
M. Mohamed Shabi
Department of Pharmacology, Amrita School of Pharmacy, Amrita Institute of Medical Science, Amrita Vishwa Vidyapeetham University, Kochi-682041, Kerala, India
L. Upadhyay
Advance Research Centre for Indian System of Medicine, PRIST University, Thanjavur-613 403, Tamil Nadu, India
International Journal of Pharmacology
Year: 2012 | Volume: 8 | Issue: 1 | Page No.: 46-52







ABSTRACT
The present study was planned to investigate the effect of Desmodium gangeticum (L.) (DG) on lysosomal hydrolases, phosphatases and electrolytes in mechanically induced myocardial ischemic injury in rats. The isolated hearts of rats were subjected to global no-flow ischemia, followed by reperfusion in a Langendorff isolated perfused heart model. The group first hearts (n = 6) served as control where as group second hearts (n = 6) were subjected 30 min of global no-flow ischemia followed by 20 min of reperfusion. The group third and fourth hearts (n = 12) were subjected for preconditioning with DG, at the doses of 50, 100 mg kg-1, before 30 min starting the coronary occlusion followed by 20 min reperfusion. At the end of the study, heart were excised and lysosomal hydrolases, membrane bound phosphatases activities and electrolytes levels were assessed in the cardiac tissues. The activities of different lysosomal enzymes and hydrolases were increased (p<0.05) significantly in the leakage as well as in heart tissue after ischemia-reperfusion injury. Whereas, in preconditioned heart with DG (50, 100 mg kg-1) significantly decreased activities of lysosomal hydrolases in the leakage as well as in the cardiac tissues were observed (p<0.05). Similarly, significant (p<0.05) decreased activity of membrane bound phosphatases such as Na+/K+ ATPase, Ca2+ and Mg2+ ATPase were also observed in the ischemic-reperfusion mediated injured hearts. It can be concluded that alteration in these enzyme activities may lead change in the electrolytes such as sodium, potassium and calcium content in the heart during ischemic reperfusion injury. However, DG preconditioned efficiently reversed the membrane-bound enzymes activity as well as maintained the normal electrolyte concentration.
PDF Abstract XML References Citation
Received: November 20, 2011;
Accepted: January 06, 2012;
Published: February 15, 2012
How to cite this article
M. Mohamed Shabi and L. Upadhyay, 2012. Effect of Myocardial Ischemic Reperfusion Injury on Lysosomal Enzymes and Electrolytes Content of Perfusate and Cardiac Tissue under the Influence of Desmodium gangeticum L.: An In-vitro Study. International Journal of Pharmacology, 8: 46-52.
DOI: 10.3923/ijp.2012.46.52
URL: https://scialert.net/abstract/?doi=ijp.2012.46.52
DOI: 10.3923/ijp.2012.46.52
URL: https://scialert.net/abstract/?doi=ijp.2012.46.52
INTRODUCTION
Myocardial Infarction (MI) is the most common cause of cardiac morbidity and mortality in developing as well as developed countries. Over the last 2 decades, coronary reperfusion therapy has been established for the management of myocardial infarction as an absolute precondition for the survival of ischemic myocardium (De Wood, 1980). Paradoxically, reperfusion of ischemic areas, in particular the re-admission of oxygen, may trigger tissue damage resulting in a spectrum of reperfusion linked pathologies, collectively called as “reperfusion injury” (Braunwald and Kloner, 1985; Moens et al., 2005). Rising evidence from both animal experiments and clinical observations indicates myocardial infarction after ischemia and reperfusion (Liu et al., 2004). Ischemic preconditioning and pharmacological preconditioning are known to protect the heart against reperfusion injury (Ueda et al., 2006).
Myocardial ischemia may directly activates complement pathway results in the cell death and it may releases the intracellular lysosomal hydrolases (Hoffstein et al., 1975). The damage caused by the enzymes of lysosomal and mitochondrial origin as well as the modification of tissue elements by these enzymes play a significant role in myocardial ischemia (Yogeeta et al., 2006). Lysosomal enzymes are major mediators of myocardial infarction and their release into the cytoplasm stimulates the formation of inflammatory mediators (Ravichandran et al., 1991). In addition, lysosomes play a major role in secretion and transport process of different mediators. It has been postulated that the intracellular release of lysosomal enzymes and their subsequent extra-lysosomal activity may exercise a pivotal role in the progressive modifications of reversible myocardial ischemia to irreversible infarction (Decker et al., 1977). In myocardial ischemia, the increased activities of glycohydrolases usually decrease the lysosomal stability resulting in necrosis of myocardium (Sreepriya et al., 1999).
Desmodium gangeticum L. (DG) is an herb belongs to the family Fabaceae widely distributed in tropical and sub-tropical habitats and particularly abundant in India. It is known as salpani in Hindi and shalaparni in Sanskrit. It has been extensively used in Indian system of medicine as a bitter tonic, febrifuge, digestive, anticatarrhal and antiemetic, in inflammatory conditions due to vata disorders (Chopra et al., 1956). This plant has been used in Ayurveda for the treatment of various diseases like typhoid fever, urinary discharges, piles, inflammations, asthma, bronchitis, vomiting, dysentery and hemicranias (Kirtikar and Basu, 1987). The DG is also used in the treatment of ischemic heart disease in the Indian system of medicine (Kirthikar and Basu, 1975). The root of DG is one of the components of Ayurvedic preparations frequently used for the management of different types of heart diseases (Kurian et al., 2005).
Three pterocarpenoids, gangetin, gangetinin and desmodin, are the major phyto-chemical constituents of the roots (Purushothaman et al., 1971, 1975). The alkaloid isolated from aerial part comprises indol-3-alkyl-amines, β-carbolines and anticholinesterase which are well known for their smooth muscle stimulant, CNS stimulant and depressor activities (Ghosal and Bhattacharya, 1972). Gangetin, a pterocarpan, shows anti-fertility activity by affecting alkaline phosphatase activity in uterine fluids (Kawai and Anno, 1971).
Thus keeping above facts in view, the present study was planned to investigate the effect of DG on lysosomal hydrolases, membrane bound phosphatases and electrolytes concentration in mechanically induced myocardial ischemic injury in rats with a view to find out its cardioprotective effect.
MATERIALS AND METHODS
Plant materials: The whole plant of DG was collected from the Kerala state of India in the month of October, 2007. The plant was identified and authenticated taxonomically by Department of environmental and Herbal Science, Tamil University, Tamil Nadu, India. A voucher specimen has been kept in the herbarium of Department of Centre for Advanced Research in Indian System of Medicine (CARISM), SASTRA University, Tamil Nadu, India, for future reference.
Preparation of Petroleum ether extract: The shade-dried roots and parts were powdered and passed through 10-mesh sieve. The coarsely powdered materials were soaked in petroleum ether in the ratio of 1:4 (w/v). The final traces of solvent were removed under reduced pressure and temperature using rotary vacuum evaporator. The yield of extract was calculated as 2.1%. A semi solid extract was obtained after complete elimination of alcohol under reduced pressure. The extract was stored in refrigerator until used.
Chemicals: All chemicals were of analytical grade and chemicals required for sensitive biochemical assays were obtained from Sigma Chemicals, St. Louis, MO, USA. Double distilled water was used in all biochemical assays.
Animal care: Wistar strain male albino rats, weighing 300-350 g were obtained from the Central Animal Facility, SASTRA University, Tamil Nadu, India. The animals were housed individually in polypropylene cages under hygienic and standard environmental conditions (22±2°C, humidity 60-70%, 12 h light/dark cycle). The animals were allowed a standard pellet (Tetragon Chemie Pvt. Ltd., Pet care division, Bangalore, Tamil Nadu, India) and water ad libitum. They were acclimatized to the environment for 10 days prior to experimental use. The study protocol was carried out as per the guidelines of the institutional animal’s ethics committee (IAEC Reg No: 817/04/ac/CPCSEA).
Experimental plan: The experimental plan consisted of four segment:
| • | Stabilization phase: In this phase, hearts were stabilising for 15 min. At the end of a 15 min stabilisation period baseline values were recorded by perfusate |
| • | Intervention phase: Hearts were preconditioned with DG for 10 min |
| • | Ischemia phase: Global no-flow ischemia for 30 min before reperfusion |
| • | Reperfusion phase: Reperfusion period for 20 min after the period of global no-flow ischemia. The pefusate values were recorded by collecting leakage at 0, 1, 2 and 10 min during reperfusion |
The Group 1 (n = 6) hearts served as control and were not provided global no-flow ischemia, DG and reperfusion. Where as Group 2 (n = 6) hearts were subjected 30 min of global no-flow ischemia, followed by 20 min of reperfusion without preconditioned with DG. Group 3 and 4 (n = 12), were preconditioning with DG at the dose of 50, 100 mg kg-1 b.wt. before starting at 30 min before coronary occlusion followed by 20 min reperfusion.
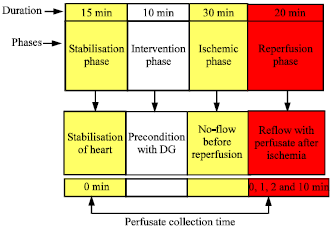 | |
| Fig. 1: | The phases of experimental design |
The perfusate values were recorded by collecting leakage at 0, 1, 2 and 10 min during reperfusion. Figure 1 shows the experimental plan consisted of four segment:
| • | Stabilization phase: Hearts were stabilising for 15 min |
| • | Intervention phase: An intervention period, heart were preconditioned with DG) |
| • | Ischemia phase: Global no-flow ischemia for 30 min before reperfusion |
| • | Reperfusion phase: Reperfusion period for 20 min after the period of ischemia |
Experimental method of global Ischemia-reperfusion in isolated perfused rat heart: The experiments were performed in accordance with the regulations of the local Institutional Animal Ethical Committee regulations. Animals were anesthetized by i.p. administration of sodium pentobarbital (60 mg kg-1). Heparin (1000 IU i.p.) was administered 20 min before sacrificing the animal. The thorax was opened and the heart was excised and rapidly attached to a Langendorff apparatus via the aorta for retrograde perfusion with Krebs-Henseleit buffer solution (Composition of perfusate : mmol L-1: NaCl 119.0, NaHCO3 25.5, KCl 4.3, KH2PO4 1.2, MgSO4 1.2, CaCl2 2.5 and glucose 11.0). The perfusate solution was equilibrated with 95% O2 and 5% CO2, maintained at 37°C and pH 7.4. Perfusion pressure was maintained at 80 cm H2O. The flow rate was maintained at 9-13 mL min-1. The heart was enclosed by a double walled jacket, the temperature of which was maintained by circulating water heated to 37°C. Global ischemia was produced for 30 min, by blocking the in-flow of Krebs Henseleit buffer solution. It was followed by reperfusion for 20 min. Basal coronary leakage was collected after stabilization, before global ischemia and 0, 1, 2 and 10 min leakage also collected after immediately, after reperfusion and heart tissue was excised immediately and rinsed in ice-cold normal saline. Blotted with a filter paper, weighed 10% homogenate was prepared in 10.0 mL of 0.1 M Tris-HCl buffer (pH 7.4) solution. The homogenate was centrifuged at 3, 000 rpm for 15 min and the supernatant was used for the estimation of various biochemical parameters.
Assay of lysosomal hydrolases: The activities of the lysosomal enzymes β-glucuronidase (Kawai and Anno, 1971), β-N-acetyl glucosaminidase (Moore and Morris, 1982) and β-galactosidase (Conchie et al., 1967) were assayed by incubating the sample of perfusate and heart tissue homogenate. The enzyme activity was calculated by estimating the concentration of p-nitrophenol liberated as hydrolytic product. Cathepsin-D activity was assayed by the method of Sapolsky et al. (1973). Acid phosphatase activity was also estimated (King, 1965).
Assay of membrane-bound enzymes: The homogenate was used for the assays of membrane-bound enzymes such as Na+ K+-ATPase (Bonting, 1970), Ca2+-ATPase (Hjerten and Pan, 1983) and Mg2+-ATPase (Ohniski et al., 1982). The activity was expressed by the amount of phosphorus liberated from the incubation mixture containing tissue homogenate, ATP and the respective chloride salt of the electrolytes. The content of protein in the heart was determined by Lowry et al. (1951).
Estimation of heart tissue electrolytes: The levels of Na+, K+ and Ca2+ were estimated in heart homogenate after digestion with nitric acid and perchloric acid as described by Ballentine and Burford (1957).
Statistical analysis: Data are presented as Mean±Standard Deviation (SD). Statistical analysis was performed by one-way Analysis of Variance (ANOVA) followed by the Duncan’s Multiple Range Test (DMRT) with control as the reference. Changes within and between groups were considered statistically significant when the p-value was less than 0.05. All data were analyzed with the aid of statistical package program SPSS 12.0 for Windows.
RESULTS
Table 1-3 show the activity of lysosomal hydrolases such as β-D-glucuronidase, β-D-galactosidase, β-D-N acetylglucosaminidase, acid phosphatase and cathepsin-D in the leakage. The lysosomal hydrolases activity was increased (p<0.05) significantly in the leakage of mechanically challenged ischemia-reperfusion injured hearts (Group 2) against the normal heart (Group 1). However, lysosomal hydrolases in leakage were decreased (p<0.05) significantly in DG (50 and 100 mg kg-1) pre-conditioned hearts (Group 3, 4).
| Table 1: | Effect of Desmodium gangeticum on the activities of β-Glucuronidase and β-Galactosidase enzymes in leakage of normal and ischemia reperfusion injured hearts |
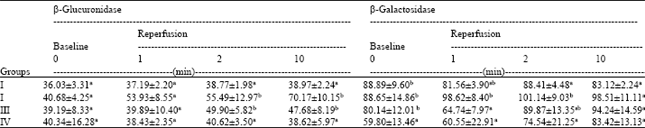 | |
| Activities are expressed as μM of p-nitrophenol liberated min-1 mg-1 of protein for β-glucuronidase and β-galactosidase. Values are expressed as Mean±SD. Values not sharing a common superscript differ significantly at p<0.05, DMRT | |
| Table 2: | Effect of Desmodium gangeticum on the activities of β-N-Acetylglucosaminidase and Cathepsin- D enzymes in leakage of normal and ischemia reperfusion injured hearts |
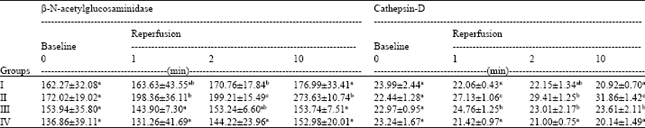 | |
| Activities are expressed as μM of p-nitrophenol liberated min-1 mg-1 of protein for β-N-acetyl glucosaminidase and μmol of tyrosine liberated per h/100 mg protein for cathepsin-D. Values are expressed as Mean±SD. Values not sharing a common superscript differ significantly at p<0.05, DMRT | |
| Table 3: | Effect of Desmodium gangeticum on the activity of Acid phosphatase enzyme in leakage of normal and ischemia reperfusion injured hearts |
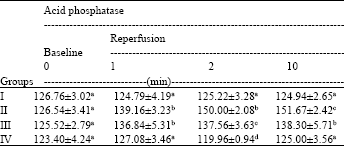 | |
| Activities are expressed as μM of phenol liberated per hour/100 mg of protein for ACP. Values are expressed as Mean±SD. Values not sharing a common superscript differ significantly at p<0.05, DMRT | |
| Table 4: | Effect of Desmodium gangeticum on heart lysosomal hydrolases (β-glucuronidase, β-galactosidase, β-N-acetylglucosaminidase) in normal and ischemia reperfusion injured hearts |
 | |
| Activities are expressed as μM of p-nitrophenol liberated/min/mg of protein for β-glucuronidase, β-galactosidase and β-N-acetylglucosaminidase. Values are expressed as Mean±SD. Values not sharing a common superscript differ significantly at p<0.05, DMRT | |
Table 4 and 5 show the activities of lysosomal hydrolases in heart tissue. The enzymes were significantly increased (p<0.05) in mechanically challenged ischemia- reperfusion injured hearts (Group 2) when compared with the normal group.
| Table 5: | Effect of Desmodium gangeticum on heart lysosomal hydrolases (cathepsin-D and acid phosphatase) in normal and ischemia reperfusion injured hearts |
 | |
| Activities are expressed as μmol of tyrosine liberated per h/100 mg protein for cathepsin-D and μM of phenol liberated per h/100 mg of protein for ACP. Values are expressed as Mean±SD. Values not sharing a common superscript differ significantly at p<0.05, DMRT | |
| Table 6: | Effect of Desmodium gangeticum on cardiac membrane bound enzymes in normal and ischemia reperfusion injured hearts |
 | |
| Activity is expressed as μmol of Pi liberated min-1 mg-1 of protein. Values are expressed as Mean±SD. Values not sharing a common superscript differ significantly at p<0.05, DMRT | |
The activity of lysosomal hydrolases were decreased (p<0.05) significantly in DG (50 and 100 mg kg-1) pre- conditioned hearts (Group 3, 4).
Table 6 shows the activities of the membrane bound phosphatases in the heart tissue. Activities of Na+-K+ ATPase, Mg2+ and Ca2+ ATPase was significantly (p< 0.05) decreased in mechanically challenged ischemia-reperfusion injured hearts (Group 2) when compared to normal heart (Group 1). Pre-conditioned with DG (50 and 100 mg kg-1) blocked the decrease of heart (Group 3, 4) ATPase activities.
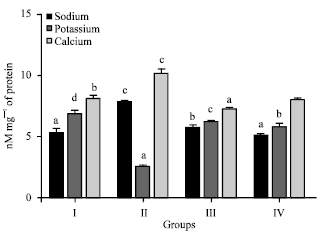 | |
| Fig. 2: | Effect of Desmodium gangeticum on sodium, potassium and calcium in normal and ischemia reperfusion injured hearts. Activity is expressed as μmol mg-1 of protein. Values are expressed as Mean±SD. Values not sharing a common superscript differ significantly at p<0.05, DMRT |
Figure 2 illustrates the electrolyte level in the heart tissues. The level of sodium and calcium were significantly (p<0.05) increased and the level of potassium was significantly (p<0.05) decreased in mechanically challenged ischemia- reperfusion injured hearts (Group 2). Hearts preconditioned with DG (50 and 100 mg kg-1) significantly (p<0.05) decreased the level of sodium and calcium, whereas the level of potassium was increased significantly (p<0.05) (Group 3, 4).
DISCUSSION
We have demonstrated that in this rat model of 30 min of myocardial ischemia and 20 min reperfusion- in vitro by ischemic-preconditioning with DG. After myocardial ischemia, lethal reperfusion injury characterizes a situation in which the myocardium loses viability by mechanisms which are triggered by the restoration of oxygen and substrate supply (Jennings and Yellon, 1992). The macromolecules that leak from damaged tissue enzymes because of their tissue specificity and catalytic activity are the best markers of tissue damage (Hearse, 1979). Lysosomes are a group of cytoplasmic organelles present in numerous animal tissues and are characterized by their acid hydrolases content. Cardiac myocytes effect may also be responsible for the release of lysosomal enzymes. Lysosomal compartment is the major site of intracellular protein degradation. Its interior is acidic and contains numerous hydrolytic enzymes that can degrade nearly all cellular components (Sudharsan et al., 2006). The release of β-D-glucouronidase is frequently used as an index of lysosomal membrane integrity (Ravichandran et al., 1990). In our finding, decrease in the level of lysosomal enzymes after global no-flow and reperfusion might be due to decreased stability of lysosomal membrane resulting in irreversible myocyte damage in ischemia- reperfused hearts (Group 2). Cathepsin is a most powerful hydrolytic enzyme with aspartic protease in action (Padmanabhan and Prince, 2007). It has been reported that localization of acid hydrolases in cardiac myocytes is in the lysosome and the release of these enzyme from the lysosome to the cytosol leads to myocardial cellular injury and death in ischemic state of the heart (Riccutti, 1972; Decker and Wildenthal, 1978). The β-adrenergic stimulation administration result in decreased stability of membranes, which was reflected by the lowered activity of cathepsin-D in lysosomes (Nirmala and Puranakrishnan, 1996). A significant loss of β-D-N-acetylglucosaminidase and β-D-glucuronidase activities in lysosomal fraction of myocardial ischemic animals (Kennett and Weglicki, 1987). In our findings, the activity of cathepsin-D, β-D-N-acetylglucosaminidase and β-D-glucuronidase were significantly decreased in myocardial ischemia- reperfused hearts (Group 2). Hearts preconditioned with DG (50 and 100 mg kg-1) lowered the activities of lysosomal hydrolases in leakage and heart tissue. This activity protects the heart from cellular injury.
ATPases of the cardiac cell play a significant role in the contraction and relaxation cycles of the cardiac muscle by maintaining the normal ion levels inside the myocyte (Mourelle and Franco, 1991). The inhibition of Na+-K+ ATPase can activate the Na+ -Ca2+ exchange mechanism in the myocardium. This Na+-Ca2+ exchange mechanism may play a role in regulating the cellular calcium level (Trump et al., 1984). Numerous factors are known to modify the levels of ATPase, particularly lipid peroxidation and membrane fluidity. Chernysheva et al. (1980) have reported that catecholamine treatment resulted in a decrease in the activities of membrane-bound ATPases. The loss of ATPase activity in the global ischemic state might be responsible for causing not only functional damage but also reversible cell damage changes in the involved cardiac cell. In our investigation, the level of Na+-K+ ATPase, Ca2+ ATPase and Mg-ATPase were significantly decreased in cardiac ischemia reperfused hearts (Group 2). The activity of membrane bound enzyme in DG preconditioned hearts might normalize the intracellular Ca2+ level thereby protecting the myocardium from excess damage by maintaining the membrane integrity. The action of DG (50 and 100 mg kg-1) might be protecting the SH groups of the membrane-bound enzymes from oxidation. In the cell, ATPases are intimately associated with the plasma membrane and participates in the energy dependent transport of sodium, potassium, calcium and magnesium translocation (Mourelle and Franco, 1991). In addition, depletion of ATP leads to the opening of K+ channel leading to the decrease in K+ ions in the myocardial tissue. This may be due to decrease in the heart tissue K+ concentration in ischemia- reperfusion injured hearts. The increase in the myocardial calcium content during the cardiac injury might be due to the enhanced Ca2+ influx (Panagia et al., 1985). Increased level of intracellular Na + concentration activates to depress Ca2+ infux. Heart preconditioned with DG (50 and 100 mg kg-1) decreased the level of sodium and calcium and increased K concentration in ischemia- reperfusion injured hearts.
CONCLUSION
In conclusion, this study reveals that the myocardial necrosis resulted in substantial alterations in the biochemical variables and lysosomal enzymes damage in the heart tissue in mechanically induced ischemia- reperfusion injured hearts. Hearts preconditioned with Desmodium gangeticum (L.) reproduced the levels of lysosomal hydrolases, membrane-bound phosphatases and electrolytes to near normal levels. This normalcy level recognizes Desmodium gangeticum (L.) is the potent cardioprotective agent. In our laboratory, in vitro studies are in progress to determine the mechanism of action of Desmodium gangeticum (L.) by isolated heart preparation.
ACKNOWLEDGMENTS
We thank the Department of Science and Technology, Government of India for providing Senio Research Fellowship and Prof. R. Sethuraman, Vice-chancellor, SASTRA University for his support and encouragement throughout the course of this work
REFERENCES
- Braunwald, E. and R.A. Kloner, 1985. Myocardial reperfusion: A double-edged sword?. J. Clin. Invest., 76: 1713-1719.
CrossRef - Chernysheva, G.V., L.V. Stoida, G.G. Amaranrova and I.L. Kuzmina, 1980. Effect of disseminated necrosis of the myocardium on ATPase activity, Ca++ transport and lipid peroxidation of mitochondrial and microsomal membranes of the heart. Bull. Exp. Biol. Med., 89: 615-618.
CrossRef - Decker, R.S., A.R. Poole, E.E. Griffin, J.T. Dingle and K. Wildenthal, 1977. Altered distribution of lysosomal cathepsin-D in ischemic myocardium. J. Clin. Invest., 59: 911-921.
CrossRef - Decker, R.S. and K. Wildenthal, 1978. Sequential lysosomal alterations during cardiac ischemia. II. Ultrastructural and cytochemical changes. Lab. Invest., 38: 662-673.
PubMed - Kurian, G.A., P. Sachu and T. Varghese, 2005. Effect of aqueous extract of the desmodium gangeticum DC root in the severity of myocardial infarction. J. Ethnopharmacol., 97: 457-461.
PubMed - Hjerten, S. and H. Pan, 1983. Purification and characterization of two forms of a low-affinity Ca2+-ATPase from erythrocyte membranes. Biochimica Biophysica Acta (BBA)-Biomembr., 728: 281-288.
CrossRefDirect Link - Hoffstein, S., D.E. Gennaro, G. Weissmann, J. Hirsch, F. Streuli and A.C. Fox, 1975. Cytochemical localization of lysosomal enzyme activity in normal and ischemic dog myocardium. Am. J. Pathol., 79: 193-206.
Direct Link - Liu, H.R., F. Gao, L. Tao, W.L. Yan and E. Gao et al., 2004. Antiapoptotic mechanisms of benidipine in the ischemic/reperfused heart. Br. J. Pharmacol., 142: 627-634.
PubMed - Kawai, Y. and K. Anno, 1971. Mucopolysaccharide-degrading enzymes from the liver of the squid, Ommastrephes sloani pacificus I. Hyaluronidase. Biochim. Biophys. Acta-(BBA) Enzymol., 242: 428-436.
CrossRefDirect Link - Ueda, K., H. Takano, H. Hasegawa, Y. Niitsuma, Y. Qin, M. Ohtsuka and I. Komuro, 2006. Granulocyte colony stimulating factor directly inhibits myocardial ischemia-reperfusion injury through akt-endothelial NO synthase pathway. Arterioscler. Throm. Vasc. Biol., 26: e108-e113.
Direct Link - Kennett, F.F. and W.B. Weglicki, 1987. Lack of effect of methyl prednisolone on lysosomal and microsomal enzymes after of well defined canine myocardial ischemic. Circ. Res., 43: 759-768.
Direct Link - Moens, AL., M.J. Claeys, J.P. Timmermans and C.J. Vrints, 2005. Myocardial ischemia reperfusion-injury, a clinical view on a complex pathophysiological process. Int. J. Cardiol., 100: 179-190.
Direct Link - Moore, J.C. and J.E. Morris, 1982. A simple automated colorimetric method for determination of N-acetyl-β-D-glucosaminidase. Ann. Clin. Biochem., 19: 157-159.
CrossRefDirect Link - Mourelle, M. and M.T. Franco, 1991. Erythrocyte defects precede the onset of CCl4 induced liver cirrhosis: Protection by silymarin. Life Sci., 48: 1083-1090.
CrossRefPubMedDirect Link - Nirmala, C. and R. Puranakrishnan, 1996. Effect of curcumin on certain lysosomal hydrolases in isoproterenol induced myocardial infarction in rats. Biochem. Pharmacol., 51: 47-51.
PubMed - Ohnishi, T., T. Suzuki, Y. Suzuki and K. Ozawa, 1982. A comparative study of plasma membrane Mg2+-ATPase activities in normal, regenerating and malignant cells. Biochim. Biophys. Acta (BBA)-Biomembr., 684: 67-74.
CrossRefPubMedDirect Link - Padmanabhan, M. and P.S.M. Prince, 2007. S-allylcysteine ameliorates isoproterenol-induced cardiac toxicity in rats by stabilizing cardiac mitochondrial and lysosomal enzymes. Life Sci., 80: 972-978.
PubMed - Purushothaman, K.K., V.M. Kishore, V. Narayanaswami and J.D. Connolly, 1971. The structure and stereochemistry of gangetin, a new pterocarpan from Desmodium gangeticum (Leguminosae). J. Chem. Soc., 1971: 2420-2422.
CrossRefDirect Link - Purushothaman, K.K., S. Chandrasekharan, K. Balakrishna and J.D. Connolly, 1975. Gangetinin and desmodin, two minor pterocarpanoids of Desmodium gangeticum. Phytochemistry, 14: 1129-1130.
Direct Link - Ravichandran, L.V., R. Puvanakrishnan and K.T. Joseph, 1991. Influence of isoproterenol-induced myocardial infarction on certain glycohydrolases and cathepsins in rats. Biochem. Med. Metab. Biol., 45: 6-15.
PubMed - Ravichandran, L.V., R. Puvanakrishnan and T.K. Joseph, 1990. Alterations in heart lysosomal stability in isoproterenol induced myocardial infarction in rats. Biochem. Int., 22: 387-396.
PubMed - Riccutti, M.A., 1972. Myocardial lysosomal stability in the early stages of acute ischemic injury. Am. J. Cardiol., 30: 492-497.
CrossRef - Sapolsky, A.I., R.D. Altman and D.S. Howell, 1973. Cathepsin-D activity in normal and osteoarthritic human cartilage. Fed. Proc., 32: 1489-1493.
PubMedDirect Link - Sudharsan, P.T., Y. Mythili, E. Selvakumar and P. Varalakshmi, 2006. Lupeol and its ester ameliorate the cyclophosphamide provoked cardiac lysosomal damage studied in rat. Mol. Cell. Biochem., 282: 23-29.
Direct Link - Trump, B.F., I.K. Bereezeky, T. Sato, K.V. Liaho, P.C. Phelps and N. Declaris, 1984. Cell calcium, cell injury and cell death. Environ. Health Pers., 57: 281-287.
Direct Link - Yogeeta, S.K., A. Gnanapragasam, S. Senthilkumar, R. Subhashini and T. Devaki, 2006. Synergestic salubrious effect of ferulic acid and ascorbic acid on membrane bound phosphatases and lysosomal hydrolase during experimental myocardial infarction in rats. Life Sci., 80: 258-263.
Direct Link - Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link