
Mahir Kula
Department of Medical Pharmacology, Faculty of Cerrahpaa Medical, Istanbul University, Turkey
Gamze Tanriverdi
Department of Histology and Embryology, Faculty of Cerrahpaa Medical, Istanbul University, Turkey
Ersoy Oksuz
Department of Medical Pharmacology, Faculty of Cerrahpaa Medical, Istanbul University, Turkey
Ayhan Bilir
Department of Histology and Embryology, Faculty of Istanbul Medical, Istanbul University, Turkey
Andleeb Shahzadi
Department of Medical Pharmacology, Faculty of Cerrahpaa Medical, Istanbul University, Turkey
Zeliha Yazici
Department of Medical Pharmacology, Faculty of Cerrahpaa Medical, Istanbul University, Turkey
International Journal of Pharmacology
Year: 2014 | Volume: 10 | Issue: 5 | Page No.: 267-274







ABSTRACT
The present study was designed to investigate the combined effect of fotemustine, a nitrosureas antineoplastic agent along with simvastatin and/or dexamethasone on C6 glioblastoma. The C6 glioblastoma cells (1x106) were inoculated in rat brains. Ten days later, rats were treated with 10 mg kg-1 fotemustine, 3 mg kg-1 day-1 dexamethasone and 3 mg kg-1 day-1 simvastatin alone or with their combination. Monoclonal antibody Ki-67 was used to evaluate cell proliferation. The effects of these drug alone or in combination on fatty acid profile of C6 glioblastoma were determined using capillary gas chromatography. Dexamethasone±simvastatin decreased the tumor weight 36-46% (p = 0.001-0.029). The fotemustine+dexamethasone+simvastatin combination was more effective than the drugs given separately in inhibiting the growth of the tumor (33.9-58.5%, p = 0.000). In the fotemustine+dexamethasone+simvastatin group, the total fatty acid amount was high compared with control tumors, whereas low in the fotemustine+simvastatin group (31.95±1.91 and 16.11±1.96 μg mg-1, p = 0.021). The number of Ki-67 positive cell decreased compared with control tumors by drug therapy except dexamethasone alone. The decrease was greatest in the treatment with fotemustine+dexamethasone+simvastatin (26.5-51.8%, p = 0.000-0.020). These data show that, fotemustine+dexamethasone+simvastatin combination is more effective than fotemustine alone on inhibition of cell proliferation and tumor progression.
PDF Abstract XML References Citation
Received: March 18, 2014;
Accepted: June 25, 2014;
Published: August 21, 2014
How to cite this article
Mahir Kula, Gamze Tanriverdi, Ersoy Oksuz, Ayhan Bilir, Andleeb Shahzadi and Zeliha Yazici, 2014. Simvastatin and Dexamethasone Potentiate Antitumor Activity of Fotemustine. International Journal of Pharmacology, 10: 267-274.
DOI: 10.3923/ijp.2014.267.274
URL: https://scialert.net/abstract/?doi=ijp.2014.267.274
DOI: 10.3923/ijp.2014.267.274
URL: https://scialert.net/abstract/?doi=ijp.2014.267.274
INTRODUCTION
Fotemustine belongs to nitrosureas group an antineoplastic agents widely used in the treatment of malignant glioma and brain metastasis. Unfortunately, the systemic use of nitrosureas is associated with considerable dose dependent toxicity including myelosuppression, gastric dysfunction and central nervous system toxicity. Alternative treatment strategies such as drug combinations is taken into consideration to ensure higher efficacy in suppressing tumor proliferation while lowering toxic effects (Khil et al., 2000; Vogelhuber et al., 2002).
It has been shown that products of mevalonate pathway play a critical role in cell proliferation. For this reason, HMG CoA reductase inhibitors (simvastatin, pravastatin, atorvastatin, etc.) not only inhibit the biosynthesis of mevalonate but also promote apoptosis in both the normal and tumor cells by inhibiting cell growth signalisation, thus reducing cell proliferation (Baetta et al., 1997; Garcia-Roman et al., 2001).
Studies have shown that dexamethasone decreases vascular permeability of brain tumors in rats. Commonly used for management of cerebral edema in patient with brain tumors, glucocorticoids also have immunosuppressive function (Heiss et al., 1996; Wolff et al., 1993). On the other hand, it has been shown that statins prevent vascular endothelial cells from entering G1 phase, inhibiting cell proliferation and that dexamethasone can block this effect (Newton et al., 2003).
Cancer cell proliferation, angiogenesis and fatty acid metabolism play an important role in evaluating tumor growth and metastasis. Changes in fatty acid composition of tumor cells may affect the drug response of the tumor cell and the sensitivity against immune attacks, altering physical properties of membrane fluidity, permeability and transport systems, receptor binding and eicosanoid production (De Kruyff et al., 1973). Brain tissue compared to other tissues contains more long-chain poly-unsaturated fatty acids, especially docosahexanoic acid (DHA, C22:6n-3) (Moore et al., 1991). The DHA is one of the major components of excitable membranes. It regulates membrane fluidity and functions of integral proteins, possibly by intercepting acyl-chain bonding or by transmembrane interactions (Applegate and Glomset, 1986; Stubbs and Smith, 1984). Arachidonic acid (C20:4n-6) is a substrate located in cell membrane and synthesized from linoleic acid (C18:2n-6) is used for prostaglandin synthesis. It makes up 5-15% of fatty acids in phospholipids (Hanai et al., 1993). Arachidonic acid is a precursor for many biological active compounds such as prostaglandins, hydroxyeicosatetraenoic acid and leukotrienes (Moore et al., 1991; Wolfe, 1982). Prostaglandins, especially Prostaglandin E2 (PGE2) promote tumor growth by affecting host immunocytes (Kokoglu et al., 1998). Eicosapentaenoic acid (C20:5n-3, EPA) is also a precursor for prostaglandins, hydroxy acids and leukotrienes (Moore et al., 1991). Linoleic acid and arachidonic acid of n-6 series are associated with tumor proliferation (Broitman et al., 1977; Hanai et al., 1993; Rogers, 1983). On the contrary, it is known that n-3 series poly-unsaturated fatty acids (α-linolenic acid, C18:3n-3, EPA and DHA) suppress carcinogenesis and tumor metastasis (Corey et al., 1983; Culp et al., 1979; Hanai et al., 1993; Karmali et al., 1984; Mengeaud et al., 1992). However, poly-unsaturated fatty acids are more effective in promoting cancer progression than saturated fatty acids (Ip et al., 1985).
The Ki-67 is a protein associated with cell proliferation. This antigen is present on chromosome surface within the nucleus during interphase. Therefore, it is present in all the active phases of the cell cycle (G1, S, G2, mitosis) but is absent in resting cells (G0). So, it is a perfect marker for determining growing cells in a cell population. Marking and determining the Ki-67 positive tumor cells is widely used in evaluating the clinical course of the disease. For that purpose, anti-Ki-67 antibodies are used to evaluate cell proliferation (Scholzen and Gerdes, 2000).
In this study, we investigated the combined effect of fotemustine with simvastatin and/or dexamethasone on cell proliferation and tumor size of C6 glioblastoma in vivo. The effect of these drugs on fatty acid profile of C6 glioblastoma also has been investigated because changes in fatty acid composition of tumor cells may affect the drug response of the tumor cell and sensitivity against immune attacks, altering physical properties of the membrane fluidity, permeability and transport systems, receptor binding and eicosanoid production.
MATERIALS AND METHODS
Cell and cell culture: The C6 gliolastoma cell line was obtained from American type culture collection (Rockville, USA) and serial passages were made at the Histology and Embryology Department of Istanbul Univeristy, Faculty of Medical in modified Eagle medium of Dulbecco which contained 15% heat inactivated fetal calf serum, 0.2 mM glutamine, 50 mg mL-1 neomycin and 100 mg mL-1 streptomycin. Culture flasks were kept in electronic incubator (Sanyo) at 37°C under humidified atmosphere containing 5% CO2.
Implantation procedure: Cells were harvested via., 2 mL trypsin-EDTA solution C (Biological Industries, Israel) and centrifuged after the addition of 1.5 mL of F12 medium and fetal calf serum mixture at 1x103 rpm for three minutes. After removing the supernatant, pellet was resuspended with 2 mL of medium. Cell suspension was concentrated, so that 5 μ of any injection volume would contain 1x106 cells (Altinoz et al., 2001).
Tumor inoculation and drug treatment: The following experiment was approved by Experimental Animals Ethics Committee of Istanbul University. Six to eight week old male Wistar albino rats weighing about 250-300 g were anesthetized by i.p. 42 mg kg-1 ketamine and 0.1 mg kg-1 chlorpromazine. The C6 glioblastoma cells (1x106) were stereotaxically injected into the frontal lobe (coordinates, 2 mm anterior and 2 mm lateral to the bregma and at a depth of 4 mm) using 10 μL Hamilton syringe. Ten days after the inoculation, tumor-bearing rats were randomly divided into 8 groups of 10 animals each (5 rats for fatty acid analysis and other 5 rats for histological and immunohistological examination) and treated with fotemustine (Servier Laboratories) dexamethasone (DEVA Pharmaceutical Co., Turkey) and/or simvastatin (NOBEL Pharmaceutical Co., Turkey) as the following: Group 1: Untreated tumor-bearing control, Group 2: The 10 mg kg-1 i.p. fotemustine (single dose), Group 3: The 10 mg kg-1 i.p. fotemustine (single dose) and 3 mg kg-1 day-1 oral simvastatin, Group 4: The 3 mg kg-1 i.p. dexamethasone (single dose), Group 5: The 10 mg kg-1 i.p. fotemustine (single dose) and 3 mg kg-1 day-1 oral simvastatin, Group 6: The 10 mg kg-1 i.p. fotemustine (single dose) and 3 mg kg-1 day-1 i.p. dexamethasone, Group 7: The 3 mg kg-1 day-1 i.p. dexamethasone and 3 mg kg-1 day-1 oral simvastatin and Group 8: The 10 mg kg-1 i.p. fotemustine (single dose), 3 mg kg-1 day-1 i.p. dexamethasone and 3 mg kg-1 day-1 oral simvastatin. On day 18, animals were euthanized and the brains were removed. The tumors were dissected and weighed. For each of the groups, 5 brains were used for fatty acid analysis and the other 5 were used for immunohistochemical analysis.
Histological and immunohistochemical analysis: Removed brains were fixated in formaldehyde and then kept in a water bath for 1 day. Following the water bath brains were kept in 70, 90 and 96° alcohol for a day in each and for 12 h in 100° alcohol.
Hemotoxylin Eosin staining: Both sections taken out from distilled water were stained with Erlich hemotoxylin for 15-20 min and then rinsed with tap water. Sections were then washed with acid-alcohol (70%, 100 mL+HCl 1 cc) to achieve a lighter color and then with water. Sections were kept in tap water for 15-30 min until the color of the nuclei were dark purple (performed under microscope). Sections then were kept in Eosin solution for 3-5 min (until sections reached a bluish color) washed with distilled water, 70% alcohol and then rinsed twice with 96% alcohol for 1-2 min. Finally, sections were rinsed with absolute alcohol, toluol 1 and 2 and then sealed with balsam.
Cell proliferation: Serial 4 μm paraffin-embedded sections were sequentially deparaffinized and rehydrated and then pre-treated with H2O2 for 10-15 min to quench endogenous peroxidase activity. Following 10 min incubation with ultra V-block solution, sections were immunostained with the specific primary antibody (Ki-67, Rabbit monoclonal). Detection reaction was developed with a secondary biotinylated goat anti-polivalent antibody followed by incubation with the streptavidin peroxidase (Kaur et al., 2004; Sonzogni et al., 2004; Yazici et al., 1994).
Fatty acid analysis: Fatty Acid (FA) profiles were prepared by a slight modification of a method previously described by Yazici et al. (1992). Accurately weighted portion of each sample was homogenized in cold 154 mM NaCl. Total lipids and added internal standard (100 μg non-adecanoic acid in choloroform, Sigma Chemical Co., St Louis) were extracted with chloroform/methanol (2:1) containing 0.005% butylated hydroxytoluene. The chloroform phase was removed and evaporated to dryness under a stream of nitrogen. Total lipids were saponified with 2% KOH in methanol and the FAs methylated with 14% BF3 in methanol. The resulting FA Methyl Esters (FAMEs) were extracted with hexane and analysed by capillary gas chromatography (Perkin-Elmer 8420 Capillary Gas Chromatography, Gouda, The Netherlands). Column: 50x0.25 mm WCOT fused silica, CP-sil 88, Flame-Ionization Detector (FID) temperature 300°C, oven temperature programme from 150-230°C at 2°C min-1, carrier gas N2. The mass spectra of FAME from representative samples were obtained using a Hewlett-Packard (HP) 6890 capillary GC interfaced with a HP mass selective detector and controlled by a HP Chem Station. Column: 25x0.25 mm ID, QC2xBPx70, detector temperature 280°C, oven temperature programme from 100-290°C at 3°C min-1 and carrier gas helium.
The FAMEs were identified by their retention time and compared to those of authentic standards (Sigma Chemical Co., St Louis) and by GC-Mass Spectrometry. The detector response factors were determined by injection equal weights of FAs and internal standard methyl esters on to the column. Their amounts were estimated by calculating the corresponding areas of FA and internal standard.
Statistical analysis: Data were analyzed using Student’s t test and One Way ANOVA, followed by the post hoc Tukey test, p-values ≤0.05 were considered statistically significant.
RESULTS
The number of Ki-67 positive cells decreased by drug therapy compared with control tumors, except dexamethasone alone. The number of Ki-67 positive cells were decreased by 40.3% with fotemustine, 26.5% with simvastatin, 42.25% fotemustine+simvastatin, 19.3% dexamethasone+simvastatin and 51.8% with fotemustine+dexamethasone+simvastatin (p = 0.000-0.020) (Fig. 1 and 2).
Dexamethasone and/or simvastatin decreased tumor size by 30-46% (p = 0.01-0.029). The fotemustine+dexamethasone+simvastatin combination was more effective than the drugs given separately in inhibiting the growth of the tumor (33.9-58.5%, p = 0.000) (Fig. 3).
In the fotemustine+dexamethasone+simvastatin group, the total fatty acid amount was high compared with the control tumors while it was low in the fotemustine+simvastatin group (31.95±1.91 and 16.11±1.96 μg mg-1, respectively, p = 0.021). In the fotemustine+dexamethasone+simvastatin group, C16:0, C18:0, C18:1n-9, C18:1n-7, C24:1n-9 fatty acid amounts were 70-100% higher than the fotemustine, simvastatin, fotemustine+simvastatin and dexamethasone+simvastatin groups (p = 0.003-0.048) (Table 1).
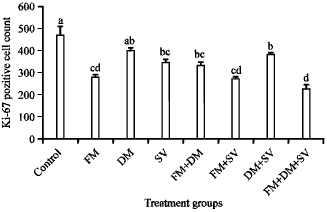 | |
| Fig. 1: | The Ki-67 positive cell count (1/1000). Data is demonstrated as Mean±SEM. The bars which do not contain the same letter are remarkably different from each other (p<0.05). FM: Fotemustine, DM: Dexamethasone, SV: Simvastatin |
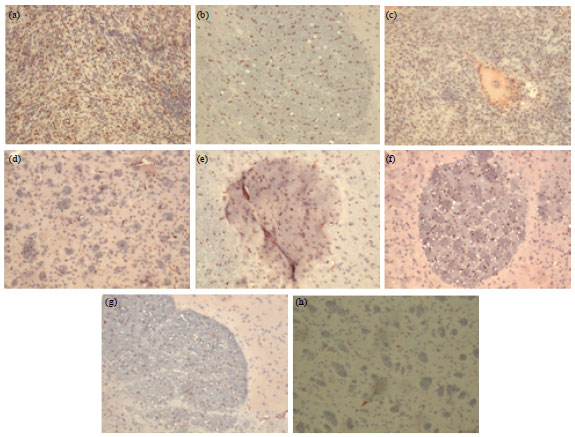 | |
| Fig. 2(a-h): | Tumor tissues immunostained with the Ki-67 antibody. Ki-67 positive cells appear in red (1x10 magnification), (a) Control, (b) Fotemustine, (c) Dexamethasone, (d) Simvastatin, (e) Fotemustine+simvastatin, (f) Fotemustine+dexamethasone, (g) Simvastatin+dexamethasone and (h) Fotemustine+dexamethasone+simvastatin |
| Table 1: | Effect of fotemustine, dexamethasone, simvastatin or their combinations on tumor fatty acid profile |
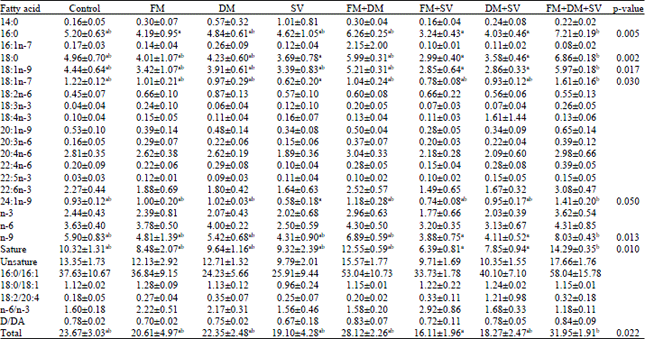 | |
| Data is demonstrated as Mean±SEM. The groups which do not contain the same letter are remarkably different from each other through the column (p<0.05). FM: Fotemustine, DM: Dexamethasone, SV: Simvastatin | |
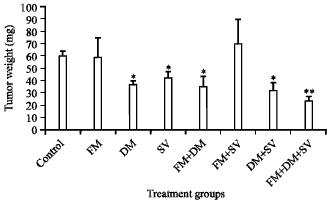 | |
| Fig. 3: | Effect of fotemustine, dexamethasone, simvastatin or their combinations on tumor size. Data is demonstrated as Mean±SEM. *p = 0.029-0.001 vs. control group, **p = 0.000 vs. control group. FM: Fotemustine, DM: Dexamethasone, SV: Simvastatin |
DISCUSSION
In this study, the combined effect of fotemustine with statin and/or dexamethasone on C6 glioblastoma were investigated to find out whether this combination increases the efficacy of fotemustine, a drug from nitrosoureas group which is widely used for the treatment of glioblastoma.
In our preliminary study, C6 glioblastoma cells were inoculated into the left flank area of mice. However, subcutaneous invasion of C6 glioblastoma was found inadequate, therefore, we used intracerebral inoculation model which is more suitable for glioblastoma growth and allows diffusion into the brain parenchyma (Branle et al., 2002).
Mevalonic acid and metabolites play an important role in the growth of tumors. Some tumors such as brain tumor use more LDL and synthesize more cholesterol than normal cells (Maltese, 1983; Rudling et al., 1990). Kikuchi et al. (1997) showed that simvastatin inhibits the growth of human glioma cells, increases the LDL receptors in these cells and that mevalonic acid reverses this effect (Kikuchi et al., 1997). Zhuang et al. (2005) showed that an increase in cholesterol levels promotes tumor growth and reduces apoptosis (Zhuang et al., 2005). Studies showed that statins inhibit cell proliferation and induce apoptosis (Baetta et al., 1997; Frick et al., 2003; Garcia-Roman et al., 2001; Newton et al., 2003). Research based on these studies determined that statins increase the effects of anti-cancer drugs (Denoyelle et al., 2003; Obara et al., 2002; Soma et al., 1995). Our data is in accordance with these findings. Simvastatin significantly inhibited the proliferation of C6 glioblastoma cells and reduced tumor size by 30% compared to the control group. It also insignificantly increased the antiproliferative effect of fotemustine.
It is known that administration of dexamethasone before chemotherapy increases the antitumoral activity of chemotherapeutics (Wang et al., 2004). However, dexamethasone is also used to reduce vasogenic oedema and pain in patients with glioblastoma before chemotherapy which antagonizes the apoptotic effect of the anti-cancer drug and therefore may cause undesired clinical effects (Das et al., 2004). Dexamethasone significantly reducing tumor size in our study as well did not affect the number of Ki-67 positive cells. On the contrary, however insignificantly, it reduced the antiproliferative effects of fotemustine and simvastatin. It has been shown that in in vitro studies, statins prevent the endothelial cells from going in to the G1 phase and inhibit cell proliferation while dexamethasone completely blocks this effect (Newton et al., 2003). It has also been suggested that the reduction in tumor size is associated with the brain oedema due to a brain tumor and that the effect of dexamethasone in reducing the increasing vessel permeability and number in brain tumors has a role in the suppression of the oedema (Heiss et al., 1996; Plate et al., 1993; Wolff et al., 1993). Studies show that dexamethasone prevents inflammation by inhibiting the lymphocyte and microglia infiltration increases the cerebral perfusion, but does not affect the tumor blood flow (Badie et al., 2000; Bastin et al., 2006). A possible mechanism in the dysfunction repairing effect of dexamethasone in the blood-brain barrier is the inhibition of cyclooxygenase-2 (COX-2) enzyme. The pro-tumorigenic effects of COX-2 are largely thought to be attributed to its role in producing PGE2. Prostaglandins which are synthesized in high levels in tumors, especially PGE2, promote tumor growth by inhibiting the cellular immune response (Kokoglu et al., 1998; Portnow et al., 2002).
The fatty acid composition of cancer cells is closely related to carcinogenesis, metastasis and response to chemotherapy (Yazici et al., 1992). Besides being saturated or unsaturated, the series that the polyunsaturated fatty acids belong to is important. Studies show that n-6 series fatty acids stimulate carcinogenesis, tumor growth and metastasis while n-3 series fatty acids have a suppression effect (Corey et al., 1983; Culp et al., 1979; Hanai et al., 1993; Karmali et al., 1984; Mengeaud et al., 1992). It has been determined that the drop in n-6/n-3 fatty acid ratio reduces metastasis potential and this ratio was suggested to play a role in the prevention of cancer and metastasis (Xia et al., 2005). In our study, we determined the fatty acid compositions of tumors in the treatment and control groups and evaluated the 18:2/20:4 ratio which shows the activity of delta-6-desaturase, elongase and delta-5-desaturase; the n-6/n-3 ratio which is closely related to the cancer and immune response and the saturated/unsaturated fatty acid ratios that show the saturation level. There was no significant difference in the n-6/n-3 and saturated/unsaturated fatty acid ratios between groups. The total fatty acid amount was highest in the fotemustine+dexamethasone+simvastatin group and was lowest in the fotemustine+simvastatin group, but there was no significant difference when compared to the control group.
The side effects of fotemustine are dose dependant, progressive and cumulative (Paoletti et al., 1990). In this study, we used 10 mg kg-1 dose of fotemustine which is one forth of Maximum Tolerated Dose (MTD) and still an efficient dose. While single dose fotemustine did not significantly affect the tumor size, it significantly reduced cell proliferation by 40%. The fotemustine+dexamethasone+simvastatin combination we used in order to increase the efficacy of fotemustine, reduced cell proliferation by 52% and tumor size by 58%. The 3 mg kg-1 day-1 dose of simvastatin only reduced cell proliferation by 26% and tumor size by 28%. Kikuchi et al. (1997) found the same dose of simvastatin to be ineffective on intradermal U87MG tumors. However, they showed that simvastatin inhibited tumor growth in combined administration with cytotoxic peroxidated LDL (31). High doses of statins were often used in studies on the antitumoral effect of statins. However, it has been suggested that slow down statin doses tumor proliferation are high, toxic effects may also be higher which might lead to the prohibition of statins being used in cancer treatments. Considering this possibility, we preferred using a dose close to the dose used in hypercholesterolemia (80 mg day-1 for adults).
CONCLUSION
In conclusion, data obtained from this study show that simvastatin increases the cytotoxic effect of fotemustine by inhibiting cell proliferation and that dexamethasone potentializes this cytotoxic effect. The simvastatin dose, we used and found effective was higher than the dose used in hypercholesterolemia treatment but it was 8-16 times lower than the doses used in determining the antitumor activity (Vogelhuber et al., 2002) and it is a tolerable dose by the patients in antitumoral treatment protocols. Normal treatment doses of dexamethasone and fotemustine were used. The first in vivo data were obtained, showing that fotemustine+dexamethasone+simvastatin combination may be useful in brain tumor treatment in this study. Further studies with different doses, time intervals and measures are necessary to strengthen these findings.
REFERENCES
- Altinoz, M.A., E. Ozar, M. Taskin, E. Bozcali and A. Bilir et al., 2001. Vascularization pattern of C6 glioma is modified with medroxyprogesterone acetate and ibuprofen in Wistar rat brain. Pathol. Oncol. Res., 7: 185-189.
CrossRef - Applegate, K.R. and J.A. Glomset, 1986. Computer-based modeling of the conformation and packing properties of docosahexaenoic acid. J. Lipid Res., 27: 658-680.
Direct Link - Badie, B., J.M. Schartner, J. Paul, B.A. Bartley, J. Vorpahl and J.K. Preston, 2000. Dexamethasone-induced abolition of the inflammatory response in an experimental glioma model: A flow cytometry study. J. Neurosurg., 93: 634-639.
Direct Link - Baetta, R., E. Donetti, C. Comparato, M. Calore and A. Rossi et al., 1997. Proapoptotic effect of atorvastatin on stimulated rabbit smooth muscle cells. Pharmacol. Res., 36: 115-121.
CrossRef - Bastin, M.E., T.K. Carpenter, P.A. Armitage, S. Sinha, J.M. Wardlaw and I.R. Whittle, 2006. Effects of dexamethasone on cerebral perfusion and water diffusion in patients with high-grade glioma. Am. J. Neuroradiol., 27: 402-408.
Direct Link - Branle, F., F. Lefranc, I. Camby, J. Jeuken and A. Geurts-Moespot et al., 2002. Evaluation of the efficiency of chemotherapy in in vivo orthotopic models of human glioma cells with and without 1p19q deletions and in C6 rat orthotopic allografts serving for the evaluation of surgery combined with chemotherapy. Cancer, 95: 641-655.
CrossRef - Corey, E.J., C. Shih and J.R. Cashman, 1983. Docosahexaenoic acid is a strong inhibitor of prostaglandin but not leukotriene biosynthesis. Proc. Natl. Acad. Sci. USA., 80: 3581-3584.
PubMedDirect Link - Das, A., N.L. Banik, S.J. Patel and S.K. Ray, 2004. Dexamethasone protected human glioblastoma U87MG cells from temozolomide induced apoptosis by maintaining Bax: Bcl-2 ratio and preventing proteolytic activities. Mol. Cancer, Vol. 3.
CrossRef - De Kruyff, B., P.W.M. Van Dijck, R.W. Goldbach, R.A. Demel and L.L.M. Van Deenen, 1973. Influence of fatty acid and sterol composition on the lipid phase transition and activity of membrane-bound enzymes in Acholeplasma laidlawii. Biochimica Biophysica Acta, 330: 269-282.
CrossRef - Denoyelle, C., P. Albanese, G. Uzan, L. Hong, J.P. Vannier, J. Soria and C. Soria, 2003. Molecular mechanism of the anti-cancer activity of cerivastatin, an inhibitor of HMG-CoA reductase, on aggressive human breast cancer cells. Cell. Signal., 15: 327-338.
CrossRef - Garcia-Roman, N., A.M. Alvarez, M.J. Toro, A. Montes and M.J. Lorenzo, 2001. Lovastatin induces apoptosis of spontaneously immortalized rat brain neuroblasts: Involvement of nonsterol isoprenoid biosynthesis inhibition. Mol. Cell. Neurosci., 17: 329-341.
CrossRef - Hanai, T., T. Hashimoto, K. Nishiwaki, M. Ono and Y. Akamo et al., 1993. Comparison of prostanoids and their precursor fatty acids in human hepatocellular carcinoma and noncancerous reference tissues. J. Surg. Res., 54: 57-60.
CrossRef - Heiss, J.D., E. Papavassiliou, M.J. Merrill, L. Nieman and J.J. Knightly et al., 1996. Mechanism of dexamethasone suppression of brain tumor-associated vascular permeability in rats. Involvement of the glucocorticoid receptor and vascular permeability factor. J. Clin. Invest., 98: 1400-1408.
CrossRefDirect Link - Ip, C., C.A. Carter and M.M. Ip, 1985. Requirement of essential fatty acid for mammary tumorigenesis in the rat. Cancer Res., 45: 1997-2001.
Direct Link - Karmali, R.A., J. Marsh and C. Fuchs, 1984. Effect of omega-3 fatty acids on growth of a rat mammary tumor. J. Natl. Cancer Inst., 73: 457-461.
Direct Link - Khil, M.S., A. Kolozsvary, M. Apple and J.H. Kim, 2000. Increased tumor cures using combined radiosurgery and BCNU in the treatment of 9l glioma in the rat brain. Int. J. Radiat. Oncol. Biol. Phys., 47: 511-516.
CrossRef - Kikuchi, T., Y. Nagata and T. Abe, 1997. In vitro and in vivo antiproliferative effects of simvastatin, an HMG-CoA reductase inhibitor, on human glioma cells. J. Neuro-Oncol., 34: 233-239.
CrossRef - Kokoglu, E., Y. Tuter, K.S. Sandikci, Z. Yazici, E.Z. Ulakoglu, H. Sonmez and E. Ozyurt, 1998. Prostaglandin E2 levels in human brain tumor tissues and arachidonic acid levels in the plasma membrane of human brain tumors. Cancer Lett., 132: 17-21.
PubMedDirect Link - Maltese, W.A., 1983. 3-hydroxy-3-methylglutaryl coenzyme A reductase in human brain tumors. Neurology, 33: 1294-1299.
PubMed - Mengeaud, V., J.L. Nano, S. Fournel and P. Rampal, 1992. Effects of eicosapentaenoic acid, γ-linolenic acid and prostaglandin E1 on three human colon carcinoma cell lines. Prostaglandins Leukotrienes Essent. Fatty Acids, 47: 313-319.
CrossRefPubMedDirect Link - Moore, S.A., E. Yoder, S. Murphy, G.R. Dutton and A.A. Spector, 1991. Astrocytes, not neurons, produce docosahexaenoic acid (22:6 omega-3) and arachidonic acid (20:4 omega-6). J. Neurochem., 56: 518-524.
PubMed - Newton, C.J., Y.X. Xie, C.H. Burgoyne, I. Adams, S.L. Atkin, A. Abidia and P.T. McCollum, 2003. Fluvastatin induces apoptosis of vascular endothelial cells: Blockade by glucocorticoids. Cardiovasc. Surg., 11: 52-60.
CrossRefPubMedDirect Link - Obara, S., M. Nakata, H. Takeshima, J.I. Kuratsu, I. Maruyama and I. Kitajima, 2002. Inhibition of migration of human glioblastoma cells by cerivastatin in association with Focal Adhesion Kinase (FAK). Cancer Lett., 185: 153-161.
CrossRef - Paoletti, P., G. Butti, R. Knerich, P. Gaetani and R. Assietti, 1990. Chemotherapy for malignant gliomas of the brain: A review of ten-years experience. Acta Neurochirurgica, 103: 35-46.
CrossRefDirect Link - Plate, K.H., G. Breier, B. Millauer, A. Ullrich and W. Risau, 1993. Up-regulation of vascular endothelial growth factor and its cognate receptors in a rat glioma model of tumor angiogenesis. Cancer Res., 53: 5822-5827.
Direct Link - Portnow, J., S. Suleman, S.A. Grossman, S. Eller and K. Carson, 2002. A cyclooxygenase-2 (COX-2) inhibitor compared with dexamethasone in a survival study of rats with intracerebral 9L gliosarcomas. Neuro-Oncology, 4: 22-25.
CrossRefDirect Link - Rogers, A.E., 1983. Influence of dietary content of lipids and lipotropic nutrients on chemical carcinogenesis in rats. Cancer Res., 43: 2477-2484.
PubMed - Rudling, M.J., B. Angelin, C.O. Peterson and V.P. Collins, 1990. Low density lipoprotein receptor activity in human intracranial tumors and its relation to the cholesterol requirement. Cancer Res., 50: 483-487.
Direct Link - Scholzen, T. and J. Gerdes, 2000. The Ki-67 protein: From the known and the unknown. J. Cell. Physiol., 182: 311-322.
CrossRefDirect Link - Soma, M.R., R. Baetta, M.R. De Renzis, G. Mazzini and C. Davegna et al., 1995. In vivo enhanced antitumor activity of carmustine [N, N’-bis (2-chloroethyl)- N-nitrosourea] by simvastatin. Cancer Res., 55: 597-602.
Direct Link - Sonzogni, A., G. Colloredo, L. Fabris, M. Cadamuro and B. Paris et al., 2004. Isolated idiopathic bile ductular hyperplasia in patients with persistently abnormal liver function tests. J. Hepatol., 40: 592-598.
CrossRef - Stubbs, C.D. and A.D. Smith, 1984. The modification of mammalian membrane polyunsaturated fatty acid composition in relation to membrane fluidity and function. Biochimica Biophysica Acta, 779: 89-137.
CrossRefPubMedDirect Link - Vogelhuber, W., T. Sprub, G. Bernhardt, A. Buschauer and A. Gopferich, 2002. Efficacy of BCNU and paclitaxel loaded subcutaneous implants in the interstitial chemotherapy of U-87 MG human glioblastoma xenografts. Int. J. Pharm., 238: 111-121.
CrossRef - Wang, H., M. Li, J.J. Rinehart and R. Zhang, 2004. Pretreatment with dexamethasone increases antitumor activity of carboplatin and gemcitabine in mice bearing human cancer xenografts: In vivo activity, pharmacokinetics and clinical implications for cancer chemotherapy. Clin. Cancer Res., 10: 1633-1644.
CrossRefDirect Link - Wolfe, L.S., 1982. Eicosanoids: Prostaglandins, thromboxanes, leukotrienes and other derivatives of carbon‐20 unsaturated fatty acids. J. Neurochem., 38: 1-14.
CrossRefDirect Link - Wolff, J.E., C. Guerin, J. Laterra, J. Bressler, R.R. Indurti, H. Brem and G.W. Goldstein, 1993. Dexamethasone reduces vascular density and plasminogen activator activity in 9L rat brain tumors. Brain Res., 604: 79-85.
CrossRef - Xia, S.H., J. Wang and J.X. Kang, 2005. Decreased n-6/n-3 fatty acid ratio reduces the invasive potential of human lung cancer cells by downregulation of cell adhesion/invasion-related genes. Carcinogens, 26: 779-783.
CrossRef - Yazici, Z., I.A. Tavares, I.F. Stamford, P.M. Bishai and A. Bennett, 1992. Changes in tissue fatty acid composition in murine malignancy and following anticancer therapy. Br. J. Cancer, 65: 163-170.
PubMedDirect Link - Zhuang, L., J. Kim, R.M. Adam, K.R. Solomon and M.R. Freeman, 2005. Cholesterol targeting alters lipid raft composition and cell survival in prostate cancer cells and xenografts. J. Clin. Invest., 115: 959-968.
CrossRefDirect Link