
Rekha Pathak
Division of Surgery,Indian Veterinary Research Institute, Izatnagar, Bareilly (UP), 243122, India
Amarpal
Division of Surgery,Indian Veterinary Research Institute, Izatnagar, Bareilly (UP), 243122, India
P. Tamilmahan
Division of Surgery,Indian Veterinary Research Institute, Izatnagar, Bareilly (UP), 243122, India
Kuldeep Dhama
Division of Pathology, Indian Veterinary Research Institute, Izatnagar, Bareilly (UP), 243122, India
Netrapal Sharma
Division of Surgery,Indian Veterinary Research Institute, Izatnagar, Bareilly (UP), 243122, India
International Journal of Pharmacology
Year: 2014 | Volume: 10 | Issue: 8 | Page No.: 440-450







ABSTRACT
The ability of the osteoblasts to lay the extracellular matrix and mineralize determines the efficiency of bone formation. Vitamin D injections are used to enhance bone mineralization in clinical cases. The present study was designed to evaluate the in vitro efficacy of Vitamin D on the osteoblastic activity in both fetal and adult osteoblasts. The long bone was collected from fetal and adult rabbits and cultured in Dulbecco’s Modified Eagle’s Medium (DMEM) containing 10% fetal bovine serum and antibiotics. The passaged cells, after attaining more than 80% confluence were induced for mineralization through osteogenic media with or without 1, 25-dihydroxyvitamin D3 (Vit D3). Fetal and adult cells were harvested after 7, 14, 21, 28 days for observation of osteoblastic differentiation and mineralization. Relative transcripts abundance of mRNA of alkaline phosphatase (ALP) and osteocalcin (OCN) were measured to assess the osteoblastic activity using quantitative real-time polymerase chain reaction (RT-qPCR) at different time intervals. The mineral accumulation was assessed by Von-Kossa staining. Fetal osteoblasts compared to adult osteoblasts showed a marked significant increase in mineralization on addition of Vit D. However, the adult osteoblasts reflected transcript abundance of ALP only after 14th day and OCN genes after 21 days post induction whereas fetal osteoblasts showed the same by 7th day post induction. This reflects a great therapeutic potential of fetal osteoblasts along with Vit D3 in bone regeneration. Further studies are required to correlate the influence of Vit D3 on ALP/OCN expression and also whether there is a comparative increase in the receptors of Vit D3 and osteocalcin protein in fetal cells as compared to adult osteoblasts.
PDF Abstract XML References Citation
Received: August 03, 2014;
Accepted: October 12, 2014;
Published: November 28, 2014
How to cite this article
Rekha Pathak, Amarpal, P. Tamilmahan, Kuldeep Dhama and Netrapal Sharma, 2014. Evaluation of in vitro Efficacy of Vitamin D3 on the Osteogenic Differentiation and Mineralization Capabilities of Fetal and Adult Osteoblasts of Rabbit Reflects Therapeutic Potential. International Journal of Pharmacology, 10: 440-450.
DOI: 10.3923/ijp.2014.440.450
URL: https://scialert.net/abstract/?doi=ijp.2014.440.450
DOI: 10.3923/ijp.2014.440.450
URL: https://scialert.net/abstract/?doi=ijp.2014.440.450
INTRODUCTION
The active hormonal form of vitamin D, calcitriol (1, 25-dihydroxyvitamin D3, or 1,25 D3) controls calcium homeostasis and modulates osteoblastic proliferation and differentiation (Hendy et al., 2006). It is also reported that Vit D3 promotes the early differentiation of progenitor cells to osteoblastic lineage and it decreases apoptosis in osteoblast like cells (Shalhoub et al., 1998). Also, Vit D3 is used clinically for the prevention and treatment of osteoporosis (Montero-Odasso and Duque, 2005). Vit D3 receptor is expressed in osteoblasts and Vit D3 modifies gene expression of various osteoblast differentiation and mineralization-related genes, such as alkaline phosphatase, osteocalcin and osteopontin (Van de Peppel and van Leeuwen, 2014). Molecular events involved in the development of fetal skeleton and adult fracture repair involves an orchestra of events which are precisely coordinated and mediate chondrogenesis, osteogenesis, angiogenesis and bone remodeling (Miclau et al., 2005). The models of cell culture are thought to recapitulate the events that occur in osteogenesis and bone repair. The various parameters by which we can assess the extent of mineralization are levels of alkaline phosphatase (early osteoblastic marker) and osteocalcin (Bharadwaj et al., 2009; Hoemanna et al., 2009). Alkaline phosphatase (ALP) is a membrane-bound enzyme abundant early in bone formation and increased ALP levels correlate with increased bone formation (Walsh et al., 2000). The ALP is a key enzyme involved in the process of bone formation and osteoid mineralization (Fernandez et al., 2014). The major non-collagenous protein of adult bone is osteocalcin (Hauschka and Reid, 1978). Osteocalcin is a bone specific protein (Beg et al., 2014). However, osteocalcin (OCN) is said to be a late stage marker for mature osteoblasts (Liu et al., 2003). The in vitro calcification abilities of osteoblast have also been well demonstrated (Hasegawa et al., 2008). There are different reports on the adult and the fetal osteoblast cells. Adult osteoblasts, even though widely characterized have typically limited expansion and are prone to dedifferentiation, mostly due to donor age-related changes (Evans et al., 1990; Katzburg et al., 1999). The fetal osteoblast cells on the other hand are immature cells with intrinsic ability to generate various mature cells that have remarkable viability and proliferative capacity. Dexamethasone containing Osteogenic Medium (OM) is normally used for osteoblastic differentiation of stem cells which induces cell growth, osteoblast maturation and in vitro mineralization in the presence of β-glycero phosphate and ascorbic acid (Fromigue et al., 1997). The cell therapy for the osteogenesis is very upcoming and promising in recent advances of orthopedics with varying opinions on the fetal and adult cell therapy. However, the in vitro studies regarding comparison of the behavior of fetal and adult osteoblast cells as regards to Vit D3 supplementation in osteogenic medium has not been conducted and investigated so far. Therefore, the present study was designed to evaluate the in vitro efficacy of Vitamin D3 on the osteogenic differentiation and mineralization capabilities of fetal and adult osteoblasts of rabbit.
MATERIALS AND METHODS
Sample collection: The present study was conducted in Division of Surgery, Indian Veterinary Research Institute, Izatnagar, Uttar Pradesh, India. Permission from the Institute Animal Ethics Committee (IAEC) was taken for the in vivo and in vitro studies. All the chemicals used in this study were procured from Sigma (St Louis, MO, USA), unless otherwise indicated.
For fetal osteoblast isolation, two full terms pregnant (28-30 days) New Zealand white rabbits of 6-8 month of age and 2-3 kg of body weight were utilized for the collection of fetuses’ long and calvarial bones after caesarian and put in sterilized Phosphate Buffered Saline (PBS) supplemented with antibiotics. The adult osteoblasts were isolated from radius bone piece from two six months old adult New Zealand white rabbits weighing 1.5-1.8 kg. The health of each rabbit was monitored for 2 weeks before the start of the experiment. All the operations were conducted in sterile manner.
Isolation and culture of fetal and adult osteoblast cells Fetus osteoblast: The isolation, culture, expansion and characterization of the osteoblasts were done by the method Cao et al. (2006) and Pathak et al. (2013).
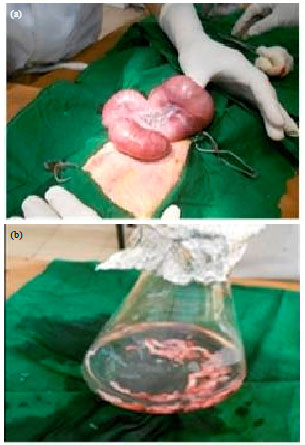 | |
| Fig. 1(a-b): | (a) Caesarean section in the female rabbit to remove the fetuses and collection of fetal bone, (b) Bone pieces collected from adult rabbit |
Fetuses were collected from a healthy pregnant New Zealand White rabbit after caesarean section under general anesthesia induced with intramuscular injection of xylazine@ 6 mg kg-1 followed by 10 min later by ketamine@ 60 mg kg-1. Fetuses were euthanized by intracerebral injection of xylazine (0.1 mL) and collected in sterile 1% PBS. Calvarial and long bones were harvested and washed with 1% PBS containing three times the usual concentration of antibiotics i.e., penicillin (100 U mL-1) and streptomycin (100 μg mL-1) (Fig. 1a). All the adhering soft tissues were stripped off and the bone was sectioned into small pieces. After three more washings in antibiotic solution, they were subjected to the enzymatic digestion in 0.25% trypsin at 37°C for 15 min by keeping on a magnetic stirrer. The bone pieces were washed in a culture medium without serum after trypsinization procedure and then dipped in culture medium (Dulbecco’s Modified Eagle’s Medium, DMEM) (Invitrogen, USA) with 20% fetal bovine serum (FBS) (Gibco, USA) and they were laid equally in T-25 culture flasks and incubated in a humidified atmosphere with 5% CO2 at 37°C for overnight.
 | |
| Fig. 2(a-h): | Primary isolation and culture cells from bone tissue of (a-d) Fetal and (e-h) Adult osteoblasts: (a, e) Cells migrated from bone pieces after 3 days of culture, (b, c, f, g) Cells with different shapes and protuberances showing triangular, short spindle, or polygonal shapes and (d, h) Cells ranked radially and forming nearly confluent cell layers after 11 days of culture in fetal osteoblast and 14 days of culture in adult osteoblast (Original micrographs were taken at x200) |
Normal culture medium, containing DMEM supplemented with 10% FBS, penicillin (100 U mL-1) and streptomycin (100 μg mL-1), was added to each flask after overnight incubation. The cells were seen to creep out after 72 h time from the bone pieces. Media was changed for at least twice a week. When 80-90% confluence was reached after 10 days, cells were transferred to new T-25 culture flask by using trypsinization method Fig. 2(a-d). The cells were passage to be seeded on glass cover slips coated with poly-d-lysine in 6 well plates at a cell density of 2x104. The purification of osteoblasts was continuously attempted by changing medium once a week.
Adult osteoblast: Similarly, the adult osteoblasts were cultured from the piece of radii from two rabbits after inducing general anesthesia by injecting xylazine at 6 mg kg-1, after 10 min ketamine at 60 mg kg-1 intramuscular (Amarpal et al., 2010). The left forelimb of the rabbit was shaved and aseptically prepared. After sedation, 2-3 cm long incision was made on the medial side of the radius, muscles were separated and radius bone was exposed. By using hacksaw, a 10 mm long radius bone with the ulna intact was removed and stored in Hank’s Balanced Salt Solutions (HBSS) (Sigma-Aldrich, USA) containing penicillin-streptomycin (MP Biomedicals, USA) (Fig. 1b). Created bone defect was filled with a non-antigenic bone graft prepared in the laboratory as a part of other experiment. Muscles were sutured with catgut No. 3-0 and skin sutured with nylon (No. 0). The adhering soft tissues and marrow were completely removed and bone was sectioned into small pieces. After three more washes with HBSS containing penicillin- streptomycin, the bone pieces were digested for 15 min at 37°C with 0.25% trypsin while being shaken with shaker. The digest was discarded in order to avoid fibroblast contamination. The bone pieces were washed repeatedly in a culture medium containing DMEM without serum and they were cultured in a plastic culture flask (T-25) containing DMEM with 100 U mL-1 penicillin, 100 μg mL-1 streptomycin, anti-fungal and 10% FBS. As the bone pieces floated on the culture medium easily, the flask was inverted and incubated in a humidified atmosphere with CO2 incubator at 37°C. This was followed by adding the adequate normal culture medium to the flask and then the cultures were replaced to culture sequentially. The medium change was carried out twice a week. After 5 days, cells were observed emerging from the bone pieces. The cell layers around the bone tissue became 70-80% confluent after approximately 11 days Fig. 2(e-h). The cells were then harvested by digestion with 0.05% trypsin-EDTA, centrifuged at 1200 rpm for 5 min, re-suspended in DMEM supplemented with 10% FBS, 100 U mL-1 penicillin, 100 μg mL-1 streptomycin and plated at a density of approximately 1x104 cells cm-2. Cells were cultured to full confluence before a second harvesting and again plated as per the procedure described above.
Geimsa staining: Giemsa stain, a polychromatic stain, was used to study the morphology of cells (Li et al., 2009). The cover slips with cells after removing culture medium, were washed with PBS. Cells were fixed in 70% ethanol (v/v) for 15 min and stained with a pre-mixed Giemsa staining solution for 30 min at room temperature. The stained cells were rinsed with tap water, air dried and examined under phase contrast microscope.
Alkaline phosphatase (ALP) activity: Cells were seeded at 1x104 in 6 well plates on cover slips and incubated for 24 h at 37°C in a humidified, 5% CO2 atmosphere in complete culture medium. Analysis for ALP activity was done in culture from day 3 using p-nitrophenol as substrate N1891 (Sigma Aldrich, USA). Measurements were performed in triplicates for specific time point.
Osteogenic induction: After primary culture in the control medium (DMEM+10% FBS+100 U mL-1 Penicillin 100 μg mL-1 streptomycin) and expansion for two passages, the cells were trypsinized and counted via cell counter using trypan blue to measure viable cells and replated onto 6-well tissue culture plates at a density of 4x104 cells per well. The cells were incubated in the control medium for 1 day to adhere them to the plates and then placed in Osteogenic Medium (OM) containing 10 mM β-glycerophosphate, 50 mg mL-1 L-ascorbic acid and 10 nM dexamethasone (Dex) (Sigma-Aldrich, USA). The other 3 wells were supplemented with osteogenic medium plus 10 nM 1, 25 Dihydroxy cholecalciferol (Vit D3) (Sigma-Aldrich, USA). Medium was changed after every third day. Osteogenic differentiation was assessed by Von Kossa staining and expression of osteoblast associated marker genes such as osteocalcin (OCN) and alkaline phosphatase (ALP) were confirmed by reverse transcription-polymerase chain reaction (RT-PCR) on 7, 14, 21 and 28 days after initial osteogenic induction. The whole experiment was performed in triplicates.
Von Kossa staining: The presence of mineralized nodules was observed by Von Kossa silver nitrate stain (Wang et al., 2006). For staining, medium was removed and cells were fixed in cold methanol for 15-20 min. After rinsing, the fixed plate was incubated with 5% silver nitrate solution under 1 h UV exposure. Mineralized nodules were seen as dark brown to black spots on observed under phase contrast microscope (CKX 41, Olympus, Japan).
RNA extraction and reverse transcription: Using the Trizol reagent-RT, the total RNA was isolated from both groups of cultured fetal and adult osteoblast cells. Cells after lysed in trizol solution were centrifuged at 15,000 rpm for 15 min at 4°C. RNA was precipitated using isopropyl alcohol on the upper aqueous phase. The RNA pellet was washed in 70% alcohol and dissolved finally in nuclease free water. Treatment with DNAase I was performed for removal of any DNA contamination as per the manufacturer’s protocol (Quigen GmbH Hilden, Germany). Quality and concentration of isolated mRNA was assessed by Nano-drop (Thermo Scientific, USA). Reverse Transcription (RT) was carried out using first strand cDNA synthesis kit (Fermentas, Maryland, USA) according to manufacturer’s instructions. Briefly, cDNA was synthesized in a total of 20 μL reaction volume using 450 ng of RNA isolated from both the groups of OM and ‘OM with Vitamin D3 (OM+Vit D3) of osteoblasts. Reaction was reverse-transcribed using Molony-Murine Leukemia Virus Transcriptase (MMLV-RT) (Fermentas, Maryland, USA) by incubating at 70°C for 5 min followed by incubation at 25°C for 5 min, 42°C for 60 min and finally reaction was stopped by incubating the reaction for 10 min at 70°C.
| Table 1: | List of primers used |
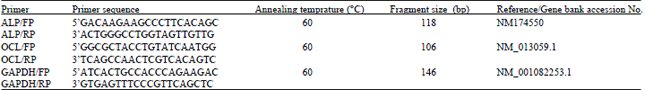 | |
The quality of cDNA was assessed by an amplification reaction for a housekeeping gene of glyceraldehyde-3-phosphate dehydrogenase (GAPDH).
Primer design for SYBR Green based qPCR: A set of primers for real time analysis of OCN and ALP gene were designed from the conserved coding region of Human and Rat sequences, respectively. OCN and ALP gene specific and housekeeping glyceraldehyde-3-phosphate dehydrogenase (GAPDH) primers for real time were synthesized using primer 3 (v. 0.4.0) through slight adjustment in default parameters set up, quality was checked on primer select software (DNA STAR, USA). Length of the amplified product was 106, 118 and 146 bp for OCN, ALP and GAPDH, respectively. The primer sequences used are listed in Table 1.
Real-time PCR: SYBR Green based real time was performed for OCN and ALP with 450 ng of initial template in each reaction of quantitative PCR (qPCR) (Stratagene MX3000P; CA, USA). Each qPCR reaction was prepared using MaximaTM SYBR Green/ROX qPCR master mix (2X) (Fermentas, Maryland USA). The procedure was optimized with regards to primer concentrations, denaturation/extension temperature and reaction efficiency. Finally, the optimized reaction was carried out in triplicates along with the housekeeping GAPDH. The relative expression of OCN and ALP were analyzed using 450 ng initial template in both the groups of osteoblast cultures. The optimized reaction was carried out in a final reaction volume of 25 μL containing 1 μL (0.5 μmol L-1 concentrations) of each forward and reverse primer, 3 μL of cDNA, 7.5 μL-1 of nuclease free water and 12.5 μL-1 SYBR Green qPCR master mix. Thermal profile used for amplification of all replicates consisted of an initial denaturing cycle of 10 min at 95°C; 40 cycles of PCR (95°C for 30 sec, 60°C for 20 sec and 72°C for 25 sec) and melting curve profile (95°C for 1 min followed by 60°C for 30 sec and 95°C for 30 sec) was set for fluorescence acquisition and reaction specificity. Fluorescence signal from primer dimers or other non-specific amplification products distinguished from the specific product through their lower Tm was also monitored. The Ct values were recorded and average Ct values from three replicates were taken into real time data analysis.
Statistical analysis: Relative expression was analyzed by method described by Pfaffl (2001). QPCR data was analyzed using one way ANOVA by Graph Pad InStat Software (Peter Russell, Royal Veterinary College, London) at 0.05% level of significance. Differences with p<0.05 were considered statistically significant. The relative abundance of OCN and ALP transcripts expressed was for 450 ng of initial template mRNA.
RESULTS
The morphology of fetal and adult osteoblast cells collected from rabbits were confirmed by Giemsa and ALP staining, showed almost the same morphology being triangular and enlarged. Triangular and spindle shaped cells with intense pink cytoplasm was observed in the culture. The nucleus was at centre or the margin with round or elliptical shape (Fig. 3). The cells which were positive for ALP activity (more than 95%) were examined for the presence of blue granules since the stain used was fast blue RR salt (Fig. 3). Cells started creeping from explants of adult and fetal at same time but fetal cells became confluent more rapidly (10-11 days) in comparison to adult cells (14-15 days). The 7-28 day culture period in media supplemented with osteogenic supplements OM and OM+ Vit D3 resulted in osteoblastic differentiation, as demonstrated by culture layer Von Kossa staining for mineralization (Fig. 4) and osteoblast specific gene expression by RT-qPCR (Fig. 5, 6). To confirm the RNA and cDNA quality, amplified product of housekeeping gene GAPDH was checked on agarose gel, a solitary intact band was observed under UV transilluminator confirming its quality (Fig. 7, 8). Cells cultured with OM and OM+Vit D, clustered and formed distinct mineralization nodule starting from 7th day of induction in both fetal and adult cells. Von Kossa staining of the mineralization activity in the cell layer was found in substantial amounts by 7th day culture, increased by 14th day and 21st day and remained at maximal levels at 28th day cultured cells (Fig. 4).
Expression of ALP and OCN: To investigate osteogenic capabilities of fetal and adult cells, OCN and ALP gene markers for osteoblast function were determined. The ALP activity was not seen in adult osteoblasts until 14th day post induction whereas it started to express at 7th day in the fetal osteoblasts.
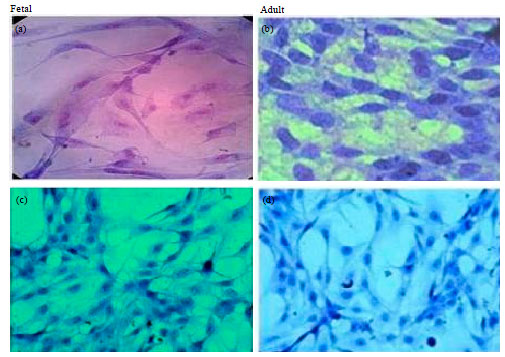 | |
| Fig. 3(a-d): | Fetal and adult osteoblast cells stained with (a, b) Giemsa staining for cell morphology and (c, d) ALP staining for alkaline phosphatase activity |
 | |
| Fig. 4(a-p): | Continuous monitoring of mineralized nodules formation in fetal osteoblast cells; A series of Von Kossa stain images were obtained from the culture incubated with OM alone and OM+Vit D in (a-h) Adult and (i-p) Fetal osteoblast cultures respectively, at different time intervals (7, 14, 21 and 28 days). Note the increased intensity of mineralization in fetal osteoblasts. The arrow indicates mineralized osteoblast |
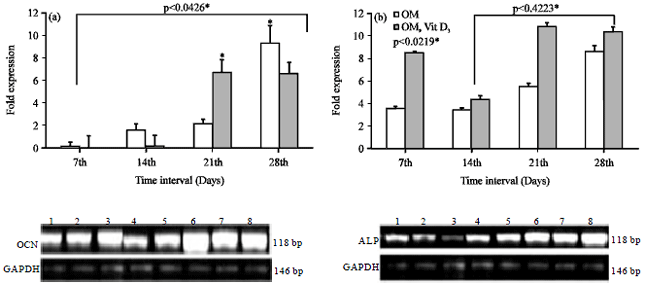 | |
| Fig. 5(a-b): | (a) Fetal osteoblast, osteogenic mineralization in fetal osteoblast cells and alkaline phosphatase and Osteocalcin expression after 7, 14, 21 and 28 days incubation, two different mineralization media (only DEX and DEX+Vit-D) were used. Gel show expression pattern, loading; 1, 2 is 7th day; 3, 4 is 14th day; 5, 6 is 21st day and 7 and 8 is 28th day of mineralization. Real time data showed no significant difference (p<0.05) in the relative transcript abundance of ALP gene in osteoblasts either cultured in OM or OM+Vit D3 at different time intervals. Gel indicates expression of (b) ALP in different time of interval with significant difference (*p<0.0222) in the relative transcript abundance of OCN gene in osteoblasts either cultured in OM or OM+Vit D3 at different time intervals. GAPDH as control |
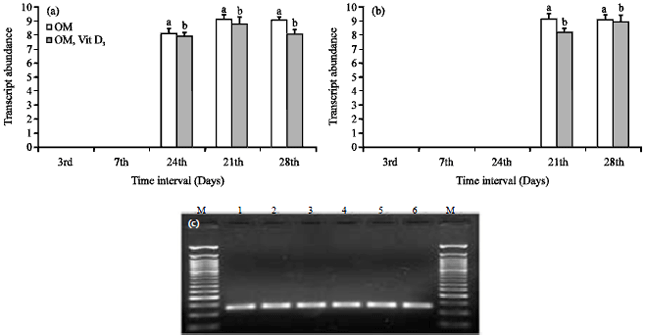 | |
| Fig. 6(a-c): | Adult osteoblast, relative transcript abundance of (a) Alkaline phosphatase, (b) Osteocalcin and in rabbit osteoblast cultured for 28 days in osteogenic medium (OM) and osteogenic medium containing Vitamin D3 (OM+Vit D3). Transcripts of ALP were expressed from 14th day onwards while OCN transcripts were observed at 21st and 28th day in osteoblast culture. Real time data showed no significant difference (p<0.05) in the relative transcript abundance of ALP and OCN gene in osteoblasts either cultured in OM or OM+Vit D3 at different time intervals and (c) Confirmation of cDNA quality through PCR amplification of GAPDH (146 bp) gene in rabbit osteoblast |
 | |
| Fig. 7(a-b): | Isolation of RNA from osteoblast cells, (a) Before DNase treatment and (b) After DNase treatment |
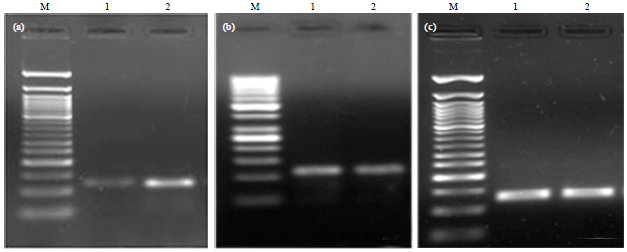 | |
| Fig. 8(a-c): | PCR amplification of (a) OCN (106 bp), (b) ALP (118 bp) and (c) Confirmation of cDNA quality through PCR amplification of GAPDH (146 bp) gene. Lane M, 50 bp+molecular weight marker; Lane 1: Adult and and Lane 2: Fetal respective amplicons in rabbit osteoblasts cultured in vitro |
The activity in fetal osteoblasts was significantly higher with Vit D3+OM as compared to the OM alone. The ALP activity in adult cells increased on 14th day and Vit D3 had no significant bearing as in fetal cells. In both the cell culture types, the activity of ALP kept on increasing up to the observation time. The expression of OCN was observed from 7th day onwards in fetal cells whereas in adult cell it was 21st day onward. Real time data showed significant difference (p<0.0222) in the relative transcript abundance of OCN gene in fetal osteoblasts either cultured in OM or OM+Vit D3 at different time intervals. Gel indicated expression of OCN in different time of interval and GAPDH used as control (Fig. 5, 6).
DISCUSSION
In this study, as a means of cell characterization, firstly fetal osteoblasts were compared to rabbit adult osteoblasts for their expression of surface and intracellular markers. The data presented herein demonstrated that adult rabbit cells isolated via standard biopsy and fetal cells by fetuses collection procedures can be used to produce osteoblast cells that may be a suitable model for in vitro investigations. Cells isolated from fetal and adult rabbit bone provided an excellent source of proliferative osteoprogenitor cells that can be induced to differentiate osteoblasts. Studies concerning the development of osteoblast phenotype, from osteoprogenitor cells to osteocytes embedded in the extracellular matrix, suggest a temporal sequence of events involving active cell proliferation, expression of osteoblast markers and matrix mineralization (Cao et al., 2006). Fetal osteoblasts have previously been shown to proliferate, differentiate, mineralize their extracellular matrix (Montjovent et al., 2004) and respond to both chemical and surface stimuli (Montjovent et al., 2004; Christodoulou et al., 2005, 2006; Tsigkou et al., 2007). The present study was designed to investigate how continuous treatment with OM and OM+Vit D3 could increase the process of fetal and adult osteoblast differentiation and matrix mineralization. This was demonstrated by the Von Kossa stain and expression of markers characteristic of the osteoblast phenotype, such as ALP activity and OCN gene expression. Fetal osteoblasts undergo programmed differentiation to synthesize OCN following stimulation with Vitamin D3 (Harris et al., 1995). The identification of a calcified Extra Cellular Matrix (ECM) in culture layers is normally assessed by Von Kossa staining (Jiang et al., 2005). It was found that calcification nodules appeared as black regions within the cell layers. Consistent with osteogenesis, several black nodules, indicative of a calcified ECM, were identified in rabbit osteoblast cell layers. Previous studies with von Kossa staining and assessment of ALP activity showed that mineralization occurred in the center of the colonies due to the effect of OM (Li et al., 1996). The OM thus had a significant effect on the commitment of cells to pursue the osteoblastic phenotype. Although in the present study the intensity of staining increased with the time interval more in fetal osteoblasts than adult osteoblast. However, it was observed that the intensity of staining was more when Vit D3 was added to the OM. 1,25 D3 is known to stimulate mineralization of human osteoblasts in vitro and recently it was shown that 1,25 D3 induces mineralization via effects in the period preceding mineralization during the pre-mineralization period. Vit D3 receptor is expressed in osteoblasts and Vit D3 modifies gene expression of various osteoblast differentiation and mineralization-related genes, such as alkaline phosphatase, osteocalcin and osteopontin (Van de Peppel and van Leeuwen, 2014).
Based on numerous previous studies and techniques, it is known that ALP expression rises as osteoblasts mature and then declines as osteoid becomes heavily mineralized, while OCN expression is acquired latest and essentially is diagnostic of post-proliferative osteoblasts (Malaval et al., 1999). ALP plays a significant role in facilitating bone mineralization and is an early marker of osteoblast differentiation (Barroga et al., 2000). In this study, ALP gene expression was significantly increased at 14 days of post induction in both medium in adult cells cultured with OM with or without Vit D3. Addition of Vit D3 did not much influence the increase in the ALP expression. However, in fetal cells the expression not only started at 7th day onwards but addition of Vit D3 to OM had a significant (p<0.02) impact on increase in ALP expression which shows that fetal osteoblasts either have more Vit D3 receptors or an increased sensitivity to Vit D3. ALP is a membrane-bound enzyme abundant early in bone formation and increased ALP levels correlate with increased bone formation (Walsh et al., 2000). Although, ALP expression is strongly upregulated in osteogenic tissues, it is expressed in several non-osteogenic cell types and tissues such as cartilage, liver and kidney (Weiss et al., 1988). Therefore, ALP expression, as an indicator of osteogenesis (Lee, 2014), should be used in conjunction with other osteogenesis-associated markers.
To further verify the osteogenic capacity of fetal and adult rabbit osteoblast cells, the expression of osteoblast-related genes including OCN was also examined by RT-PCR assays. Clinically, OCN as a marker protein of mature osteoblast is routinely used as a serum marker for bone formation as well as for bone turn over (Delmas et al., 1986). Both Vit D3 and OM induced differentiated adult rabbit osteoblast cells expressed OCN as late as 21st day post induction which negates the role of supplemented Vit D3 on OCN expression but in fetal osteoblast it was expressed from 7th day in both OM and OM+Vit D3 group. The addition of Vit D3 boosts the OCN expression on 21st day however by 28th day there was an increase in OCN expression both in OM and Vit D3 where OM group significantly (p< 0.042) expressed more OCN than Vit D3 added to OM. This needs a further investigation. The OCN is a late bone marker that is secreted only by osteoblasts and signals terminal osteoblastic differentiation and represents matrix maturation (Aubin and Triffitt, 2002). The early expression of OCN in fetal osteoblasts shows rapid differentiation and matrix maturation than adult osteoblasts. Further, the addition of Vit D3 does not bear any significant increase or decrease in the adult osteoblastic marker whereas it did show significant increase in the fetal ALP and OCN.
CONCLUSION
Adult osteoblast cells of rabbit did not show significant mineralization on adding of Vit D3 to the Osteogenic Medium (OM) whereas fetal osteoblast showed a marked significant increase in mineralization with Vit D3 treatment. This reflects a great therapeutic potential of fetal osteoblasts along with Vit D3 in bone regeneration. Further studies are required to correlate the influence of Vit D3 on ALP/OCN expression and also whether there is a comparative increase in the receptors of Vit D3/osteocalcin protein in fetal cells as compared to adult osteoblasts.
ACKNOWLEDGEMENT
This research study was financially supported from Department of Biotechnology (DBT), New Delhi, Government of India and carried out at Indian Veterinary Research Institute, Izatnagar.
REFERENCES
- Amarpal, P. Kinjavdekar, H.P. Aithal, A.M. Pawde, J. Singh and R. Udehiya, 2010. Evaluation of xylazine, acepromazine and medetomidine with ketamine for general anaesthesia in rabbits. Scand. J. Lab. Anim. Sci., 37: 223-229.
Direct Link - Barroga, E.F., T. Kadosawa and M. Okumura, 2000. Influence of vitamin D and retinoids on the induction of functional differentiation in vitro of canine osteosarcoma clonal cells. Vet. J., 159: 186-193.
CrossRefDirect Link - Beg, M., N. Akhtar, M.F. Alam, I. Rizvi, J. Ahmad and A Gupta, 2014. Vitamin D status and serum osteocalcin levels in post-menopausal osteoporosis: Effect of bisphosphonate therapy. J. Indian Acad. Clin. Med., 15: 172-176.
Direct Link - Bharadwaj, S., A.G.T. Naidu, G.V. Betageri, N.V. Prasadarao and A.S. Naidu, 2009. Milk ribonuclease-enriched lactoferrin induces positive effects on bone turnover markers in postmenopausal women. Osteoporos Int., 20: 1603-1611.
CrossRefDirect Link - Christodoulou, I., L.D. Buttery, P. Saravanapavan, G. Tai, L.L. Hench and J.M. Polak, 2005. Dose- and time-dependent effect of bioactive gel-glass ionic-dissolution products on human fetal osteoblast-specific gene expression. J. Biomed. Mater. Res. B: Applied Biomater., 74: 529-537.
CrossRefDirect Link - Christodoulou, I., L.D. Buttery, G. Tai, L.L. Hench and J.M. Polak, 2006. Characterization of human fetal osteoblasts by microarray analysis following stimulation with 58S bioactive gel-glass ionic dissolution products. J. Biomed. Mater. Res. B: Applied Biomater., 77: 431-446.
CrossRef - Delmas, P.D., B. Demiaux, L. Malaval, M.C. Chapuy, C. Edouard and P.J. Meunier, 1986. Serum bone gamma carboxyglutamic acid-containing protein in primary hyperparathyroidism and in malignant hypercalcemia. Comparison with bone histomorphometry. J. Clin. Invest., 77: 985-991.
CrossRefDirect Link - Fromigue, O., P.J. Marie and A. Lomri, 1997. Differential effects of transforming growth factor β2, dexamethasone and 1,25-dihydroxyvitamin d on human bone marrow stromal cell. Cytokine, 9: 613-623.
CrossRefDirect Link - Fernandez, J.M., M.S. Molinuevo, A.D. McCarthy and A.M. Cortizo, 2014. Regarding: Strontium ranelate stimulates the activity of bone-specific alkaline phosphatase: Interaction with Zn2+ and Mg2+. Biometals, 27: 601-607.
CrossRefDirect Link - Hasegawa, Y., K. Shimada, N. Suzuki, T. Takayama and T. Kato et ak., 2008. The in vitro osteogenetic characteristics of primary osteoblastic cells from a rabbit calvarium. J. Oral Sci., 50: 427-434.
PubMed - Harris, S.A., R.J. Enger, B.L. Riggs and T.C. Spelsberg, 1995. Development and characterization of a conditionally immortalized human fetal osteoblastic cell line. J. Bone Miner. Res., 10: 178-186.
CrossRefDirect Link - Hendy, G.N., K.A. Hruska, S. Mathew and D. Goltzman, 2006. New insights into mineral and skeletal regulation by active forms of vitamin D. Kidney Int., 69: 218-223.
CrossRefPubMedDirect Link - Hoemanna, C.D., H. El-Gabalawy and M.D. McKee, 2009. In vitro osteogenesis assays: Influence of the primary cell source on alkaline phosphatase activity and mineralization. Pathologie Biologie, 57: 318-323.
CrossRefDirect Link - Van de Peppel, J. and J.P.T.M. van Leeuwen, 2014. Vitamin D and gene networks in human osteoblasts. Front. Physiol., Vol. 5.
CrossRefDirect Link - Jiang, X.Q., J. Chen, G. Sebastien, C. Chen, X. Zhang and Z. Zhang, 2005. The ectopic study of tissue-engineered bone with hBMP-4 gene modified bone marrow stromal cells in rabbits. Chin. Med. J., 118: 281-288.
PubMed - Katzburg, S., M. Lieberherr, A. Ornoy, B.Y. Klein, D. Hendel and D. Somjen, 1999. Isolation and hormonal responsiveness of primary cultures of human bone-derived cells: Gender and age differences. Bone, 25: 667-673.
CrossRefDirect Link - Li, I.W., S. Cheifetz, C.A. McCulloch, K.T. Sampath and J. Sodek, 1996. Effects of osteogenic protein-1 (OP-1, BMP-7) on bone matrix protein expression by fetal rat calvarial cells are differentiation stage specific. J. Cell. Physiol., 169: 115-125.
PubMed - Li, S.H., D.Z. Guo, B. Li, H.B. Yin, J.K. Li, J.M. Xiang and G.Z. Deng, 2009. The stimulatory effect of insulin-like growth factor-1 on the proliferation, differentiation and mineralisation of osteoblastic cells from Holstein cattle. Vet. J., 179: 430-436.
CrossRefDirect Link - Liu, F., L. Malaval and J.E. Aubin, 2003. Global amplification polymerase chain reaction reveals novel transitional stages during osteoprogenitor differentiation. J. Cell Sci., 116: 1787-1796.
Direct Link - Malaval, L., F. Liu, P. Roche and J.E. Aubin, 1999. Kinetics of osteoprogenitor proliferation and osteoblast differentiation in vitro. J. Cell. Biochem., 74: 616-627.
CrossRefDirect Link - Montjovent, M.O., N. Burri, S. Mark, E. Federici and C. Scaletta et al., 2004. Fetal bone cells for tissue engineering. Bone, 35: 1323-1333.
CrossRefDirect Link - Pfaffl, M.W., 2001. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res., 29: e45-e45.
CrossRefPubMedDirect Link - Pathak, R., Amarpal, A.K. Tiwari, N.P. Kurade and Amarnath, 2013. Isolation, expansion and characterization of rabbit fetal osteoblast cultured from rabbit calvarial and limb bones. J. Cell Tissue Res., 13: 3859-3864.
Direct Link - Shalhoub, V., F. Aslam, E. Breen, A. van Wijnen and R. Bortell et al., 1998. Multiple levels of steroid hormone-dependent control of osteocalcin during osteoblast differentiation: glucocorticoid regulation of basal and vitamin D stimulated gene expression. J. Cell. Biochem., 69: 154-168.
CrossRefDirect Link - Tsigkou, O., L.L. Hench, A.R. Boccaccini, J.M. Polak and M.M. Stevens, 2007. Enhanced differentiation and mineralization of human fetal osteoblasts on PDLLA containing Bioglass® composite films in the absence of osteogenic supplements. J. Biomed. Mater. Res. A, 80: 837-851.
CrossRefDirect Link - Walsh, S., C. Jefferiss, K. Stewart, G.R. Jordan, J. Screen and J.N. Beresford, 2000. Expression of the developmental markers STRO-1 and alkaline phosphatase in cultures of human marrow stromal cells: Regulation by Fibroblast Growth Factor (FGF)-2 and relationship to the expression of FGF receptors 1-4. Bone, 27: 185-195.
CrossRefDirect Link - Wang, J., S. Rao, J. Chu, X. Shen, D.N. Levasseur, T.W. Theunissen and S.H. Orkin, 2006. A protein interaction network for pluripotency of embryonic stem cells. Nature, 444: 364-368.
CrossRefDirect Link - Weiss, M.J., K. Ray, P.S. Henthorn, B. Lamb, T. Kadesch and H. Harris, 1988. Structure of the human liver/bone/kidney alkaline phosphatase gene. J. Biol. Chem., 263: 12002-12010.
PubMed - Montero-Odasso, M. and G. Duque, 2005. Vitamin D in the aging musculoskeletal system: An authentic strength preserving hormone. Mol. Aspects Med., 26: 203-219.
CrossRefPubMedDirect Link - Hauschka, P.V. and M.L. Reid, 1978. Vitamin D dependence of a calcium-binding protein containing gamma-carboxyglutamic acid in chicken bone. J. Biol. Chem., 253: 9063-9068.
PubMedDirect Link