
C. Gnoula
Laboratory of Pharmacognosy, Bromatology and Human Nutrition, Free University of Brussels, Bd du Triomphe, CP 205/9, 1050 Brussels, Belgium
P. Guissou
Health Sciences Training and Research Unit, University of Ouagadougou
P. Duez
Laboratory of Pharmacognosy, Bromatology and Human Nutrition, Free University of Brussels, Bd du Triomphe, CP 205/9, 1050 Brussels, Belgium
M. Frederich
Pharmacognosy Laboratory, University of Liege, CHU du Sart Tilman B36: 4000 Li�ge: Belgium
J. Dubois
Laboratory of Bioanalytical Chemistry, Toxicology and Applied Physical Chemistry,
Free University of Brussels, Bd du Triomphe, CP 205/1, 1050 Brussels, Belgium
International Journal of Pharmacology
Year: 2007 | Volume: 3 | Issue: 3 | Page No.: 280-284







ABSTRACT
The research aims are to characterize this anthelmintic activity and to isolate the main nematocidal agent of Balanties aegyptiaca plant. The anthelmintic activity was evaluated in vitro by means of an original anthelmintic assay using Caenorhabditis elegans as a biological model. Fluorescence microscopy was used for the determination of the percentage of worms death. The structure elucidation was based on NMR, mass spectroscopic analysis and chemical methods. A bioassay-directed fractionation of the aqueous extract of Balanites aegyptiaca led to the isolation of balanitin-7 (also named diosgenin 3β-O-β-D-xylopyranosyl-(1→3)-β-D-glucopyranosyl-(1→4)-[α-L-rhamnopyranosyl-(1→2)]-β-D-glucopyranoside), as being the principal nematicidal agent. These data indicate that balanitin-7 has an appreciable nematocidal activity, which is not mediated by inducing an anti-acetylcholinesterase activity.
PDF Abstract XML References
How to cite this article
C. Gnoula, P. Guissou, P. Duez, M. Frederich and J. Dubois, 2007. Nematocidal Compounds from the Seeds of Balanites aegyptiaca Isolation and Structure Elucidation. International Journal of Pharmacology, 3: 280-284.
DOI: 10.3923/ijp.2007.280.284
URL: https://scialert.net/abstract/?doi=ijp.2007.280.284
DOI: 10.3923/ijp.2007.280.284
URL: https://scialert.net/abstract/?doi=ijp.2007.280.284
INTRODUCTION
The prevalence of nematode intestinal infections is apparently very high and on a global scale, these infections cause severe health problems in man and domestic animals, especially in developing countries. More than 1 billion people are infected with Ascaris lumbricoïdes and hundreds of millions are infected with hookworms and trichuris (Guyatt and Evans, 1992). These infections cause intestinal disorders, discomfort and loss of productivity through direct or indirect interference with host nutrition and metabolism. A number of control measures to combat these infections are available and several classes of synthetic anthelmintic have been shown to be very effective when used strategically in the right epidemiological context.
Unfortunately, in many developing countries people cannot afford anthelmintic drugs (Guyatt and Evans, 1992) and still have to resort to traditional medicine for primary health cares. Indeed, for centuries populations inhabiting the Sahel have used indigenous wild plant to satisfy their requirements for health.
Balanites aegyptiaca (L.) Delile (Zygophyllaceae) is a tropical plant largely used in Africa. All the parts of the plant are used as a component of several medicinal remedies. The bark is employed like purgative and analgesic in the treatment of the colics while the barks of roots are employed for the treatment of psychoses and epilepsy. The fruits are generally employed to purge, to remove intestinal parasites and to treat Schistosomum japonicum (Koko et al., 2000) the bark aqueous extract is traditionally used as anti-jaundice, whereas the mesocarps of the fruit is administered as oral hypoglycaemic (Kamel, 1991) and seems effective against Fasciola gigantica (Koko et al., 2000).
Phytochemical investigations on Balanites aegyptiaca yielded several classes of secondary metabolites, such as coumarins, flavonoïds and steroidal saponins, many of which express biological activities (Sarker et al., 2000). From roots and bark, several steroidal saponins, including yamogenin glycosides were isolated (Liu and Nakanishi, 1982; Pettit et al., 1991; Speroni et al., 2005). From the fruits were obtained 2 furostanol glycosides and 6-methyl disogenin (Hosny et al., 1992; Kamel, 1998).
Using the free living nematode Caenorhabditis elegans as model system for the development of nematocides and anthelmintics (Gnoula et al., 2006), the present study proposes to investigate the in vitro anthelmintic activity of the seeds of Balanites aegyptiaca followed by a bioassay-guided fractionation, isolation and structure elucidation of the nematocidal compound(s).
MATERIALS AND METHODS
Plant material and isolation of the compounds: The fruits of Balanites aegyptiaca were collected in Burkina Faso near Ouagadougou and a voucher specimen was deposited in the BRLU herbarium, Belgium. This study has been conducted in Belgium (Free University of Brussels, ULB) from October 2004 to April 2006.
Powdered seeds (1.5 kg) were defatted with petroleum ether, dried and successively exhausted using chloroform, dichloromethane, ethyl acetate, methanol and water.
Three grams of the dry aqueous extract were chromatographed on silica gel 60 column (300 g, particle size: 0.063-0.200 mm) and eluted successively with chloroform, chloroform/methanol mixtures in increasing proportions from 90:10 to 10:90 and methanol. Fractions were combined upon TLC analysis (spraying with anisaldehyde reagent and heating at 105° for 15 min) to yield 10 fractions which were screened for anthelmintic activity. The most active fraction (F4) was further purified by Centrifugal Partition Chromatography (CPC) on a Kromaton Hydrostatic-Proto equipped with a 200 mL rotor (Angers, France), using chloroform/methanol/butanol/ water (35:30:15:20, v/v) as a solvent system, the supernatant being used as stationary phase and the heaviest phase as mobile phase. 1.4 g of the fraction was dissolved in 10 mL of the stationary phase and submitted to chromatography (900 rpm, 4.5 mL min-1, fractions collected every 2 min, descending mode for 80 min and ascending mode for 40 min) to yield 60 sub fractions including 1 pure compound.
Structure determination: 1H and 13C spectra NMR were measured on a Bruker Avance 300 at 300 and 75 MHZ, respectively, with TMS as an internal standard; 2-D NMR spectra including COSY, HMQC and HMBC were recorded in DMSO on a Varian Unity 600 at 25°C. HRESI-MS were performed on a Micromass QTOF II Mass Spectrometer at a capillary and cone voltage of 2.8 and 80 kV, respectively and a mass resolution of approximately 10,000.
Sugars were identified by HPTLC after hydrolysis with Trifluoroacetic acid 1 M.
Anthelmintic assay: Caenorhabditis elegans wild-type strain and Escherichia coli OP50 strain were generous gifts from Devgen (Gent, Belgium). The worms were grown on Escherichia coli layers in Petri dishes maintained in a thermostated oven at 20°C; worms for the test were larva L3 obtained after a synchronous culture as previously described by Hope (1999). For the nematocidal assays, 5 mL of a worms suspension (approximately 6250 to 75000 worms) in PBS (Phosphate Buffer Solution) were added with the tested drug and maintained at 20°C for up to 7 days. At each time point, a 500 μL aliquot of this suspension was labeled by adding 200 μL of a CFDA (5(6)-Carboxyfluorescein diacetate) working solution (dilution 1/25 in PBS of a stock solution of 3.0 mg CFDA in 1 mL acetone) and leaving for 30 min at room temperature in the dark. After centrifugation (1300 rpm, 2 min) and washing with 5 mL PBS, the pellet was resuspended in 1 mL PBS and divided into 200 μL aliquots containing around 125-150 worms. The proportion of fluorescence-labeled dead worms was then measured by visual counting (triplicate counting) in fluorescence microscopy.
Statistics were computed by Excel 2000 equipped with the add-on Analyse-It (Microsoft), the level of significance being classically set at 0.05.
Acetylcholinesterase assay: The acetylcholinesterase inhibition was measured by a method based on the enzymatic hydrolysis of acetylthiocholine in a thiol that was condensed with 5, 5’-dithiobis [2-nitrobenzoic acid] to yield a yellow anion (Ingkaninan et al., 2003). Galanthamine was used as a positive inhibitor (in our hands, IC50 was 1.44 μg mL-1).
RESULTS
Nematocidal activity: Table 1 presents the data obtained with different Balanites aegyptiaca extracts; whereas marginal activity is observed with the chloroform, dichloromethane, ethyl acetate and methanol extracts, the aqueous extract was found quite active and the dose 1 mg mL-1 killed 50 and 75% worms in 5 and 7 days, respectively (Table 1 and 3).
| Table 1: | Effect of different extracts from Balanites aegyptiaca against Caenorhabaditis elegans |
 | |
| Table 2: | Bio-guided fraction of the aqueous extracts from Balanites aegyptiaca |
 | |
| Table 3: | Toxicity of the aqueous extracts of Balanites aegyptiaca and Balanitin-7 against C. elegans’ larva in 3rd stage or in adult stage (5 doses tested; CFDA fluorescence labelling and counting every 24 h) |
 | |
| Test in triplicate (125 to 150 worns evaluated per sampling points) | |
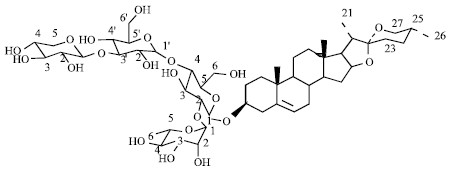 | |
| Fig. 1: | Chemical structure of Balanitin-7 |
Isolation of active compound and structure determination: The C. elegans assay allowed us to easily screen the fractions obtained from the column chromatography of the aqueous extract and to concentrate on the most active fraction (Table 2). CPC yielded 60 sub-fractions.
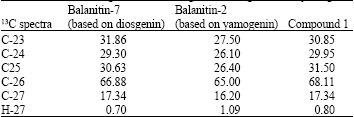
A pure compound, 1 (11.2 mg, 0.8%), positive with anisaldehyde reagent could be isolated by CPC and characterized by NMR spectroscopy. Comparison of 13C spectra with published data (Liu and Nakanishi, 1982; Pettit et al., 1991) for Balanitin- 2 (based on yamogenin) and Balanitin-7 (based on diosgenin) confirmed structures of steroidal saponins. The distinction between the 2 genins relies on the chemical shifts of C-23, C-24, C-25, C-26, C-27 and H-27 (Table 4) and detailed HMBC analysis allowed unambiguous assignation of methyl 27, confirming the diosgenin for compound 1. Sugars were identified as xylose, rhamnose and glucose by TLC. Complete assignment of C and H (Liu and Nakanishi, 1982; Pettit et al., 1991) allowed identifying compound 1 as Balanitin-7 (Fig. 1). Present data fully agree with the literature, except for the 13C signal of xylose 5 located at 67 ppm instead of the published 62 ppm (Liu and Nakanishi, 1982; Pettit et al., 1991) as the same authors assign the 13C signal of xylose 5 to 67 ppm in Balanitin-2, which, given the identical position and configuration of this xylose, should not be different from Balanitin-7, we believe that published data should be revised for this carbon.
| Table 4: | 13C NMR assignment for distinguished diosgenin from yamogenin |
 | |
The exact mass of 1016.5017 obtained by mass spectrometry confirms this assumption.
Balanitin-7 accounts for most of the activity of the extract (1 mg of extract corresponds to approximately 2.3 μg of Balanitin-7); at a concentration of 52 μg mL-1, it killed 75% of larval L3 stage worms in 7 days (Table 3); at 8.3 μg mL-1, it killed about 100% of adult stage worms in 7 h (Table 3). The activity is then strongly stage-dependent.
Acetylcholinesterase inhibition: None of the tested extracts and fractions was inhibitory to acetylcholinesterase; the mechanism of action is then different from the anthelmintics pyrantel and piperazine.
DISCUSSION
The discovery of new anthelmintics relies to a large extent on the use of an effective screening assay to detect activity. Although the best test for an anthelmintic uses the target species in its normal host, this requires relatively large quantities of chemicals and animals breeding facilities, which can be quite expensive. Several nematodes, among which Caenorhabditis elegans, a free-living soil nematode, susceptible to all commercially available anthelmintics (Simpkin and Coles, 1981; Burglin et al., 1998), have been proposed for the development of in vitro drug screening assays (O'Grady and Kotze, 2004). As these assays are quite tedious to perform and rely on the visual appreciation of motility, a new bioscreening test was recently introduced and biologically and analytically validated (Gnoula et al., 2006). Treatment of a suspension of worms with 5(6)-Carboxyfluorescein Diacetate (CFDA) transiently induces fluorescence in dead worms only, allowing a fast and efficient determination of the proportion of dead worms by fluorescence microscopy. The transient aspect of labelling allows for kinetic studies to precise or infirm possible modes of action.
Balanites aegyptiaca is known for a wide range of biological activities which were mainly attributed to its saponin constituents (Sparg et al., 2004).
The present research elaborates on a preliminary work on the efficacy of the aqueous extract of Balanites aegyptiaca; a dose of 2.5 mg mL-1 was reported to cause the death of C. elegans adult worms (45 and 100% after 2 and 6 h contact, respectively; data base on motility assay) (Ibrahim, 1992). The efficiency of the CFDA test allowed us not only to confirm this activity but also to isolate a steroidal saponin with a high potential of anthelmintic activity, Balanitin-7. Comparing to well-known anthelminitics previously tested (Gnoula et al., 2006), Balanitin-7 is more potent than pyrantel and piperazine but less active than mebendazole, thiabendazole and levamisole. On L3 larval stage, the activity of levamisole was marked after only one day of contact with the drug (Gnoula et al., 2006) whereas Balanitin-7 needs a minimum of 3 days. This difference of kinetic, combined with a non-inhibition of acetylcholinesterase, indicates a mechanism of action of Balanitin-7 different from those of levamisole, pyrantel and piperazine.
Further investigations are being carried out to precise the mechanism of action of this promising anthelmintic drug as well as its selectivity index. Given the high activity of the saponin and the common use of Balanites aegyptiaca seeds as an anthelmintic in many countries, it would be extremely interesting to standardize the crude drug with regard to Balanitin-7 content.
ACKNOWLEDGMENTS
This research is funded by the Université Libre de Bruxelles. The authors gratefully acknowledge Pr. M. Devleeschouwer (ULB, Institute of Pharmacy) for his help, Mrs. M. Faes and T. Lamkami for their technical assistance.
REFERENCES
- Burglin, T.R., E. Lobos and M.L. Baxter, 1998. Caenorhabditis elegans as a model for parasitic nematodes. Int. J. Parasitol., 28: 395-411.
PubMedDirect Link - Gnoula, C., P. Guissou, J. Dubois and P. Duez, 2007. 5(6)-Carboxyfluorescein diacetate as an indicator of Caenorhabditis elegans viability for the development of an in vitro anthelmintic drug assay. Talanta, 71: 1886-1892.
CrossRefDirect Link - Ingkaninan, K., P. Temkitthawon, K. Chuenchom, T. Yuyaem and W. Thongnoi, 2003. Screening for acetylcholinesterase inhibitory activity in plants used in Thai traditional rejuvenating and neurotonic remedies. J. Ethnopharmacol., 89: 261-264.
CrossRefDirect Link - Kamel, M.S., 1998. A furostanol saponin from fruits of Balanites aegyptiaca. Phytochemistry, 48: 755-757.
CrossRefDirect Link - Koko, W.S., M. Galal and H.S. Khalil, 2000. Fasciolicidal efficacy of Albizia anthelmintica and Balanites aegyptiaca compared with albendazole. J. Ethnopharmacol., 71: 247-252.
CrossRefDirect Link - O'Grady, J. and A.C. Kotze, 2004. Haemonchus contortus: In vitro drug screening assays with the adult life stage. Exp. Parasitol., 106: 164-172.
CrossRefDirect Link - Sarker, S.D., B. Bartholomew and R.J. Nash, 2000. Alkaloids from Balanites aegyptiaca. Fitoterapia, 71: 328-330.
CrossRefDirect Link - Sparg, S.G., M.E. Light and J. van Staden, 2004. Biological activities and distribution of plant saponins. J. Ethnopharmacol., 94: 219-243.
CrossRefDirect Link - Speroni, E., R. Cervellati, G. Innocenti, S. Costa, M.C. Guerra, S. Dall' Acqua and P. Govoni, 2005. Anti-inflammatory, anti-nociceptive and antioxidant activities of Balanites aegyptiaca (L.) Delile. J. Ethnopharmacol., 98: 117-125.
CrossRefDirect Link