmacol., 789: 280-290.
CrossRefDirect Link - Su, Z., Z. Yang, Y. Xu, Y. Chen and Q. Yu, 2015. Apoptosis, autophagy, necroptosis, and cancer metastasis. Mol. Cancer, Vol. 14.
CrossRef - Gamet-Payrastre, L., P. Li, S. Lumeau, G. Cassar, M.A. Dupont et al., 2000. Sulforaphane, a naturally occurring isothiocyanate, induces cell cycle arrest and apoptosis in HT29 human colon cancer cells. Cancer Res., 60: 1426-1433.
Direct Link - Park, S.Y., G.Y. Kim, S.J. Bae, Y.H. Yoo and Y.H. Choi, 2007. Induction of apoptosis by isothiocyanate sulforaphane in human cervical carcinoma HeLa and hepatocarcinoma HepG2 cells through activation of caspase-3. Oncol. Rep., 18: 181-187.
CrossRefDirect Link - Delbridge, A.R.D. and A. Strasser, 2015. The BCL-2 protein family, BH3-mimetics and cancer therapy. Cell Death Differentiation, 22: 1071-1080.
CrossRefDirect Link - Shang, H.S., Y.L. Shih, C.H. Lee, S.C. Hsueh and J.Y. Liu et al., 2017. Sulforaphane-induced apoptosis in human leukemia HL-60 cells through extrinsic and intrinsic signal pathways and altering associated genes expression assayed by cDNA microarray. Environ. Toxicol., 32: 311-328.
CrossRefDirect Link - Czabotar, P.E., G. Lessene, A. Strasser and J.M. Adams, 2014. Control of apoptosis by the Bcl-2 protein family: Implications for physiology and therapy. Nat. Rev. Mol. Cell Biol., 15: 49-63.
CrossRefDirect Link - Yan, F., Q. He, X. Hu, W. Li and K. Wei et al., 2013. Direct regulation of caspase-3 by the transcription factor AP-2α is involved in aspirin-induced apoptosis in MDA-MB-453 breast cancer cells. Mol. Med. Rep., 7: 909-914.
CrossRefDirect Link - Behroozaghdam, M., M. Hashemi, G. Javadi, R. Mahdian and M. Soleimani, 2015. Expression of Bax and Bcl2 genes in MDMA-induced hepatotoxicity on rat liver using quantitative Real-Time PCR method through triggering programmed cell death. Iran. Red Crescent Med. J., Vol. 17.
CrossRef - Lin, L.C., C.T. Yeh, C.C. Kuo, C.M. Lee and G.C. Yen et al., 2012. Sulforaphane potentiates the efficacy of imatinib against chronic leukemia cancer stem cells through enhanced abrogation of Wnt/β-catenin function. J. Agric. Food Chem., 60: 7031-7039.
CrossRefDirect Link - Keum, Y.S., 2011. Regulation of the Keap1/Nrf2 system by chemopreventive sulforaphane: Implications of posttranslational modifications. Ann. N. Y. Acad. Sci., 1229: 184-189.
CrossRefDirect Link - Li, X.M., F. Han, D.J. Liu and Y.X. Shi, 2010. Single-prolonged stress induced mitochondrial-dependent apoptosis in hippocampus in the rat model of post-traumatic stress disorder. J. Chem. Neuroanatomy, 40: 248-255.
CrossRefDirect Link - Zhang, W., H. Feng, Y. Gao, L. Sun and J. Wang et al., 2013. Role of Pigment Epithelium-Derived Factor (PEDF) in arsenic-induced cell apoptosis of liver and brain in a rat model. Biol. Trace Element Res., 151: 269-276.
CrossRefDirect Link - Giotakis, A.I., C.K. Kontos, L.D. Manolopoulos, A. Sismanis, M.M. Konstadoulakis and A. Scorilas, 2016. High Bax/Bcl2 mRNA ratio predicts favorable prognosis in laryngeal squamous cell carcinoma, particularly in patients with negative lymph nodes at the time of diagnosis. Clin. Biochem., 49: 890-896.
CrossRefDirect Link - Cosse, J.P., A. Sermeus, K. Vannuvel, N. Ninane, M. Raes and C. Michiels, 2007. Differential effects of hypoxia on etoposide-induced apoptosis according to the cancer cell lines. Mol. Cancer, Vol. 6.
CrossRef - Esperanza, J.A.A., M.V. Aguirre, G.R. Aispuru, C.N. Lettieri, J.A. Juaristi, M.A. Alvarez and N.C. Brandan, 2008. In vivo 5-flourouracil-induced apoptosis on murine thymocytes: Involvement of FAS, Bax and Caspase3. Cell Biol. Toxicol., 24: 411-422.
CrossRefDirect Link - Park, H.J., Y.J. Kim, K. Leem, S.J. Park, J.C. Seo, H.K. Kim and J.H. Chung, 2005. Coptis japonica root extract induces apoptosis through caspase3 activation in SNU-668 human gastric cancer cells. Phytother. Res., 19: 189-192.
CrossRefDirect Link - Wen, X.R., M. Tang, D.S. Qi, X.J. Huang and H.Z. Liu et al., 2016. Butylphthalide suppresses neuronal cells apoptosis and inhibits JNK-Caspase3 signaling pathway after brain ischemia/reperfusion in rats. Cell. Mol. Neurobiol., 36: 1087-1095.
CrossRefDirect Link - Abdi, A., H. Sadraie, L. Dargahi, L. Khalaj and A. Ahmadiani, 2011. Apoptosis inhibition can be threatening in Aβ-induced neuroinflammation, through promoting cell proliferation. Neurochem. Res., 36: 39-48.
CrossRefDirect Link