
Murugan Sengottuvelan
Department of Biochemistry and Biotechnology, Faculty of Science,
Annamalai University, Annamalainagar-608 002, Tamilnadu, India
Namasivayam Nalini
Department of Biochemistry and Biotechnology, Faculty of Science,
Annamalai University, Annamalainagar-608 002, Tamilnadu, India
International Journal of Pharmacology
Year: 2006 | Volume: 2 | Issue: 3 | Page No.: 335-340







ABSTRACT
The effect of resveratrol supplementation on the carcinogenic response, hepatic lipid peroxidation and antioxidant levels/activities of rats administered with 1,2-dimethylhydrazine (DMH) (20 mg kg-1 body weight) was studied in male albino Wistar rats. DMH-induced oxidative stress (increased lipid peroxidation and decreased antioxidant defense) was modulated by oral administration of resveratrol (8 mg kg-1 body weight). Liver lipid peroxidation markers CD, LOOHs and TBARS were increased, in contrast hepatic antioxidants enzymes such as superoxide dismutase, catalase, glutathione peroxidase, glutathione S-transferases, glutathione reductase, reduced glutathione, Vitamin C and E activities/levels were decreased on DMH administration. Resveratrol supplementation in three dietary regimens (initiation, post-initiation and entire period) attenuated the liver lipid peroxidation with concomitant up-regulation of enzymic and nonenzymic antioxidant reserves. The reversal of oxidative stress was more pronounced in the entire period resveratrol supplementation. In conclusion resveratrol, a natural product is a suitable candidate as a chemopreventive agent in suppressing the adverse effect of DMH-induced oxidative stress.
PDF Abstract XML References Citation
How to cite this article
Murugan Sengottuvelan and Namasivayam Nalini, 2006. Resveratrol, a Phytoalexin Enhances Hepatic Antioxidant Defense in 1,2-dimethylhydrazine-induced Colon Carcinogenesis. International Journal of Pharmacology, 2: 335-340.
DOI: 10.3923/ijp.2006.335.340
URL: https://scialert.net/abstract/?doi=ijp.2006.335.340
DOI: 10.3923/ijp.2006.335.340
URL: https://scialert.net/abstract/?doi=ijp.2006.335.340
INTRODUCTION
Dimethylhydrazine hydrochloride (DMH) is a potent necrogenic heptocarcinogen that alkylates hepatocellular DNA leading to carcinogenesis (Ying et al., 1979). DMH, an aliphatic methylating carcinogen, is metabolized rapidly by the liver and it induces zonal necrosis and oxidative stress (Hayes et al., 1987). Necrosis can promote hepatocarcinogensis by enhancing growth of initiated hepatocytes resistant to toxicity (Farber and Sarma, 1987). DMH administration has been shown to induce hepatocellular carcinoma and angiosarcoma in mice (St Clair et al., 200l). DMH is also a powerful colon carcinogen (Ohno et al., 2001), which induces colorectal tumors in experimental animals (Weed et al., 1985).
Free radicals and Reactive Oxygen Species (ROS) are involved in the pathogenesis of large number of diseases including cancer is well documented (Feig et al., 1994). The metabolic activation of DMH occurs in the liver (LaMont and O'gorman, 1978) with the formation of active intermediates such as azoxymethane and methylazoxymethanol, are subsequently decomposed to methyldiazonium, an alkylating agent, which generates a reactive carbonium ion capable of methylating DNA, RNA or protein (Matsumoto and Higa, 1966) Moreover, previous reports have pointed out the tendency of DMH to produce free radicals in blood, liver and large bowel of experimental models (Arutiunian et al., 1997).
Lipid peroxidation in vivo is a degenerative, free radical mediated process that primarily involves polyunsaturated fatty acids (PUFA) of biomembranes (Mead, 1976). Auto oxidation of PUFA is known to accompanied by the formation of a complex mixture of products, including aldehydes such as Malondialdehyde (MDA) and other related carbonyl compounds (Esterbauer, 1982). Measurement of primary [conjugated dienes (CD) and lipid hydroperoxides (LOOH)] and secondary products (MDA) is a very sensitive way to determine the effect of antioxidants on in vivo lipid peroxidation (Cheeseman, 1993). An elevation in the hepatic lipid peroxidation with a concomitant decrease in antioxidant defense systems (enzymic-superoxide dismutase, SOD; catalase, CAT; glutathione peroxidase, GPx; glutathione S-transferase, GST and glutathione reductase, GR; non-enzymic antioxidants-reduced glutathione, GSH; Vitamin C and E) intensify peroxidation in blood and large bowel after carcinogen exposure which may influence free radical mediated neoplastic growth (Arutiunian et al., 1997).
 | |
| Fig. 1: | Structure of resveratrol |
A potent scavenger of free radical species may serve as a possible preventive intervention for free radical-mediated diseases (Ames et al., 1995). In recent times therefore, the search for natural antioxidants and other preparations of plant origin to achieve this objective has been intensified. Phytoalexins are low molecular weight secondary metabolites made by plants as a defense response to microbial infections (Soleas et al., 1997). Resveratrol (3,5,4'-trihydroxy-trans-stilbene), a phytoalexin that belongs to the groups of compounds known as stilbenes, can be found in dietary items including red wine, grapes and peanuts. It has been shown to confer many beneficial effects in humans and animal models and it is being studied as a chemopreventive agent for cancer, probably due its antioxidant and anti-proliferative properties (Jang et al., 1997; Lin and Tsai, 1999; Fremont et al., 2000). Figure 1 shows the structure of resveratrol.
The present study was designed to assess the efficacy of resveratrol supplementation in rats administered with potent necrogenic carcinogen 1,2-dimethylhydrazine mediated hepatic oxidative stress.
MATERIALS AND METHODS
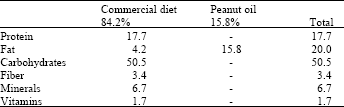
Animals and diet: Five weeks old, male Wistar rats were purchased from Central Animal House, Rajah Muthiah Institute of Health Sciences, Annamalai University, Tamilnadu, India, were quarantined for one week and allocated randomly to experimental or control groups. Commercial pellet diet containing 4.2% fat (Hindustan Lever Ltd, Mumbai, India) was powdered and mixed with 15.8% peanut oil making a total of 20% fat in the diet (Table 1). This modified powdered pellet diet was fed to rats in all groups throughout the experimental period of 31 weeks (including 1 week for acclimatization). The total caloric intake by the rats in all the groups was adjusted to be the same. Animals were maintained as per the principles and guidelines of the Ethical Committee of Animal Care of Annamalai University in accordance with the Indian National Law on animal care and use (Reg. No.160/1999/CPCSEA).
| Table 1: | Composition of the diet |
 | |
The animals were housed four per cage in a specific-pathogen free animal room under controlled conditions of a 12 h light/12 h dark cycle, with temperature of 22±1°C and relative humidity of 50±10% till the end of 30 weeks.
Chemicals: 1,2-dimethylhydrazine and trans-resveratrol were purchased from Sigma Chemical Company, St. Louis, MO, USA. All other chemicals used were of analytical grade and obtained from Hi-Media Laboratories, Mumbai.
Carcinogen and chemopreventive agent administration: All animals in groups 3-6 received 20 mg kg-1 body weight DMH injection once a week subcutaneously for the first 15 weeks. Prior to injection, DMH was dissolved in EDTA (1 mmol), the pH was adjusted to 6.5 with NaOH (1 mmol) and used immediately. Due to its low solubility in water, trans-resveratrol (8 mg kg-1 body weight) was suspended in 1% carboxymethylcellulose (CMC) and given orally using an intragastric tube.
Experimental design: The animals were randomly assigned to the six experimental groups (six animals per group). Initial body weights of all animals in this study protocol were ensured between 120-130 g. Food consumption and animal body weight were monitored weekly throughout the experimental period of 30 weeks. For more clarity experimental design is shown in Fig. 2.
| Group 1 (Control) | Rats received modified diet along with intragastric intubation of 1% CMC (1 mL), throughout the experiment. |
| Group 2 (Control + RES) | Rats received modified diet + 8 mg kg-1 body weight resveratrol suspended in 1% CMC (1 mL), p.o. everyday throughout the experiment. |
| Group 3 body (DMH) | Rats were administered 20 mg kg-1 weight DMH (carcinogen) s.c. once a week for first 15 consecutive weeks and kept with out any treatment for the next 15 weeks. |
| Group 4 DMH + RES (I) | The animals were supplemented with resveratrol (8 mg kg-1 body weight, p.o.) everyday starting 2 weeks before carcinogen treatment for the first 15 weeks and kept with out any treatment for the next 15 weeks. (initiation-I) |
| Group 5 DMH + RES (PI) | The animals were supplemented with resveratrol (8 mg kg-1 body weight, p.o.) two days after the last injection of the carcinogen and continued till the end of the experiment (post initiation-PI) |
| Group 6 DMH + RES (EP) | The animals were supplemented with resveratrol (8 mg kg-1 body weight, p.o.) from the day of carcinogen treatment and continued till the end of the entire experimental period of 30 weeks (entire period-EP) |
Animal autopsy and tissue preparation: At the end of 30 weeks all the animals were sacrificed under anesthesia (i.p. administration of ketamine hydrochloride, 30 mg kg-1 body weight), by cervical dislocation between 8 am to 10 am after an overnight fast. The colon was examined grossly and histologically for the number and type of tumors. The entire study was conducted in the Department of Biochemistry and Biotechnology, Annamalai University, Annamalainagar, Tamilnadu, India.
Preparation of liver homogenate: Liver sections were homogenized in ice-cold phosphate buffer (pH 7.0, 0.01 M) with a Teflon pestle in a glass tube. Lipid peroxidation in fresh whole tissue homogenate was assessed by measuring CD by the method of Recknagel and Glende (1984), LOOHs and thiobabituric acid reactive substances (TBARS) by Jiang et al. (1992) and Okhawa et al. (1979), respectively.
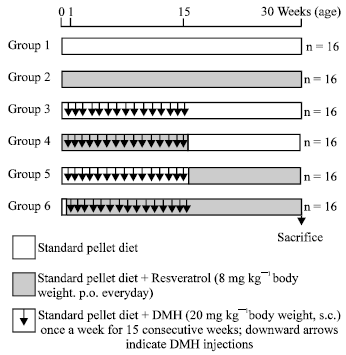 | |
| Fig. 2: | The experimental design is figuratively represented for clarity |
The activities of SOD (EC 1.15.1.1) and CAT (EC 1.11.1.6) were assessed in the liver homogenates as described previously (Kakkar et al., 1984; Sinha, 1972). Hepatic GSH, Gpx (EC 1.11.1.9), GST (EC 2.5.1.18) and GR (EC 1.6.4.2) activities were determined spectrophotometrically by the method of Boyne and Ellman (1972), Flohe and Gunzler (1984), Habig and Jakoby (1981) and Carlberg and Mannervik (1985), respectively.
Vitamin C and E was determined by the method of Omaye et al. (1979) and Desai (1971), respectively. Protein content was determined by the method of Lowry et al. (1951) using bovine serum albumin as standard at 660 nm.
Statistical analysis: Values are expressed as means±SD of 6 animals in each group. Data within and between the groups were analyzed using one-way analysis of variance (ANOVA) followed by Duncan’s Multiple Range Test (DMRT) using SPSS version 11.0 software (SPSS Inc., Chicago, IL, USA).
RESULTS
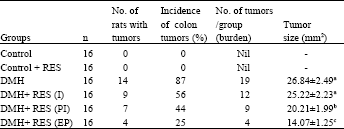
Effect of resveratrol on DMH-induced colonic tumor burden: No tumors were observed in group 1 and 2 animals. In group 3 rats the tumor incidence was 87%, which was significantly higher than resveratrol supplemented groups 4, 5 and 6 (56, 44 and 25%, respectively). The average size of the tumor was 26.84 mm2 in group 3. Entire period resveratrol supplementation to DMH-exposed rats (Group-6) significantly reduced tumor size to 14.07 mm2 (Table 2).
Effect of resveratrol on hepatic lipid peroxidation: Liver CD, LOOHs and TBARS was significantly elevated (p<0.05), in the DMH-treated rats (Group 3) at the end of 30 weeks as compared to the control rats (Group 1). Resveratrol supplementation during the initiation (Group 4) stages of carcinogenesis did not have a significant effect on hepatic CD, LOOHs and TBARS as compared to the unsupplemented DMH-treated rats (Group 3).

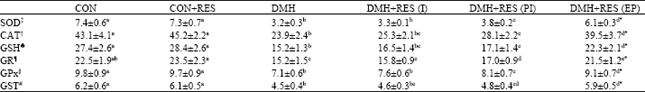
| Table 4: | Effect of resveratrol on hepatic antioxidants system |
 | |

On the other hand supplementing resveratrol in post-initiation (Group 5) and throughout the entire period of the study (Group 6) significantly lowered liver CD, LOOHs and TBARS (p<0.05) as compared to the DMH-alone treated rats (Group 3). This tendency was more pronounced when resveratrol was supplemented in the entire period of the study (Group 6) (Table 3).
Effect of resveratrol on hepatic enzymic and non-enzymic antioxidants: SOD, CAT, GPx, GST, GR activities and the levels of GSH, Vitamin C and E were significantly decreased (p<0.05) in the liver of DMH treated rats (Group 3) as compared to the control rats (Group 1). Resveratrol supplementation during the initiation (Group 4) stage of carcinogenesis did not have a significant effect on these parameters as compared to the unsupplemented DMH-treated rats (Group 3) (Table 4 and 5). But supplementing resveratrol during the post-initiation stage as well as throughout the entire experimental period significantly elevated the liver SOD, CAT, GPx, GST, GR activities (p<0.05) and GSH, Vitamin C and E levels (p<0.05) as compared to the DMH-alone exposed rats (Group 3). The effect of resveratrol in restoring tissue antioxidant status was more pronounced in group 6 (p<0.01) as compared to DMH-alone treated rats (Table 4 and 5).
DISCUSSION
Colon carcinogenesis is a multistep process and is thought to arise from the accumulation of mutation involving a variety of oncogenes and tumor suppressor genes in a single epithelial cell of the colon and rectum (Fearon and Vogelstein, 1990). Recently, a general scheme has been proposed for the role of oxy-radicals in initiation and promotion stages of carcinogenesis. Free radical mediated mechanisms represent a significant pathway for metabolic activation of carcinogens (Fiala, 1977). DMH undergoes metabolic activation in the liver to produce active electrophilic carbonium ion through several process, which are known to elicit oxidative stress. Carbonium ion methylate DNA bases, induces point mutations, micronuclei and sister chromatid exchanges leading to colon specific carcinogenesis (Fujita et al., 2004).
Many reports are available regarding the role of free radicals and lipid peroxidation at the tissue level as a cause of cancer (Masotti et al., 1988). The recent evidence indicates that generation of active oxygen species and formation of reactive products may be involved in various carcinogenic processes (Fang et al., 2002). Cells are equipped with an impressive repertoire of antioxidant enzymes as well as non-enzymic small antioxidant molecules (Sun, 1990). SOD, CAT and GPx are considered to be the primary antioxidant enzymes, since they are involved in the direct elimination of reactive oxygen species. GST and GR are redox-regulating enzymes, which help in the detoxification of reactive oxygen species by decreasing peroxide levels or by maintaining a steady supply of metabolic intermediates like GSH for the primary defense enzymes. The non-enzymic small molecular antioxidants include sulphydryl compounds such as GSH, ascorbate (Vitamin C) and α-tocopherol (Vitamin E). Since these antioxidants work co-operatively, a change in any one of them may break the equilibrium and cause cell damage leading to malignancy (Navarro et al., 1999).
Resveratrol, a polyphenolic compound has received wide attention lately due to its ability to serve as an effective anti-oxidative agent (Fremont et al., 2002). It inhibits cellular events associated with tumor initiation, promotion/progression (Cadenas and Barja, 1999). The results described here clearly indicate that the entire period supplementation of dietary resveratrol attenuated hepatic lipid peroxidation and enhances antioxidant defense system in tumor bearing animals with concomitant inhibition of tumor formation in DMH-induced colon carcinogenesis.
DMH administration decreases the hepatic SOD and CAT, the primary defense against oxidative damage of tissues. Increased exposure to radicals or from impaired efficiency of these protective enzymes leads to diseases including cancer. Resveratrol supplementation significantly enhances the liver SOD and CAT activities, the increased cellular capacity to neutralize ROS may account to some extent for the anticancer activity of resveratrol.
It is generally accepted that reduced GSH and GSH dependent enzymes have an important role in the protection of mammalian cells against oxidative and alkylating agents. Chemical carcinogen, DMH requires metabolic activation in order to exert their mutagenic and carcinogenic effects. The GST and GSH play an important role in the detoxification of many environmental carcinogens (Hayes and McLellan, 1999). The depletion in hepatic GSH level and decrease in GST activity could be responsible for the changing effectiveness of alkylating agents by decreasing the conjugation with GSH and could enhance the mutagenic and carcinogenic effects of these agents.
GPx is an enzyme plays a very important protective role by removing hydrogen peroxide and lipidic (organic) hydroperoxides. Thus, the reduction in GPx activity would cause the formation of potential carcinogenic toxic compounds. GR, a glutathione regenerating enzyme plays a major role to restore the depleted GSH during carcinogen exposure (Navarro et al., 1999). Vitamin C and E plays a significant role in reducing the oxidative stress during carcinogen exposure (Nordberg et al., 2001). In our study on DMH treatment the hepatic GSH, GSH-dependent enzymes, Vitamin C and E levels was found to be reduced, which in turn increases the carcinogenicity of DMH. Resveratrol supplementation has been show to enhance GSH levels, GPx, GST and GR activities in the liver. Resveratrol ameliorate the hepatic damage induced by DMH possibly by (1) enhancing GSH levels by induction of GSH synthesizing enzymes such as γ-glutamylcysteine synthetase and GSH synthetase (2) increasing adaptive response in order to compensate the depleted GSH levels by reduction of oxidized glutathione (GSSH) to GSH by GR (3) increasing the rate of free radical utilization via GST and GPx system (4) inhibiting lipid peroxidation by elevation of hepatic Vitamin C and E levels.
In conclusion resveratrol a natural compound with profound pharmacological and biological activity could ameliorate the hepatic damage induced by DMH.
ACKNOWLEDGMENT
The financial support by Lady Tata Memorial Trust, Mumbai in the form of Senior Research Fellowship to M. Sengottuvelan is gratefully acknowledged.
REFERENCES
- Ames, B.N., L.S. Gold and W.C. Willet, 1995. The causes and prevention of cancer. Proc. Natl. Acad. Sci. USA., 92: 5258-5265.
PubMedDirect Link - Arutiunian, A.V., V.M. Prokopenko, S.O. Burmistrov, T.I. Oparina and E.V. Frolova et al., 1997. Free-radical processes in blood serum, liver and large bowel during 1,2-dimethylhydrazine-induced carcinogenesis in rats. Vopr Onkol., 43: 618-622.
PubMedDirect Link - Boyne, A.F. and G.L. Ellman, 1972. A methodology for analysis of tissue sulfhydryl components. Anal. Biochem., 46: 639-653.
CrossRefDirect Link - Cadenas, S. and G. Barja, 1999. Resveratrol, melatonin, vitamin E and PBN protect against renal oxidative DNA damage induced by the kidney carcinogen KBrO3. Free Radical Biol. Med., 26: 1531-1537.
CrossRefDirect Link - Fang, Y.Z., S. Yang and G. Wu, 2002. Free radicals, antioxidants, and nutrition. Nutrition, 18: 872-879.
CrossRefDirect Link - Farber, E. and D.S. Sarma, 1987. Hepatocarcinogenesis: A dynamic cellular perspective. Lab. Invest., 56: 4-22.
PubMedDirect Link - Fearon, E.R. and B. Vogelstein, 1990. A genetic model for colorectal tumorigenesis. Cell, 61: 759-767.
PubMedDirect Link - Freig, D.I., T.M. Reid and L.A. Loeb, 1994. Reactive oxygen species in tumorigenesis. Cancer Res., 54: 1890-1894.
Direct Link - Fiala, E.S., 1977. Investigations into the metabolism and mechanism of action of the colon carcinogens 1,2-dimethylhydrazine and azoxymethane. Cancer, 40: 2436-2445.
PubMed - Flohe, L. and W.A. Gunzler, 1984. Assays of glutathione peroxidase. Methods Enzymol., 105: 114-121.
CrossRefPubMedDirect Link - Fremont, L., 2000. Biological effects of resveratrol. Life Sci., 66: 663-673.
CrossRefPubMedDirect Link - Fujita, K., E. Matsuda, K. Sekine, M. Iigo and H. Tsuda, 2004. Lactoferrin enhances Fas expression and apoptosis in the colon mucosa of azoxymethane-treated rats. Carcinogenesis, 25: 1961-1966.
CrossRefDirect Link - Habig, W.H. and W.B. Jakoby, 1981. Glutathione S-Transferases (rat and human). Methods Enzymol., 77: 218-231.
PubMedDirect Link - Hayes, J.D. and L.I. McLellan, 1999. Glutathione and glutathione-dependent enzymes represent a co-ordinately regulated defence against oxidative stress. Free Radical Res., 31: 273-300.
CrossRefDirect Link - Hayes, M.A., T.H. Rushmore and M.Y. Goldberg, 1987. Inhibition of hepatocarcinogenic responses to 1,2-dimethylhydrazine by diallyl sulfide, a component of garlic oil. Carcinogenesis, 8: 1155-1157.
Direct Link - Jang, M., L. Cai, G.O. Udeani, K.V. Slowing and C.F. Thomas et al., 1997. Cancer chemopreventive activity of resveratrol, a natural product derived from grapes. Science, 275: 218-220.
CrossRefPubMedDirect Link - Jiang, Z.Y., J.V. Hunt and S.P. Wolff, 1992. Ferrous ion oxidation in the presence of xylenol orange for detection of lipid hydroperoxide in low density lipoprotein. Anal. Biochem., 202: 384-389.
CrossRefDirect Link - Kakkar, P., B. Das and P.N. Viswanathan, 1984. A modified spectrophotometric assay of superoxide dismutase. Indian J. Biochem. Biophys., 21: 130-132.
PubMedDirect Link - Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - Masotti, L., E. Casali and T. Galeotti, 1988. Lipid peroxidation in tumour cells. Free Radic. Biol. Med., 4: 377-386.
Direct Link - Matsumoto, H. and H.H. Higa, 1966. Studies on methylazoxymethanol, the aglycone of cycasin: Methylation of nucleic acids in vitro. Biochem. J., 98: 20C-22C.
Direct Link - Navarro, J., E. Obrador, J. Carretero, I. Petschen, J. Avino, P. Perez and J.M. Estrela, 1999. Changes in glutathione status and the antioxidant system in blood and in cancer cells associate with tumour growth in vivo. Free Radic. Biol. Med., 26: 410-418.
PubMed - Ohkawa, H., N. Ohishi and K. Yagi, 1979. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem., 95: 351-358.
CrossRefPubMedDirect Link - Ohno, K., S. Narushima, S. Takeuchi, K. Itoh, T. Itoh, K. Hioki and T. Nomura, 2001. Effect of bacterial metabolism in the intestine on colorectal tumors induced by 1,2-dimethylhydrazine in transgenic mice harboring human prototype c-Ha-ras genes. J. Exp. Clin. Cancer Res., 20: 51-56.
PubMedDirect Link - Omaye, S.T., J.D. Turnbull and H.E. Sauberlich, 1979. Selected methods for the determination of ascorbic acid in animal cells, tissues and fluids. Methods Enzymol., 62: 3-11.
CrossRefPubMedDirect Link - Recknagel, R.O. and E.A. Glende Jr., 1984. Spectrophotometric detection of lipid conjugated dienes. Methods Enzymol., 105: 331-337.
CrossRefPubMedDirect Link - Sinha, A.K., 1972. Colorimetric assay of catalase. Anal. Biochem., 47: 389-394.
CrossRefPubMedDirect Link - Soleas, G.J., E.P. Diamandis and D.M. Goldberg, 1997. Resveratrol: A molecule whose time has come? And gone?. Clin. Biochem., 30: 91-113.
PubMedDirect Link - St Clair, W.H., P.C. Billings, J.A. Carew, C.K. Mc Gandy, P. Newberne and A.R. Kennedy, 1990. Suppression of dimethylhydrazine-induced carcinogenesis in mice by dietary addition of the Bowman-Birk protease inhibitor. Cancer Res., 50: 580-586.
PubMedDirect Link - Weed, H.G., R.B. Mc Gandy and A.R. Kennedy, 1985. Protection against dimethylhydrazine-induced adenomatous tumors of the mouse colon by the dietary addition of an extract of soybeans containing the Bowman-Birk protease inhibitor. Carcinogenesis, 6: 1239-1241.
PubMedDirect Link - Ying, T.S., D.S. Sarma and E. Farber, 1979. Induction of presumptive preneoplastic lesions in rat liver by a single dose of 1,2-dimethylhydrazine. Chem. Biol. Interact., 28: 363-366.
PubMedDirect Link