
Charles Ansah
Department of Pharmacology, Faculty of Pharmacy and Pharmaceutical Sciences,College of Health Sciences, Kwame Nkrumah University of Science and Technology, Kumasi, Ghana
Evans A.A. Mfoafo
Department of Pharmacology, Faculty of Pharmacy and Pharmaceutical Sciences,College of Health Sciences, Kwame Nkrumah University of Science and Technology, Kumasi, Ghana
Eric Woode
Department of Pharmacology, Faculty of Pharmacy and Pharmaceutical Sciences,College of Health Sciences, Kwame Nkrumah University of Science and Technology, Kumasi, Ghana
Mahama Duwiejua
Department of Pharmacology, Faculty of Pharmacy and Pharmaceutical Sciences,College of Health Sciences, Kwame Nkrumah University of Science and Technology, Kumasi, Ghana
International Journal of Pharmacology
Year: 2008 | Volume: 4 | Issue: 1 | Page No.: 20-26







ABSTRACT
We studied the behavioural effects of the aqueous extract of Cryptolepis sanguinolenta (cryptolepis) in mice based on findings of a sedative action of cryptolepis in the pentobarbitone-induced sleeping-time model and the reported traditional use in the management of insomnia. Cryptolepis (100, 300 and 1000 mg kg-1, p.o) was evaluated in the elevated plus maze, open field and the hole board. We assessed entries and the time spent in the two arms of the elevated plus maze, entries and the time spent in the centre, periphery and corners of the open field and head-dipping behavior in the hole board. Cryptolepis (100, 300 and 1000 mg kg-1) significantly (p<0.05) increased the time spent in the closed arms of the elevated plus-maze, increased the time spent (p<0.001) in the corners of the open field apparatus and decreased head-dipping (p<0.01) behaviour in the hole board. The effects of cryptolepis were similar to that of caffeine used as a reference anxiogenic but completely opposite to that of diazepam, a typical anxiolytic. Present findings indicate that cryptolepis has an anxiogenic-like effect in mice.
PDF Abstract XML References Citation
How to cite this article
Charles Ansah, Evans A.A. Mfoafo, Eric Woode and Mahama Duwiejua, 2008. Anxiogenic Effects of an Aqueous Crude Extract of Cryptolepis sanguinolenta (Periplocaceae) in Mice. International Journal of Pharmacology, 4: 20-26.
DOI: 10.3923/ijp.2008.20.26
URL: https://scialert.net/abstract/?doi=ijp.2008.20.26
DOI: 10.3923/ijp.2008.20.26
URL: https://scialert.net/abstract/?doi=ijp.2008.20.26
INTRODUCTION
About 450 million people are estimated to suffer from some mental or behavioural disorder (WHO, 2001). Among this number only a small fraction receive some level of treatment contributing to the global burden of disease. This number is expected to increase by the year 2020 (Reynolds, 2003). Most of the conventional drugs for the treatment of neurological disorders are associated with unwanted side effects. Research is therefore being conducted to investigate safer and more specific and perhaps lower cost therapies. Natural agents including herbs feature in such research because herbs have been reported to be used in the management of psychiatric disorders with fewer harmful effects (Carlini, 2003).
Cryptolepis sanguinolenta (Asclepiadaceae or Periplocaceae) grows abundantly in the West African sub region. It is used primarily in the treatment of malaria but is also reported to possess a variety of biological effects including antimalarial, (Grellier et al., 1996), antibacterial, (Boakye Yiadom, 1979; Cimanga et al., 1996), antifungal, (Cimanga et al., 1998); antiplasmodial, (Paulo et al., 2000), anti inflammatory, (Bamgbose and Noamesi, 1981); hypoglycaemic activities (Bierer et al., 1998), hypotensive and antipyretic properties (Noamesi and Bamgbose, 1980; Raymond-Hamet, 1937), presynaptic alpha adrenoreceptor blocking action (Noamesi and Bamgbose, 1982) and anti-muscarinic properties (Rauwald et al., 1992). Additionally cryptolepis is reported to be used in traditional medicine for the treatment of insomnia (Mshana et al., 2000), but the underlying mechanism has not been elucidated. We were prompted by an observation of a sedative action of cryptolepis in rats in the pentobarbitone-induced sleeping time (unpublished) to study the behavioural effects of cryptolepis in mice using standard and validated animal models such as the elevated plus-maze (Dawson and Tricklebank, 1995), open field (Asano, 1986) and hole board (Takeda et al., 1998).
MATERIALS AND METHODS
Animals: The animals used in the study, ICR mice (20-30 g), were obtained from the animal house of the Department of Pharmacology, Kwame Nkrumah University of Science and Technology, Kumasi, Ghana in July 2006. They were housed in groups of six in stainless steel cages (34x47x18 cm) with soft wood shavings as bedding, fed with normal commercial pellet diet (GAFCO, Tema, Ghana) and given water ad libitum. The animals were kept and handled under conditions similar to what one of us (MD) operated under a UK Home Office License for five years.
Preparation of the aqueous extract from the roots: Powdered roots of the plant Cryptolepis sanguinolenta were obtained from the Centre for Scientific Research into Plant Medicine (CSRPM), Mampong Akwapim, Ghana, where the plant material is routinely used for the treatment of malaria. The powdered roots (600 g) was boiled for 30 min in 5 L of distilled water, decanted and filtered. The filtrate was transferred into a container and kept. The residue was further extracted successively with (3x2500 mL) quantities of distilled water. The filtrates were pooled together and freeze dried to obtain a freeze-dried sample of the crude extract (Yield = 9.2% w/w), subsequently called cryptolepis, which was reconstituted in distilled water prior to use.
Assessment of anxiogenic activity in the open field test: The open field test is one of the tests used to observe general motor activity, exploratory behavior and measures of anxiety (Asano, 1986; Whimbey and Denenberg, 1967). The open field area was made of plain Plexiglas and consisted of a square arena (45x45x20 cm). The floor had a square sheet of Plexiglas (45x45 cm) with the surface divided into sixteen small squares. The apparatus was illuminated by 40 Watts bulb placed at a height of 100 cm. Mice of both sexes were divided into ten groups of six animals in each group. The groups received diazepam (0.1, 0.3 and 1.0 mg kg-1 i.p.), caffeine (10, 30, 100 mg kg-1 i.p.) and cryptolepis (100, 300 and 1000 mg kg-1 p.o.). The last group received normal saline (10 mL kg-1 p.o.) and served as the vehicle-treated control for the treated groups. Thirty minutes after treatment, animals in both the control and treated groups were placed individually in the center of the open field for five minutes and videotaped with a Sony video camera (model DCR-DVD705E, Sony Corporation, Japan). The behaviour tracker software (Version 1.5) was then used to determine the following behavioural parameters for a period of five minutes (Turner, 1972), the number of entries and time spent in the centre, periphery and corners of the field, rearing and nose poking. Immediately after observation in the open field, animals were placed in the elevated plus maze.
Elevated plus maze-induced anxiety: The Elevated plus Maze (EPM), a widely used test based on the natural aversion of rodents to height and open spaces and which is sensitive to both anxiolytics and anxiogenics has been validated for both rats and mice (Dawson and Tricklebank, 1995; Lister, 1987). The maze which was made of plain Plexiglas and was elevated 60 cm from the floor was adapted from Pellow et al. (1985). It consisted of two open arms (15x5x1 cm) and two enclosed arms (15x5x 5 cm) with an open roof, arranged in such a way that the two open arms are opposite each other. After mice had been treated and gone through the open field, they were placed individually at the center of the maze with their heads facing a closed arm and allowed to explore the maze for five minutes during which period they were videotaped with a Sony video camera (model DCR-DVD705E, Sony Corporation, Japan). The behaviour tracker software (Version 1.5) was then used to determine the following behavioural parameters for 5 min; the number of entries and time spent in the closed and open arms and head-dipping. Immediately after observation in the elevated plus maze, animals were placed into the hole board.
Assessment of anxiogenic activity in mice in the Hole board test: The hole board comprised of a square open field (30x30x30 cm) made of plain Plexiglas. A raised floor insert (2.5 cm above the floor) had four holes (2.5 cm diameter) each situated at five cm from each of the corners and towards the centre. It has gained popularity as a model of anxiety and offers a simple method to measure the response of an animal to an unfamiliar environment (Takeda et al., 1998). After mice had been treated and gone through the open field and the elevated plus maze, they were placed individually at the center of the four hole board for a period of five minutes and videotaped with a Sony video camera (model DCR-DVD705E, Sony Corporation, Japan). The behaviour tracker software (Version 1.5) was then used to determine the number and duration of head dipping for a five minute period.
Analysis of results: The results were presented as mean±SEM. The presence of significant differences among means of the groups was determined by one-way ANOVA using Graph Pad Prism for Windows Version 4.02 (Graph Pad Software, San Diego, CA, USA). Significant differences between pairs of groups were calculated using the Newman-Keuls Multiple Comparison Test with the level of significance set at p<0.05.
RESULTS
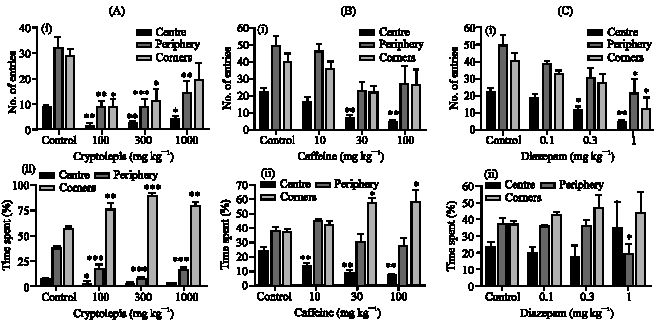
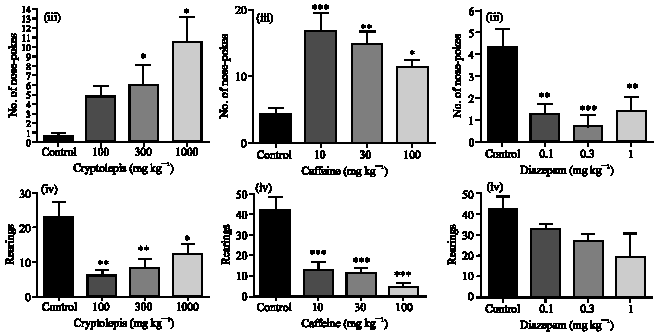
In the test period of 5 min, cryptolepis (100, 300 and 1000 mg kg-1) caused a significant reduction in the number of entries into the corners (p<0.05), centre (p<0.01) and periphery (p<0.01) of the field compared to the vehicle treated control (Fig. 1 ,iA). Whereas the extract caused a decrease in the duration of time spent by mice at the centre and periphery (p<0.001) there was a significant increase in the time spent at the corners (p<0.01) compared to the vehicle treated control (Fig. 1,iiA). There were also significant increases in nose poking (p<0.05) (Fig. 1,iiiA) but reductions in rearing behaviour (p<0.01) (Fig. 1,(ivA) at all doses tested compared to the vehicle treated control. The results obtained for cryptolepis were similar to the typical anxiogenic caffeine (Fig. 1,(i-ivB) but opposite to that of diazepam (Fig. 1,(i-ivC).
| |||
| Fig. 1: | Effect of cryptolepis (100-1000 mg kg-1) (A), caffeine (10-100 mg kg-1) (B) and diazepam (0.1-1.0 mg kg-1) (C) on entries (i), time spent in the periphery, centre and corners (ii), nose poking (iii) and rearing (iv), respectively by mice in the open field. Values are presented as the mean±SEM (n = 6). *: Indicates (p<0.05), **: Indicates (p<0.01) and ***: Indicates (p<0.001) compared to the control group by one-way ANOVA followed by Newman-Keuls Multiple Comparison Test | ||
Cryptolepis (100, 300 and 1000 mg kg-1, p.o) did not significantly (p>0.05) reduce the number of entries and the duration of time spent by mice in the open arm (Table 1). Even though the number of entries into the closed arm was not affected significantly, there was a general increase in the duration of time spent in the closed arm which was significant at the highest dose of 1000 mg kg-1 (p<0.05) (Table 1). The number and duration of head-dips were significantly reduced (both p<0.01) (Table 1) by cryptolepis. The results obtained for cryptolepis were similar to that obtained for the typical anxiogenic caffeine (Table 2) but opposite to that for diazepam (Table 3).
Mice treated with cryptolepis (100, 300 and 1000 mg/kg-1) did show a significant decrease in head dipping behaviour (P < 0.01) and the time spent head dipping (P < 0.01) (Table 4), compared to the vehicle treated control. Caffeine, an anxiogenic produced results similar to cryptolepis with diazepam showing the opposite effect (Table 5).
| Table 1: | Effect of cryptolepis (100-1000 mg kg-1) on the behavioural parameters of mice in the elevated plus maze model of anxiety |
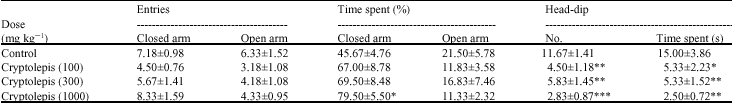 | |
*: Indicates (p<0.05), **: Indicates (p<0.01) and ***: Indicates (p<0.001) compared to the control group by one way ANOVA followed by Newman-Keuls Multiple Comparison Test, Values are presented as the mean±SEM (n = 6) | |
| Table 2: | Effect of caffeine (10-100 mg kg-1) on the behavioural parameters of mice in the elevated plus maze model of anxiety |
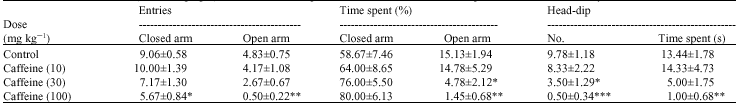 | |
*: Indicates (p<0.05), **: Indicates (p<0.01) and ***: Indicates (p<0.001) compared to the control group by one way ANOVA followed by Newman-Keuls Multiple Comparison Test, Values are presented as the mean±SEM (n = 6) | |
| Table 3: | Effect of diazepam (0.1-1 mg kg-1) on behavioural parameters of mice in the elevated plus maze model of anxiety |
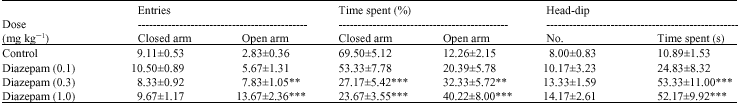 | |
| **: Indicates (p<0.01) and ***: Indicates (p<0.001) compared to the control group by one way ANOVA followed by Newman-Keuls Multiple Comparison Test, Values are presented as the mean±SEM (n = 6) | |
| Table 4: | Effect of cryptolepis on head-dipping behaviour in mice in the hole board |
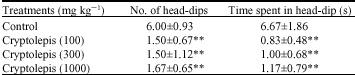 | |
**: Indicates (p<0.01) compared to the control group by one way ANOVA followed by Newman-Keuls Multiple Comparison Test, Values are presented as the mean±SEM (n = 6) | |
| Table 5: | Effect of diazepam and caffeine on head-dipping behaviour in mice in the hole board |
 | |
| *: Indicates (p<0.05) compared to the control group by one way ANOVA followed by Newman-Keuls Multiple Comparison Test, Values are presented as the mean±SEM (n = 6) | |
DISCUSSION
In the present study, the antimalarial herbal, cryptolepis, induced an anxiogenic-like action in ICR mice in all the models used. The EPM represents one of the most widely used animal models for screening anxiogenics (Lister, 1987). The test is based on the apparent natural aversion of rodents to open and high spaces in the measurement of anxiety states in animals. In the test, rodents avoid the exposed open areas of the maze and prefer sections enclosed by protective walls (Weiss et al., 1998). This is because the exposed open areas are presumed to be the most aversive. As a result the animals spend more time in the enclosed arms because they dislike the open arms. This behaviour of rodents on the open arms of the elevated plus maze is considered to be related to anxiety. This anxiety may stem from the novelty, fear and exploration of this aversive environment (Pellow et al., 1985; Briley et al., 1990; Wright et al., 1992; Handley and McBlane, 1993; Rogers and Dalvi, 1997).
Cryptolepis reduced both the number of entries and the time spent by mice in the open arms of the EPM but significantly increased the time spent in the closed arm. Head dipping behavior was significantly reduced by cryptolepis. Whereas anxiolytics reduce the animal`s natural aversion to the open arms and promote exploration, anxiogenic compounds increase the animal`s natural aversion to the open arms and reduce exploration. This makes the animals spend less time in the open arms and spend more time in the closed arm. Therefore, the decreased time spent in the open arms and increased time spent in the closed arm was considered to reflect an anxiogenic effect by cryptolepis in comparison to the control group.
The open-field model also examines anxiety-related behaviour. This behaviour is characterized by the normal aversion of the animal to an open, brightly lit area (Whimbey and Denenberg, 1967; Asano, 1986). Animals are afraid of the aversive centre and as such spend more time in the protective corners. Anxiolytics reduce this natural aversion and promote exploration such that the time spent in the corners is reduced significantly and the time spent in the centre is increased. Anxiogenics increase the animal`s natural aversion and as such increase the time spent by animals in the more protective corners compared to the centre. Cryptolepis treated mice made fewer entries and spent less time in the centre and periphery (the more aversive areas) of the field compared to the corners. However, there was a significant increase in the time spent at the protective corners of the field. There was also a decrease in rearing and an increase in nose poking behaviour in the cryptolepis treated mice compared to the control. Rearing behaviour is widely accepted as an indicator of exploration (Barnett, 1963; Kelley, 1993). Some authorities in this field have suggested that exploration can actually be inhibited by anxiety. Therefore, rearing, an index of exploratory behaviour could represent an indirect measurement of anxiety (Crawley and Goodwin, 1980; Handley and Mithani, 1984; Pellow et al., 1985).
Hence, the observed decrease in rearing by cryptolepis treated mice further supports the anxiogenic-like effect. Rodents tend to display certain specific behavioural patterns such as stretch-attend posture and nose-poking (directed sniffing) when confronted with danger. These behavioural patterns which are categorized as risk assessment behaviour (Blanchard and Blanchard, 1989; Cruz et al., 1994; Rodgers et al., 1997) is the animal`s way of gathering information about the potential threat or danger. It does this by approaching the area cautiously or scanning its surroundings. Risk assessment behaviour is considered a defensive mechanism (Blanchard et al., 1993) and hence an index of anxiety. Therefore the observed increase in nose-poking behaviour in the cryptolepis treated mice is consistent with an anxiogenic-like action. These observations in the open field suggest an increase in anxiety in the cryptolepis treated mice.
The hole-board apparatus has recently become very popular as a model for the measurement of the response of an animal to an unfamiliar environment (Takeda et al., 1998). Studies have revealed that any change in the emotional state of an animal could be depicted by its head-dipping behaviour (Takeda et al., 1998). Since anxiolytics and anxiogenics affect the emotional states of animals they will affect an animal`s head-dipping behaviour. Anxiolytic states are reflected by increases in head-dipping behaviour. This agrees with studies which show that low doses of benzodiazepines (0.3 mg kg-1 diazepam) in mice frequently increase the number and the duration of head-dip in the hole board test (Lister, 1987). Decreases may be an indication of anxiogenicity (Takeda et al., 1998). Caffeine, a typical anxiogenic is reported to decrease the number as well as the duration of head-dipping in the hole board. In this study the significant decreases in head dipping behaviour exhibited by cryptolepis treated mice in the hole board highly suggest an anxiogenic effect for cryptolepis.
Cryptolepis contains several related indoloquinoline alkaloids with the major one being cryptolepine (Dwuma-Badu et al., 1978; Tackie et al., 1993). Cryptolepine is reportedly an alpha-adrenoceptor blocker (Noamesi and Bamgbose, 1980). Certain authorities reported of the anxiolytic-like actions of the α-2 adrenoceptor agonists clonidine and guanabenz in different animal models of anxiety (Fontana et al., 1989; Handley and Mithani, 1984). Though the exact mechanism involved in the anxiolytic-like effect was not elucidated, it may probably be due to the negative feedback inhibition of noradrenaline release mediated by α-2-adrenoceptors present in nerve terminals. Blockade of these receptors could result in a suppression of this inhibition and result in a continual release of noradrenaline. Anxiety and the behavioural effects associated with it in rodents are produced as a result of increase in brain levels of catecholamines. Alpha-2 agonists being implicated in anxiolysis presupposes that an alpha antagonist may show the opposite effect i.e., be anxiogenic-like. This proposition is consistent with the present findings since cryptolepis is reportedly an alpha adrenoceptor blocker (Noamesi and Bamgbose, 1980).
Normally, an agent that prolongs pentobarbitone sleeping time in animals would be expected to be anxiolytic in animal models of anxiety. This has been reported before for some crude extracts of plants (Carvalho-Freitas and Costa, 2002). Cryptolepis reduces spontaneous locomotor activity in mice and prolongs pentobarbitone sleeping time but induced anxiogenic-like behaviour in the present study. Interestingly, this is not the first reported case of a sedative agent inducing anxiogenic-like effect in experimental animals. Indeed, the acetone soluble part of n-hexane extract of Myristica fragrans seeds is reported to be a sedative but possesses anxiogenic-like actions (Ganeshchandra et al., 2001). Furthermore, though buspirone a partial 5HT1A agonist reportedly induced anxiolytic-like behaviour similar to the benzodiazepine-anxiolytics in the modified light-dark model of anxiety (Takashi et al., 1995), other workers have demonstrated its anxiogenic actions in handling-habituated animals (Andrews and File, 1993). These reports suggest that buspirone can be anxiogenic or anxiolytic by acting through different mechanisms. It is plausible that the sedative and anxiogenic-like effects observed with cryptolepis in this study may be by different mechanisms on different receptors. The crude extract of cryptolepis is known to contain several indoloquinoline alkaloids (Paulo et al., 2000), which may be acting by different mechanisms.
Altogether, our study has shown that the aqueous extract of Cryptolepis sanguinolenta is anxiogenic in mice.
ACKNOWLEDGMENT
The authors are grateful to Mr. Thomas Ansah for his technical assistance.
REFERENCES
- Andrews, N. and S.E. File, 1993. Handling history modifies effects of drugs in the elevated-plus maze test of anxiety. Eur. J. Pharmacol., 235: 109-112.
CrossRef - Asano, Y., 1986. Characteristics of open field behavior of Wistar and Sprague Dawley rats. Jikken Dobutsu, 35: 505-508.
PubMedDirect Link - Bamgbose, S.O.A. and B.K. Naomesi, 1981. Studies on cryptolepine II: Inhibition of carrageenan induced oedema by cryptolepine. Planta Med., 41: 392-396.
PubMedDirect Link - Bierer, D.E., D.M. Fort, C.D. Mendez, J. Luo and P.A. Imbacha et al., 1998. Ethnobotanical-directed discovery of the antihyperglycaemic properties of Cryptolepine: Its isolation from Cryptolepis sanguinolenta, synthesis and in vitro and in vivo activities. J. Med. Chem., 41: 894-901.
Direct Link - Blanchard, R.J. and D.C. Blanchard, 1989. Anti-predator defense behaviors in a visible burrow system. J. Comp. Psychol., 103: 70-82.
Direct Link - Blanchard, R.J., E.B. Yudko, R.J. Rodgers and D.C. Blanchard, 1993. Defense system psycho- pharmacology: An ethological approach to the pharmacology of fear and anxiety. Behav. Brain Res., 58: 155-165.
PubMedDirect Link - Boakye-Yiadom, K., 1979. Antimicrobial properties of some West African medicinal plants. II. Antimicrobial activity of aqueous extracts of Cryptolepis sanguinolenta (Lindl.) Schlechter. Q. J. Crude Drug Res., 17: 78-80.
CrossRefDirect Link - Briley, M., P. Chopin and C. Moret, 1990. Effect of serotonergic lesion on anxious behaviour measured in the elevated plus-maze test in the rat. Psychopharmacology, 101: 187-189.
CrossRefDirect Link - Carlini, E.A., 2003. Plants and the central nervous system. Pharmacol. Biochem. Behav., 75: 501-512.
CrossRefPubMedDirect Link - Carvalho-Freitas, M.I.R. and M. Costa, 2002. Anxiolytic and sedative effects of extracts and essential oil from Citrus aurantium L. Biol. Pharma. Bull., 25: 1629-1633.
CrossRefPubMedDirect Link - Cimanga, K., T. De Bruyne and L. Pieters, 1998. Antibacterial and antifungal activities of neocryptolepine, biscryptolepine and cryptoquindoline, alkaloids isolated from Cryptolepis sanguinolenta. Phytomedicine, 5: 209-214.
Direct Link - Crawley, J. and F.K. Goodwin, 1980. Preliminary report of a simple animal behavior model for the anxiolytic effects of benzodiazepines. Pharmacol. Biochem. Behav., 13: 167-170.
PubMedDirect Link - Cruz, A.P.M., F. Frei and F.G. Graeff, 1994. Ethopharmacological analysis of rat behavior on the elevated plus-maze. Pharmacol. Biochem. Behav., 49: 171-176.
PubMedDirect Link - Dawson, G.R. and M.D. Tricklebank, 1995. Use of the elevated plus maze in the search for novel anxiolytic agents. Trends Pharmacol. Sci., 16: 33-36.
CrossRef - Dwuma-Badu, D., J.S. Ayim, N.I. Fiagbe, J.E. Knapp, P.L. Jr, Schiff and D.J. Slatkin, 1978. Constituents of West African medicinal plants XX: Quindoline from Cryptolepis sanguinolenta. J. Pharm. Sci., 67: 433-444.
Direct Link - Fontana, D.J., D.M. Schefke and R.L. Commissaris, 1989. Acute versus chronic clonidine treatment effects on conflict behaviour in the rat. Behav. Pharmacol., 1: 201-208.
Direct Link - Ganeshchandra, S., S. Vikram, K. Veena and B.K. Sanjay, 2001. Behavioral actions of Myristica fragrans seeds. Indian J. Pharmacol., 33: 417-424.
Direct Link - Grellier, P., L. Ramiaramanana, V. Milleriox, E. Deharo and J. Shrevel et al., 1996. Antimalarial activity of cryptolepine and isocryptolepine, alkaloids isolated from Cryptolepis sanguinolenta. Phytother. Res., 10: 317-321.
CrossRefDirect Link - Handley, S.L. and S. Mithani, 1984. Effects of alpha-adrenoceptor agonists and antagonists in a maze exploration model for fear-motivated behavior. Naunyn Schmiedebergs Arch. Pharmacol., 327: 1-5.
CrossRefDirect Link - Handley, S.L. and J.W. McBlane, 1993. 5-HT drugs in animal models of anxiety. Psychopharmacology, 112: 13-20.
CrossRefDirect Link - Lister, R.G., 1987. The use of a plus-maze to measure anxiety in the mouse. Psychopharmacology, 92: 180-185.
CrossRefDirect Link - Noamesi, B.K. and S.O.A. Bamgbose, 1980. The α-adrenoceptor blocking properties of cryptolepine on the rat isolated vas deferens: Studies on cryptolepine. Part I. Planta Med., 39: 51-56.
PubMedDirect Link - Noamesi, B.K. and S.O.A. Bamgbose, 1982. Preferential blockade of pre-synaptic α-adrenoceptors on the rat isolated vas deferens: Studies on cryptolepine. Planta Med., 44: 241-245.
PubMedDirect Link - Paulo, A., E.T. Gomes, J. Steele, D.C. Warhurst and P.J. Houghton, 2000. Antiplasmodial activity of Cryptolepis sanguinolenta alkaloids from leaves and roots. Planta Med., 66: 30-34.
Direct Link - Pellow, S., P. Chopin, S.E. File and M. Briley, 1985. Validation of open: Closed arm entries in an elevated plus-maze as a measure of anxiety in the rat. J. Neurosci. Methods, 14: 149-167.
CrossRefDirect Link - Rauwald, H.W., M. Kober, E. Mutschler and G. Lambrecht, 1992. Cryptolepis sanguinolenta: Antimuscarinic properties of cryptolepine and the alkaloid fraction at M1, M2 and M3 receptors. Planta Med., 58: 486-488.
CrossRefPubMedDirect Link - Reynolds, E.H., 2003. Brain and mind: A challenge for WHO. Lancet, 361: 1924-1925.
CrossRefDirect Link - Rodgers, R.J. and A. Dalvi, 1997. Anxiety, defence and the elevated plus-maze. Neurosci. Biobehav. Rev., 21: 801-810.
CrossRefDirect Link - Rodgers, R.J., B.J. Cao, A. Dalvi and A. Holmes, 1997. Animal models of anxiety: An ethological perspective. Braz. J. Med. Biol. Res., 30: 289-304.
CrossRefDirect Link - Tackie, A.N., G.L. Boye, M.H.M. Sharaf and P.L. Schiff, 1993. Cryptospirolepine, an unique spiro-nonacyclic alkaloid isolated from Cryptolepis sanguinolenta. J. Nat. Prod., 56: 653-670.
Direct Link - Takashi, S., M. Kinzo, O. Miyuki, M. Harumi, T. Katsutoshi and W. Hiroshi, 1995. The modified light/dark transition test in mice: Evaluation of classic and putative anxiolytic and anxiogenic drugs. Genet. Pharmacol., 26: 205-210.
CrossRef - Takeda, H., M. Tsuji and T. Matsumiya, 1998. Changes in head-dipping behavior in the hole-board test reflect the anxiogenic and/or anxiolytic state in mice. Eur. J. Pharmacol., 350: 21-29.
CrossRefDirect Link - Weiss, S.M., G. Wadsworth, A. Fletcher and C.T. Dourish, 1998. Utility of ethological analysis to overcome locomotor confounds in the elevated maze models of anxiety. Neurosci. Biobehav. Rev., 23: 265-271.
CrossRef - Whimbey, A.E. and V.H. Denenberg, 1967. Two independent behavioral dimensions in open field performance. J. Comput. Physiol. Psychol., 63: 500-504.
Direct Link - Wright, I.K., M. Heaton, N. Upton and C.A. Marsden, 1992. Comparison of acute and chronic treatment of various serotonergic agents with those of diazepam and Idazoxan in the elevated plus maze. Psychopharmacology, 107: 405-414.
CrossRefDirect Link