
Faisal Abdelrahman Alzahrani
Department of Biological Sciences, Rabigh College of Science and Arts, King Abdulaziz University, Jeddah, Rabigh Branch, Rabigh 21911, Saudi Arabia
Saleh Alkarim
Department of Biology, College of Science, King Abdulaziz University, Saudi Arabia
Islam Mohamed Saadeldin
Department of Animal Production, College of Food and Agricultural Science, King Saud University, 11451, Saudi Arabia
LiveDNA: 20.8408
International Journal of Pharmacology
Year: 2018 | Volume: 14 | Issue: 8 | Page No.: 1128-1135







ABSTRACT
Background and Objective: Regenerative medicine through stem cells holds a great promise and potential therapeutics to regenerate the damaged tissue. Exosomes through their complex cargo of proteins and genetic materials can potentiate treatment of damaged tissues and circumvents some of the concerns and limitations in using viable replicating stem cell. The therapeutic effect of bone marrow mesenchymal stem cells (BM-MSC)-derived exosomes on diethylnitrosamine-induced liver injury was explored in the present study. Materials and Methods: The MSCs were isolated from rats’ bone marrow and characterized by flow cytometry. Exosomes were isolated from MSCs through gradient ultracentrifugation and identified by transmission electron microscopy. Liver was injured with a single intraperitoneal injection of 100 mg kg–1 diethylnitrosamine in 1 mL PBS. BM-MSCs-exosomes were intravenously injected in liver-injured rats. Serum levels of aspartate aminotransferase (AST), alanine aminotransferase (ALT), Alkaline phosphatase (ALP) and albumin were measured. Liver homogenate levels of superoxide dismutase (SOD), catalase (CAT), glutathione peroxidase (GPX) and malondialdehyde (MDA) were measured. Cellular changes have been investigated through histopathological examination and relative quantitative polymerase chain reaction (qPCR) of Bax, Bcl2, Tgfβ1, Tnfa, Cox2 and NFkB transcripts. Data were analyzed by one-way ANOVA followed by Tukey's Honestly Significant Difference test. Significance was considered at p<0.05. Results: MSCs-exosomes significantly reduced serum levels of liver enzymes ALT, AST and ALP, as compared to liver-injured rats. Exosomes also significantly improved the antioxidant enzymatic activities of SOD, CAT, GPx and decreased MDA in liver tissues. In addition to restoration of the deteriorated hepatic histology, exosomes also inhibited expression of the apoptotic gene, Bax and the tissue transforming gene, Tgfβ, whereas they induced the expression of the anti-apoptotic gene, Bcl2 and liver regeneration-related genes, TNFa, NFkB, Cox2. Conclusion: These results suggest ameliorative effect for exosomes on liver injury through induction of liver cells regeneration and inhibition of oxidative stress, apoptosis and epithelial-to mesenchymal transition (EMT) marker, which may be helpful to understand the mechanism of liver diseases and highlight new therapeutic strategies for these diseases.
PDF Abstract XML References Citation
Received: March 28, 2018;
Accepted: August 10, 2018;
Published: October 15, 2018
Copyright: © 2018. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Faisal Abdelrahman Alzahrani, Saleh Alkarim and Islam Mohamed Saadeldin, 2018. Ameliorative Effect of Mesenchymal Stem Cells-derived Exosomes on Diethylnitrosamine-induced Liver Injury in Albino Rats. International Journal of Pharmacology, 14: 1128-1135.
DOI: 10.3923/ijp.2018.1128.1135
URL: https://scialert.net/abstract/?doi=ijp.2018.1128.1135
DOI: 10.3923/ijp.2018.1128.1135
URL: https://scialert.net/abstract/?doi=ijp.2018.1128.1135
INTRODUCTION
Liver injuries are the main cause for liver fibrosis, cirrhosis and finally hepatocellular carcinoma (HCC). Liver transplantation is the final treatment option for patients at late stages of advanced cirrhosis or HCC. However, the lack of donors and high cost of such operation are major setbacks for liver transplantation, with no other solution but to wait1.
Previous studies have elucidated that mesenchymal stem cells (MSCs) have therapeutic capabilities on many diseases through paracrine mechanism2. These paracrine actions are mediated mainly by release of soluble factors and encapsulated factors inside nanovesicles particles called exosomes. The latter transfer mRNA, microRNA, long noncoding RNA and proteins between cells during intercellular communication3,4. Moreover, exosomes are ideal therapeutic agents because their complex cargo of proteins and genetic materials has the potential of treating complex diseases5. In addition, exosome-based therapy circumvents some of the concerns and limitations in using viable replicating cells (including MSCs) without compromising the advantages of using complex therapeutic agents such as cells6.
Recently, exosomes produced by MSCs were found to reduce myocardial ischemia/reperfusion damage and renal injury7. Moreover, the cellular interaction regulated by these exosomes plays pivotal roles in liver homeostasis and pathophysiology8-13. However, little is known regarding the therapeutic effect of exosomes on liver injury disease. One published study showed the ability of liver-derived exosomes to induce hepatocyte proliferation after promotion of liver injury in vitro and in vivo14. However, researchers in this previous study used partial surgical removal of liver lobe to check their hypothesis. This model does not mimic the liver injury in patients and is only suitable to study effect of exosomes on regeneration of damage hepatic tissues. Another study by Li et al.15 have reported that exosomes derived from human umbilical cord MSCs have an ameliorative effect on liver fibrosis induced by CCL4 mainly through inhibition of the epithelial-to mesenchymal transition (EMT) marker, TGFb. Similarly, Tan et al.16 found hepatoprotective effect for human embryonic stem cells-MSCs-derived exosomes on drugs induced liver injuries in mice mainly through activation of proliferating cell nuclear antigen (PCNA), Cyclin D1 and Bcl2. Apart from these four molecules (TGFb, PCNA, Cyclin D1, Bcl2), the underlying molecular signaling pathways regulating exosomes therapeutic potential to liver injuries have not elucidated yet. Moreover controversial results were obtained regarding the effect of MSCs-derived exosomes on the oxidative stress generated by damaged liver tissue. Some researchers have shown no effect for exosomes16 and others found significant elevation of antioxidant enzymes and inhibition of reactive oxygen species (ROS) production17. Taken together, the present study aims to evaluate the therapeutic effect of bone marrow -MSCs-derived exosomes on diethylnitrosamine (DEN)-induced liver injury through investigation of structural, functional and molecular changes.
MATERIALS AND METHODS
Chemicals: Unless otherwise stated, all chemicals and reagents were purchased from Sigma-Aldrich (St. Louis, MO, USA).
Isolation and identification of mesenchymal stem cells (MSCs) and their exosomes: The MSCs were isolated from rats bone marrow according to protocol previously described in detail by Soleimani and Nadri18. The MSCs were characterized by flow cytometry using positive stem cell markers anti-CD90 and negative marker anti-CD34, according to the manufacturer’s instruction (Becton, Dickinson). Exosomes were isolated from MSCs using gradient ultracentrifugation method (Optima L-90K, Beckman Coulter) as previously detailed by Bruno et al.19. The isolated exosomes were identified by transmission electron microscopy (TEM) (JEM2100, Joel Inc.) at 80 KV.
Animals and experimental design: This study was conducted on 27 healthy male albino rats with age of 4 weeks and weight of 130 g). The animals were kept in cages and provided by constant temperature (25-27°C) and light (12 h light/dark cycle) sources with free access to food (standard diet) and water. A period of 2 weeks acclimation was given to rats before the start of the experiment. Herein, it followed the guidelines of the Institutional Animal Care and Use Committee of King Abdulaziz University. Male albino rats with the same ages and weights were randomly divided into three groups (n = 9/group): Control vehicle (PBS)-injected group (Cnt), DEN-induced acute liver injury injected by PBS (PBS) and liver-injured group treated by MSC-derived exosomes (Exo) groups. Liver was injured with a single intraperitoneal injection of 100 mg kg–1 DEN in 1 mL PBS. This low single dose was able to induce pre-carcinogenic acute liver injury without notable carcinogenesis20. At the 8th week of the experiment, 100 μL PBS (PBS group) and 250 μg MSC-derived exosomes (Exo group) were intravenously injected. At the end of the experiment (12th week), the rats were sacrificed, after being anesthetized by ether.
| Table 1: | Primers used for real-time PCR |
 | |
During sacrifice time, blood samples were collected and centrifuged at 3500 rpm for 5 min to get serum.
Biochemical parameters: The serum levels of the liver injury biomarkers [aspartate aminotransferase (AST), alanine aminotransferase (ALT), Alkaline phosphatase (ALP)] and albumin were measured using commercially available kits. Liver tissues were homogenized using cold PBS, followed by centrifugation at 5000 rpm for 15 min at 4°C. The supernatants were used to measure the concentration of the lipid peroxidation biomarker MDA and the activity of antioxidant enzymes SOD, CAT, GPX using commercial kits (Bio-diagnostics Co., Cairo, Egypt).
Histopathological staining: Liver tissue samples fixed in 10% neutral-buffered formalin solution were dehydrated in ethanol, cleared in xylene and impeded in paraffin to form tissue blocks. The latter were sectioned (4-5 μm) and the slides were either stained by hematoxylin and eosin (H and E). To determine the extent of liver injuries, randomly selected section fields were examined from each animal.
Molecular analysis by real-time PCR: The qPCR was used to detect the changes in the relative expression of Bax, Bcl2, Tgfβ1, Tnfa, Cox2 and NFkB genes in the liver tissues of rats following treatment by exosomes or PBS (vehicle). To conduct qPCR, isolated total RNA from liver tissue which was then reverse transcribed to form cDNA synthesis as previously described21. Specific primers (Table 1) were designed by the Primer 3 based on the published rat sequence. SYBR green Master Mix was mixed to cDNA and primers and transferred into 0.1 mL Eppendorf tube then placed in a StepOnePlus real time PCR system (Applied Biosystem, USA). The reaction cycle was 95°C for 10 min, followed by 40 cycles at 95°C for 15 sec and 60°C for 30 sec. All reactions were done in triplicate. Relative quantification was performed following the delta-delta Ct method22. Levels were relatively quantified to control samples with normalization to house-keeping genes β-actin.
Statistical analysis: One-way ANOVA using GraphPad Prism 5 (GraphPad Software, Inc., LaJolla, CA, USA) was used to determine the difference between the groups. Comparison of means was carried out with Tukey's Honestly Significant Difference (Tukey’s HSD) test. Data were presented as Mean±SEM and significance was considered at p<0.05.
RESULTS
The isolated BM-MSCs were characterized by their spindle shape and confirmed by positive expression of CD90 and negative expression of CD34 as detected by flow cytometry (Fig. 1a). Exosomes derived from BM-MSCs were examined by TEM as spherical nanovesicles with diameters ranged from 30-100 nm (Fig. 1b). Further confirmation of successive isolation of exosomes was done by flow cytometric analysis which revealed positive expression of exosomal specific marker, CD63 (Fig. 1a).
Serum levels of ALT, AST and ALP were significantly reduced in animals injected by BM-MSCs-derived exosomes when compared with DEN-induced liver injury animals. However, the level of these enzymes in exosomes-treated rats remained higher than that in control (vehicle treated) animals. On the other hand, administration of these exosomes resulted in a significant elevation of serum albumin as compared not only to liver injured animals but also to control (vehicle treated) animals (Table 2).
Remarkably, the liver of DEN-induced sliver injured rats showed lower activities of the three antioxidant enzymes (SOD, GPX and CAT) and higher levels of the lipid peroxidation marker, MDA, than the vehicle treated rats. Notably, injection of MSCs-exosomes significantly increased the activities of these enzymes and decreased the MDA levels in liver (Fig. 2).
Examination of liver tissues slides stained with H and E in DEN-treated rats showed moderate necro-degenerative changes, whereas minimal necrotic cells were detected in the exosomes-treated rats (Fig. 3).
The above mentioned findings suggested a possible regenerative effect for MSCs-exosomes, to further confirm this effect on a molecular basis, changes in the expression of some genes related to apoptosis (Bax), anti-apoptosis (Bcl2) genes were detected , liver regeneration (Tnfa, Cox2, NfkB) and tissue transformation (Tgfβ1) in the liver tissue using qPCR.
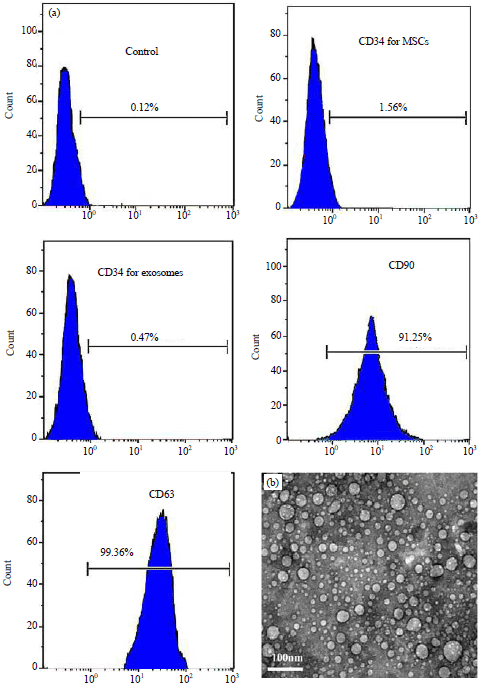 | |
| Fig. 1(a-b): | Identification of cultured MSCs and their exosomes, (a) The isolated MSCs confirmed by flow cytometry which revealed that MSCs were CD90+ and CD34-, while exosomes were CD63+ and also CD34- and (b) Transmission electron microscopic examination shows small nanovesicles (30-100 nm) in the sample isolated from the MSCs culture media by ultracentrifugation |
| Table 2: | Serum levels of cancer markers and liver damage enzymes |
 | |
| Means with different letters within the same row are significantly different at p<0.05. Data was presented as mean±SEM (n = 9). ALT: Alanine aminotransferase, AST: Aspartate aminotransferase, ALP: Alkaline phosphatase | |
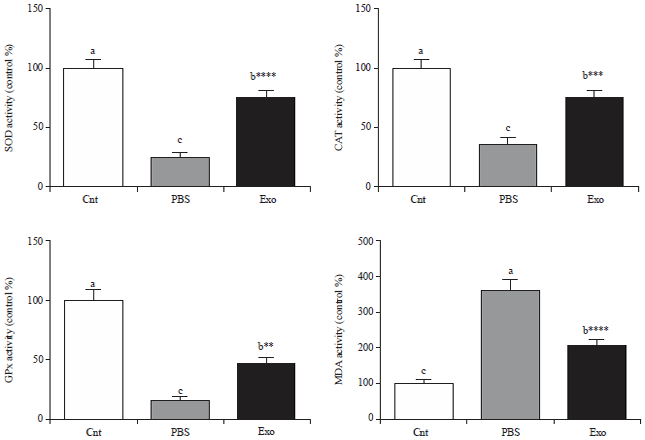 | |
| Fig. 2: | Antioxidant/oxidative stress status of control (vehicle treated) and liver injured rats after treatment by PBS or exosomes derived from MSCs shows levels of SOD, CAT, GPx, and MDA in liver. Values are expressed as Mean±SEM (n = 9). **p<0.01, ***p<0.001, ****p<0.0001 vs PBS group |
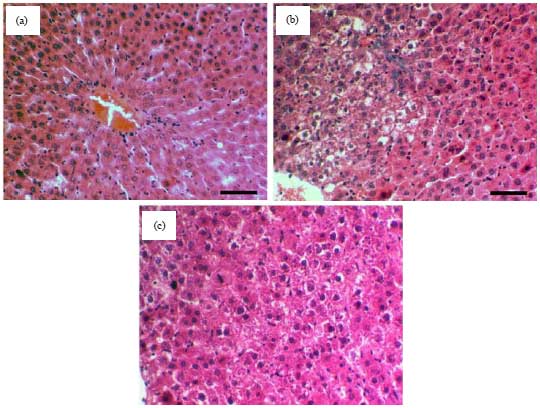 | |
Fig. 3(a-c): | Photomicrographs of H and E stained liver sections of the (a) Control group, (b) PBS group and (c) Exo group |
| Scale bar = 50 µm | |
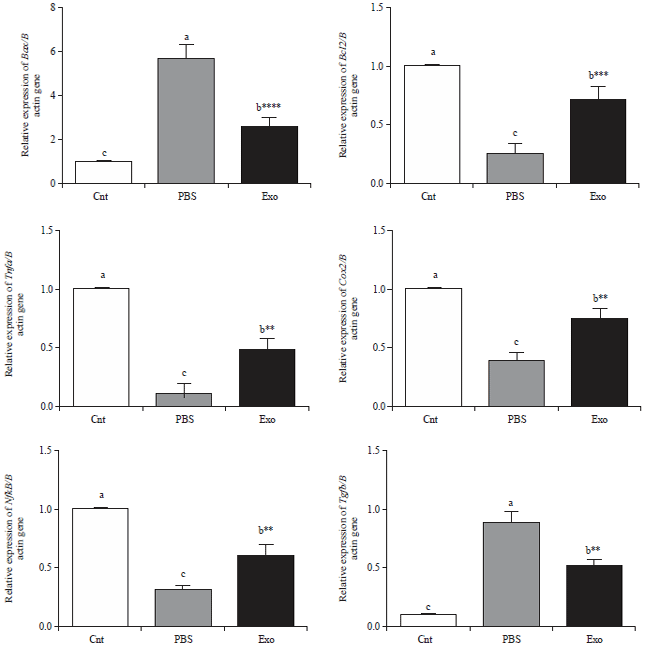 | |
| Fig. 4: | Real time PCR analysis shows changes in relative gene expression of apoptotic gene (Bax), anti- apoptotic gene (Bcl2), liver regenerative genes (Tnfα, Cox2 and NfκB) and EMT gene TGFβ1 in livers of control rats (Cnt) and DEN-induced liver injured rats treated by PBS (PBS) and exosomes derived from MSCs (Exo). Data presented as fold change from the control group. Values are expressed as mean±SEM (n = 9). **p<0.01, ****p<0.0001 vs PBS group |
The inhibition of apoptosis was demonstrated by significant higher Bcl2 and lower Bax mRNA levels in animals treated by MSCs-exosomes as compared to liver injured animals treated by PBS (Fig. 4). Moreover, the MSC-exosomes treated group showed significant increase in mRNA levels of Tnfa, Cox2, NfkB and robust down regulation of Tgfβ1 expression in contrast to the PBS-treated liver injured group (Fig. 4).
DISCUSSION
The first objective for the present study was to evaluate the effect of MSCs-derived exosomes on the rat model of liver injury induced by DEN. To achieve this objective, monitored the functional (liver damage enzymes, albumin and oxidative stress) and structural (histopathology) changes following exosomes injection. MSCs-exosomes induced a significant reduction in serum levels of hepatic damage enzymes (ALT, AST and ALP) in contrast to liver injured rats. In support, similar results were obtained by Li et al.15, who found elevated liver enzymes after treatment of liver fibrosis in mice by exosomes derived from human umbilical cord. Interestingly, serum albumin was significantly higher in the exosomes-treated group than the liver injury group, suggesting a reversal relation between albumin levels and liver injury progression. On the other hand, exosomes originated from MSCs induced a significant increase in serum levels of antioxidant enzymes (SOD, CAT, GPx) and a significant decrease in the level of lipid peroxidation marker MDA in contrast to the liver injured group. Hence, exosomes could mitigate liver injury through regulation of oxidative stress/antioxidant pathway. In contrast to the current results, Tan et al.16 found no effect for exosomes in regeneration of liver damage induced by CCL4 in mice. This contradiction may be attributed to variation in detection method (enzyme activities vs. MnSOD gene expression) and animal model of liver injury (DEN in rats vs. CCL4 in mice). The most important possible cause is detection method as change in gene expression is not always followed by the same change at protein level or enzyme activity. Similar ameliorative effect for exosomes was also observed at histological level in a way that exosomes restored the necro-degenerative changes induced by DEN. In consistent with the current histological findings, Tan et al.16 also reported similar effect for exosomes.
The above biochemical and histological results indicate a possible liver regenerative effect for MSCs-exosomes. Thus, second objective was to explore the molecular mechanism underlying this regenerative effect. To fulfill this goal, changes in the expression of some genes related to liver regeneration (Tnfa, Cox2, NfkB), apoptosis (Bax), anti-apoptosis (Bcl2) and tissue transformation (Tgfβ1) following treatment with exosomes were examined. Current results supported by those reported by 16 demonstrated a significant increase in the expression of all regeneration related genes, Tnfa, Cox2, NfkB, after administration of exosomes. Following liver injury, TNFα, a major initiator for regeneration, activates NFκB and its downstream target COX2, which is a rate limiting step in the synthesis of prostaglandins (PGs) that mediate the regenerative process23.
This TNFα- NFκB- COX2 signaling pathway activated by exosomes may trigger the injured cells arrested in G0/G1 phase to reenter S phase16. Therefore, it is likely that exosomes relieve liver injury, at least in part, via induction of liver cells regeneration. Taken together the anti-oxidative stress effect of exosomes and oxidative stress ability to induce mitochondrial damage and subsequently apoptosis, exosomes may exert its ameliorative effect on liver injury through inhibition of apoptosis. Indeed, the present results demonstrated inhibition of the apoptotic gene, Bax and induction of the ant-apoptotic gene, Bcl2, after exosomes treatment. For its significant role in EMT, TGFb1 is now an important target for EMT inhibitors which used in liver injuries treatment. Consequently, the molecular investigation to involve this gene was examined. Expectedly, exosomes treatment led to significant down regulation to Tgfb1 gene expression in the liver. Other studies also showed inhibition for TGFb1 not only at mRNA level but also at protein level in both liver and serum of rodent models of liver injuries15. As TGFb1 induces liver EMT, mainly via modulation of the Smad2, exosomes may be used, alone or in combination with other drugs, as an EMT inhibitor for treatment of liver injury disease. The findings obtained from molecular investigation collectively suggest that exosomes mainly affirm anti-oxidative stress, anti-apoptotic, anti-EMT and regenerative responses to liver injury. Recent reports showed the ameliorative effect of MSCs-derived exosomes on liver injury induced by CCL4 and concanavalin A, in rats and mice, respectively24,25. Additionally, MSC-derived exosomes showed inhibitory effects on hepatocellular carcinoma growth, progression and metastasis that were induced by DEN toxicity in rats26.
CONCLUSION
Exosomes mediate induction of liver cells regeneration as well as inhibition of oxidative stress, apoptosis and EMT. Exosome as non-cell-based therapy for liver injury circumvents some of the concerns and limitations in using viable replicating stem cells without compromising the advantages of using these cells.
Further investigations are needed to elucidate exact mechanism of action to develop a robust therapeutic method for liver injury.
SIGNIFICANCE STATEMENT
The current results provide a paradigm for studying the paracrine interaction between mesenchymal stem cells and injured liver. This study would offer an alternative therapeutic tool for treating liver injury through the exosomes to overcome the limitations of using the whole mesenchymal stem cells. Further investigations are required to explore the molecular cargoes carried by exosomes which affect the hepatocellular activity either through the ready-made proteins, antioxidants and micro RNA (miRNA) or by translation of transferred mRNAs.
ACKNOWLEDGMENT
This project was funded by the Deanship of Scientific Research (DSR) at King Abdulaziz University, Jeddah, under grant No. G-579-662-38. The authors, therefore, acknowledge with thanks DSR for technical and financial support.
REFERENCES
- El-Serag, H.B., 2012. Epidemiology of viral hepatitis and hepatocellular carcinoma. Gastroenterology, 142: 1264-1273.
CrossRefDirect Link - Abdel Aziz, M.T., H.M. Atta, S. Mahfouz, H.H. Fouad and N.K. Roshdy et al., 2007. Therapeutic potential of bone marrow-derived mesenchymal stem cells on experimental liver fibrosis. Clin. Biochem., 40: 893-899.
CrossRefDirect Link - Jiang, Y., B.N. Jahagirdar, R.L. Reinhardt, R.E. Schwartz and C.D. Keene et al., 2002. Pluripotency of mesenchymal stem cells derived from adult marrow. Nature, 418: 41-49.
CrossRefDirect Link - Phinney, D.G. and D.J. Prockop, 2007. Concise review: Mesenchymal stem/multipotent stromal cells: The state of transdifferentiation and modes of tissue repair-current views. Stem Cells, 25: 2896-2902.
CrossRefDirect Link - Santangelo, L., C. Battistelli, C. Montaldo, F. Citarella, R. Strippoli and C. Cicchini, 2017. Functional roles and therapeutic applications of exosomes in hepatocellular carcinoma. BioMed Res. Int., Vol. 2017.
CrossRef - Lee, Y., S. El Andaloussi and M.J.A. Wood, 2012. Exosomes and microvesicles: Extracellular vesicles for genetic information transfer and gene therapy. Hum. Mol. Genet., 21: R125-R134.
CrossRefDirect Link - Van Balkom, B.W., T. Pisitkun, M.C. Verhaar and M.A. Knepper, 2011. Exosomes and the kidney: Prospects for diagnosis and therapy of renal diseases. Kidney Int., 80: 1138-1145.
CrossRefDirect Link - Ratajczak, M.Z. and J. Ratajczak, 2016. Horizontal transfer of RNA and proteins between cells by extracellular microvesicles: 14 years later. Clin. Trans. Med., Vol. 5.
CrossRef - Desrochers, L.M., M.A. Antonyak and R.A. Cerione, 2016. Extracellular vesicles: Satellites of information transfer in cancer and stem cell biology. Dev. Cell, 37: 301-309.
CrossRefDirect Link - Cai, S., X. Cheng, X. Pan and J. Li, 2017. Emerging role of exosomes in liver physiology and pathology. Hepatol. Res., 47: 194-203.
CrossRefDirect Link - Lou, G., Z. Chen, M. Zheng and Y. Liu, 2017. Mesenchymal stem cell-derived exosomes as a new therapeutic strategy for liver diseases. Exp. Mol. Med., Vol. 49.
CrossRefDirect Link - Sato, K., F. Meng, S. Glaser and G. Alpini, 2016. Exosomes in liver pathology. J. Hepatol., 65: 213-221.
CrossRefDirect Link - Shen, J., C.K. Huang, H. Yu, B. Shen and Y. Zhang et al., 2017. The role of exosomes in hepatitis, liver cirrhosis and hepatocellular carcinoma. J. Cell. Mol. Med., 21: 986-992.
CrossRefDirect Link - Nojima, H., C.M. Freeman, R.M. Schuster, L. Japtok and B. Kleuser et al., 2016. Hepatocyte exosomes mediate liver repair and regeneration via sphingosine-1-phosphate. J. Hepatol., 64: 60-68.
CrossRefDirect Link - Li, T., Y. Yan, B. Wang, H. Qian and X. Zhang et al., 2013. Exosomes derived from human umbilical cord mesenchymal stem cells alleviate liver fibrosis. Stem Cells Dev., 22: 845-854.
CrossRefDirect Link - Tan, C.Y., R.C. Lai, W. Wong, Y.Y. Dan, S.K. Lim and H.K. Ho, 2014. Mesenchymal stem cell-derived exosomes promote hepatic regeneration in drug-induced liver injury models. Stem Cell Res. Therapy, Vol. 5.
CrossRef - Arslan, F., R.C. Lai, M.B. Smeets, L. Akeroyd and A. Choo et al., 2013. Mesenchymal stem cell-derived exosomes increase ATP levels, decrease oxidative stress and activate PI3K/Akt pathway to enhance myocardial viability and prevent adverse remodeling after myocardial ischemia/reperfusion injury. Stem Cell Res., 10: 301-312.
CrossRefDirect Link - Soleimani, M. and S. Nadri, 2008. A protocol for isolation and culture of mesenchymal stem cells from mouse bone marrow. Nat. Protocols, 4: 102-106.
CrossRefDirect Link - Bruno, S., C. Grange, F. Collino, M.C. Deregibus and V. Cantaluppi et al., 2012. Microvesicles derived from mesenchymal stem cells enhance survival in a lethal model of acute kidney injury. PloS One, Vol. 7.
CrossRef - Weber, S.N., A. Bohner, D.H. Dapito, R.F. Schwabe and F. Lammert, 2016. TLR4 deficiency protects against hepatic fibrosis and Diethylnitrosamine-induced pre-carcinogenic liver injury in fibrotic liver. PloS One, Vol. 11.
CrossRef - El-Magd, M.A., W.S. Abdo, M. El-Maddaway, N.M. Nasr and R.A. Gaber et al., 2017. High doses of S-methylcysteine cause hypoxia-induced cardiomyocyte apoptosis accompanied by engulfment of mitochondaria by nucleus. Biomed. Pharmacother., 94: 589-597.
CrossRefDirect Link - Livak, K.J. and T.D. Schmittgen, 2001. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method. Methods, 25: 402-408.
CrossRefDirect Link - Karin, M. and H. Clevers, 2016. Reparative inflammation takes charge of tissue regeneration. Nature, 529: 307-315.
CrossRefDirect Link - Jiang, W., Y. Tan, M. Cai, T. Zhao and F. Mao et al., 2018. Human umbilical cord MSC-derived exosomes suppress the development of CCl4-induced liver injury through antioxidant effect. Stem Cell. Int., Vol. 2018.
CrossRef - Tamura, R., S. Uemoto and Y. Tabata, 2016. Immunosuppressive effect of mesenchymal stem cell-derived exosomes on a concanavalin A-induced liver injury model. Inflammation Regeneration, Vol. 36.
CrossRef - Alzahrani, F.A., M.A. El-Magd, A. Abdelfattah-Hassan, A.A. Saleh, I.M. Saadeldin, E.S. El-Shetry, A.A. Badawy and S. Al-Karim, 2018. Potential effect of exosomes derived from cancer stem cells and MSCs on progression of DEN-induced HCC in rats. Stem Cell. Int., Vol. 2018.
CrossRefDirect Link