
Dwi Sutiningsih
Department of Epidemiology and Tropical Disease, Faculty of Public Health, Diponegoro University, Semarang, Indonesia
LiveDNA: 62.16151
Mohamad Arie Wuryanto
Department of Epidemiology and Tropical Disease, Faculty of Public Health, Diponegoro University, Semarang, Indonesia
Henry Setyawan Susanto
Department of Epidemiology and Tropical Disease, Faculty of Public Health, Diponegoro University, Semarang, Indonesia
Sujud Hariyadi
Department of Epidemiology and Tropical Disease, Faculty of Public Health, Diponegoro University, Semarang, Indonesia
Mustofa
Department of Pharmacology, Faculty of Medicine, University of Gadjah Mada, Yogyakarta, Indonesia
International Journal of Cancer Research
Year: 2020 | Volume: 16 | Issue: 1 | Page No.: 18-27







ABSTRACT
Background and Objective: Linamarin is an active compound isolated from the leaves of cassava (Manihot esculenta Cranz) that has cytotoxic effects on HT-29, MCF-7 and HL-60 cells. This study was aimed to determine the cytotoxic and antiproliferation activity and induction of p53 protein in Raji cells after administration of various concentrations of linamarin from cassava leaves (Manihot esculenta Cranz). Materials and Methods: Linamarin was isolated from cassava leaves (Manihot esculenta Cranz) using a multilevel purification method. Linamarin cytotoxicity was tested on Raji cells using the MTT method, while antiproliferation activity was tested using a doubling time test. The p53 protein expression was observed by immunocytochemical tests. The cytotoxic activity of Raji cells was expressed by the value of Inhibitory concentration 50 μg mL–1. The doubling time was calculated by comparing the slope values of the log graphs of the number of cells at various times. Raji cells that were positive for p53 protein showed brown painted nuclei or cytoplasm. Results: Linamarin from cassava leaves can inhibit cytotoxic activity and proliferation on Raji cells. The higher the linamarin concentration, the longer the doubling time of Raji cells. The expression of p53 protein on Raji cells after linamarin administration was higher than the control. The p53 protein expression was found in the nuclei (91.05%) and cytoplasm (8.95%). Conclusion: Based on those findings, linamarin from cassava leaves has the potential to be developed as an anticancer agent.
PDF Abstract XML References Citation
How to cite this article
Dwi Sutiningsih, Mohamad Arie Wuryanto, Henry Setyawan Susanto, Sujud Hariyadi and Mustofa, 2020. Anticancer Activity of Linamarin from Cassava Leaves (Manihot esculenta Cranz) on Raji Cells. International Journal of Cancer Research, 16: 18-27.
DOI: 10.3923/ijcr.2020.18.27
URL: https://scialert.net/abstract/?doi=ijcr.2020.18.27
DOI: 10.3923/ijcr.2020.18.27
URL: https://scialert.net/abstract/?doi=ijcr.2020.18.27
INTRODUCTION
Obstacles and side effects caused by various cancer treatments have necessitated the discovery of highly effective alternatives with minimal side effects. One such effort is the development of drugs from plants that contain anticancer compounds. The development of cancer drugs from plants has several advantages, among which are their low cost, availability and relatively few side effects1.
In Indonesia, cassava has considerable economic value compared to other tubers. Not only is cassava (Manihot esculenta Cranz) one of the world’s principal food staples after grains and corn1, their leaves, widely consumed in Indonesia and elsewhere, are rich in vitamins A, C, K, among others and minerals, including iron, calcium and phosphorus. The energy content of cassava leaves is greater than most other green vegetables2. Cassava also contains cyanogenic glucoside compounds, which consist of linamarin and lotaustrain3,4 at a ratio of 10:1. Linamarin has potential use as an anti-neoplastic compound5,6. The mechanism of linamarin in the treatment of cancer using linamarase gene therapy has been investigated.
Meanwhile the Idibie et al.6 study states that linamarin in root tubers has been proven in vitro to have cytotoxic effects on HT-29, MCF-7 and HL-60 cells. From the results of this study, Inhibitor Concentration 50 (IC50) was obtained in the amounts of >300 μg mL–1, 235.96±9.87 and 246.51±10.12 μg mL–1 after incubation for 48 h. In this study, linamarin was obtained from cassava leaf extracted with methanol. The study of Yusuf et al.7 using linamarin isolated from cassava leaves also showed cytotoxic effects on Caov-3 cells and Hela cells. The IC50 value of the two cell lines is 38 and 57 μg mL–1, respectively. Cancer cell death has been caused by the linamarin content found in cassava plants8-11. Carotene and vitamin C compounds found in cassava leaves are thought to have anticancer properties12-15. Research by Enger et al.16, stated that carotene is protective toward colon adenoma rather than other carotenoids in the early stages of tumor formation. Kontek et al.17 stated that vitamin C had a positive effect on the damage level of oxidative DNA in colon cancer cells.
The benefits of cassava as an anticancer agent have been proven in several cancer cells, but have not yet been widely studied regarding its potential in Raji cells. This study was aimed to determine cytotoxic and antiproliferative activity and induction of p53 protein in Raji cells following treatment of linamarin from cassava leaves.
MATERIALS AND METHODS
Cassava leaves (Manihot esculenta Cranz) were obtained from the local market in Yogyakarta, Indonesia and then identified at the Laboratory of Pharmaceutical Biology, Faculty of Pharmacy, Gadjah Mada University. This research project was conducted from June 4, 2018 to December 4, 2018. Raji cells were obtained from the collection of the Laboratory of Parasitology, Faculty of Medicine, Gadjah Mada University. This cell is a continuous cell line that grows floating, similar to lymphoblast cells (B lymphocytes) from Burkitt's lymphoma infected by Epstein-Barr Virus (EBV). Materials for growing Raji cells are RPMI solution (Sigma-Aldrich, Saint Louis, Missouri, USA), Dulbecco's Modified Eagle's Medium (DMEM) (Sigma-Aldrich, Saint Louis, Missouri, USA), HEPES, Fetal Bovine Serum (FBS) (Gibco, Grand Island, N.Y, USA), penicillin-streptomycin) (Gibco, Grand Island, N.Y, USA), DMSO (Sigma-Aldrich, Saint Louis, Missouri, USA), tripan blue (E-Merck, Darmstadt, Germany) and 3-(4-, 5 dimethylthiazol-2-yl) -2.5-diphenyl tetrazolium bromide (MTT) (Sigma-Aldrich, Saint Louis, Missouri, USA).
Linamarin isolation from cassava leaves: A 5 g batch of cassava leaves was cut into small pieces and then pounded in a mortar. The result was blended thoroughly with a total of 10 ml of 0.1 M HCl solution. The mixture solution was centrifuged at 3500 rpm to obtain the supernatant. The supernatant liquid obtained was transferred to the Falcon tube (Thermo Fisher Scientific, Waltham, MA USA). The supernatant liquid mixture with 0, 1 M HCl was linamarin extract of cassava leaf, which was then isolated. Finally, the linamarin extract was frozen at -20°C18.
Cytotoxic test on Raji cells: The cytotoxicity test was done colorimetrically using MTT reagents (Sigma-Aldrich, Darmstadt, Germany). Linamarin of 10 μL at various concentrations was added to Raji cell culture the day after transplantation. The concentration of linamarin used for treatment of Raji cells was 31.25, 62.5, 125, 250, 500 and 1000 μg mL–1. Cells that were not treated were used as controls. On the third day, 20 μL of MTT reagent was added to approximately 5 mg mL–1 per well. After four hours of incubation, 100 μL of 0.1 N HCl-isopropanol was added to each well to dissolve the formazan crystals that had formed. Absorbance (A) was measured using a microplate reader at a wavelength of 595 nm. All steps were carried out three times.
Antiproliferation test (doubling time) in Raji cells: Cells were fasted for 24 h in culture media containing 0.5% of FBS. Afterwards, they were grown in a plate with a medium added, with linamarin at a non-lethal concentration of three series below the IC50 value. Then it was incubated in a 5% CO2 incubator (Nuaire, Plymouth, MN 5547, USA) at 37°C for 24, 48 and 72 h. Each well was calculated by the number of cells living using hemocytometrics (Neubauer haemocytometry, EMS, 1560 Industry Road, Hatfield, PA).
Immunocytochemical test in Raji cells: Immuno-cytochemical staining was performed using the avidin-biotin-peroxidase complex with monoclonal antibodies against p53 according protocols of p53 Assay kit (Colorimetric) (NCL-L-p53-DO7, Novocastra, Benton Lane, Newcastle, United Kingdom)). In a microculture, 96 wells containing 100 μL of test cells, with a density of 2×104 cells/well, 100 μL of the test compound were added at concentrations of 10 μg mL–1. They were then incubated with 5% CO2 flow at 37°C for 24 h. After being incubated overnight, 200 μL of cells from each well were taken and inserted in eppendorf tubes, then centrifuged to 1200 rpm×5 min. The supernatant liquid was removed, leaving the pellet and then re-suspended. The cell suspension was extracted and placed on a glass object that had been coated with poly-lysine. The cells were fixated with acetone for 10 min. Later, they were washed with PBS (Phosphate Buffered Saline)(E. Merck, Darmstadt, Germany)×5 min and etched with hydrogen peroxidase 0, 1% for 10 min. After washing them with running water, they were rinsed with PBS for 5 min, dripped with 100 μL normal horse serum for 10 min and cleaned without water. Finally, they were dripped with anti-p53 protein primary antibodies (Novocastra, Benton Lane, Newcastle, United Kingdom) and left for 24 h.
The next day the suspension was:
| • | Washed twice with PBS×5 min each |
| • | Dripped with biotinylated secondary antibodies (Novocastra)×10 min |
| • | Washed×2 with PBS×5 min each |
| • | Dripped then incubated with Avidin Biotin reagent enzyme (Novocastra)×10 min |
| • | Washed×2 with PBS×5 min each |
| • | Incubated with a peroxidase substrate (DAB) (Novocastra) ×10 min or until the coloring appeared |
| • | Washed with running water |
| • | Counter stained with hematoxylin for 10-20 sec, then washed with running water |
| • | Dehydrated using 95% ethanol and xylene ×10 min each |
| • | The mounting media was dripped and then covered with a glass deck |
The results were observed under a light microscope (Olympus, Japan) with 400× magnification. Cells positive for p53 protein showed nuclei or cytoplasm painted brown.
Data analysis: Raji cell cytotoxicity was analyzed using probit analysis to determine 50% Inhibition Concentration (IC50). Probit analysis was obtained from the conversion of the percentage of inhibition to the probit value. Percentage of inhibition was calculated as follows19:
where, ΣA is the number of living cells in untreated controls, ΣB is the number of living cells due to the treatment of compounds at various concentrations.
The difference in percentages of cell inhibition between each treatment group was tested statistically using a one-way ANOVA test with 95% Confidence Interval. Analysis of doubling times was calculated by comparing the slope of the log graphs of the number of cells at different observation times. To find differences between groups, the average number of cells living at the various times was analyzed statistically using the one-way ANOVA test with a 95% confidence level. Expression of p53 protein was analyzed by observing its percentages as expressed in Raji cells after immuno-histochemical treatment. Cells that were positively stained with p53 protein showed nuclei or cytoplasm painted brown. The proportion of cells that were positively p53 protein was determined by calculating the presence of stained nuclei or cytoplasm per 100 cells.
RESULTS AND DISCUSSION
Linamarin cytotoxic activity in Raji cells: Cytotoxic activity was tested to determine the toxicity of a linamarin compound on Raji cells. Raji cells are continuous cell lines that grow floating and unattached to the bases of flasks. The cell is similar to lymphoblast cells (B lymphocytes) from Burkitt's lymphoma infected by Epstein-Barr Virus (EBV). The cells are round and clustered. Living cells will appear bright under a contrast phase microscope while dead cells will appear dark.
The parameters used to express the potency of linamarin toxicity from cassava leaves were IC50 values. The results of calculating cell inhibition percentage of Raji cells after linamarin administration from cassava leaves are presented in Table 1. Table 1 shows that at the highest linamarin concentration (1000 μg mL–1), the percentage of Raji cell inhibition was 97.550%, while at the lowest concentration (31.25 μg mL–1), the percentage was 27.194%.
| Table 1: | Average number of living cells vs. percentage of Raji cell inhibition after administration of various concentrations of linamarin |
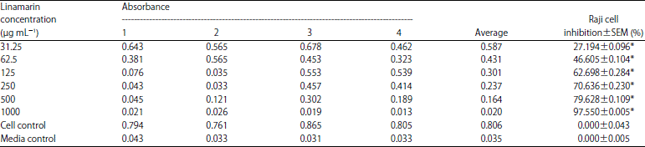 | |
| *p<0.05 with one-way ANOVA test, SEM: Standard error of the mean | |
| Table 2: | Doubling time of Raji cells after treatment with various concentrations of linamarin vs. control |
 | |
| Table 3: | p53 protein (%) expression on Raji cell control and linamarin concentrations of 62.5 and 31.25 μg mL–1 |
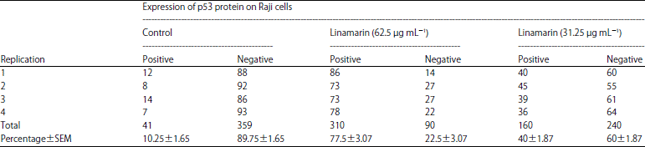 | |
| SEM: Standard error of the mean | |
The results of Kolmogorov-Smirnov's analysis showed that the average Raji cell inhibition was normally distributed (p = 0.135), while homogeneity test results were homogeneous (p = 0.088). The one-way ANOVA test was used to determine the differences in Raji cell inhibition between various linamarin treatments. The results of the one-way ANOVA analysis revealed significant differences between the Raji cell inhibition levels at various linamarin concentrations (p = 0.000).
Antiproliferation activity of linamarin in Raji cells: The concentration of the test compound used in the doubling time test was three concentrations below the IC50 value (15.63, 31.25 and 62.50 μg mL–1). Cell counts are carried out at 0, 24, 48 and 72 h. Raji cells had been previously fasted (starved) for 24 h using RPMI 1640 media containing FBS 0.5%. Data of doubling time analysis of Raji cells after linamarin treatment and control (without treatment) can be seen in Table 2.
Data from Table 2 shows how the multiplication times of Raji cells after linamarin treatment, at concentrations below the IC50 value, run greater than the control times. Linamarin concentration of 62.50 μg mL–1 can delay the doubling times of Raji cells by±2×those of the Raji control cells.
From Fig. 1, it can be seen that at 30 min after the treatment of the test compound, there has been no inhibition of Raji cell growth, in contrast to observations at 24, 48 and 72 h. One-way ANOVA analysis showed that there were significant differences (p = 0.023) in the average number of living Raji cells, dependent upon the elapsed time post-linamarin treatment (24, 48 and 72 h).
Expression of p53 protein on Raji cells: The immunocytochemical test results showed that linamarin can increase the expression of p53 protein on Raji cells. Complete results of p53 protein expression tests are presented in Table 3.
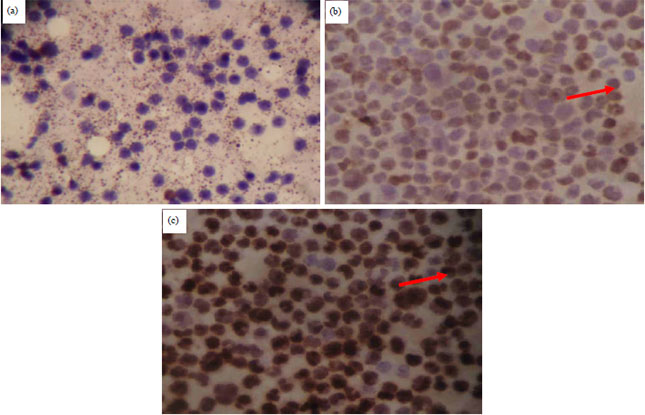 | |
| Fig. 1:(a-c): | Microscopic photograph of Raji, (a) Control cells (without treatment), (b) Cells treated with linamarin 32.5 μg mL–1 and (c) Cells treated with linamarin 62.5 μg mL–1 with immunocytochemical staining (magnification 400X) |
| Positive cells with expression of p53 protein have brown nuclei or cytoplasm, cells that are negative for p53 protein expression have purple nuclei or cytoplasm | |
| Table 4: | Location of p53 protein expression of Raji cells control and linamarin concentrations of 62.5 and 31.25 μg mL–1 |
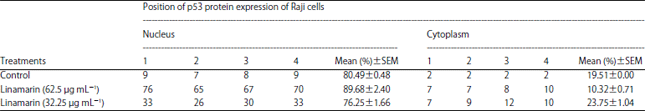 | |
| SEM: Standard error of the mean | |
According to those results, there is a tendency for greater p53 protein expression in the treatment group compared to the control group. Linamarin concentration of 62.5 μg mL–1 showed increased positive p53 protein expression in Raji cells by 77.5±3.07%, while linamarin concentration was 31.25 μg mL–1 at 40±1.87%. The one-way ANOVA test results showed a significant difference in the number of p53 protein expression in Raji cells at various linamarin concentrations (p = 0.000). Details pertaining to the expression of p53 protein in the nuclei and cytoplasm of Raji cells are presented in Table 4 and Fig. 1.
From Fig. 1, it can be seen that in the Raji control cell there was a tendency to decrease the positive p53 protein expression, whereas in the Raji cells with linamarin, 32.5 and 62.5 μg mL–1 concentrations appeared to increase positive p53 protein expression, with most located in the nuclei (Table 4).
Linamarin cytotoxic activity in Raji cells: The cytotoxicity test determined the value of IC50, which is a concentration capable of inhibiting cell growth, such as Raji cells, by up to 50%. The smaller the IC50 value, the more toxic the compound is. The potential for linamarin toxicity from cassava leaves (Manihot esculenta Cranz) to Raji cells is indicated by IC50 values of 71.865±0.229 μg mL–1. At its highest concentration (1000 μg mL–1), the percentage of Raji cell growth inhibition was 97.550±0.005%, while the lowest concentration of linamarin (31.25 μg mL–1) was 27.194±0.096% (Table 1). It shows that at the higher the concentration of linamarin, the greater the percentage of Raji cell growth inhibition, with a significant statistical difference (p <0.05). This proves that linamarin obtained from cassava leaves (M. esculenta Cranz) can suppress the growth of Raji cancer cells. Linamarin is found in all parts of cassava plants (M. esculenta Cranz), but most abundantly at the roots, leaves and root tuber skin5.
Yusuf et al.7 found that linamarin from cassava leaves can inhibit the growth of Caov-3 cancer cells and Hela cells with IC50 values of 38 and 57 μg mL–1, respectively. Idibie et al.6 in his research, stated that IC50 values decreased when pure linamarin compounds and crude extracts of cassava tubers were given along with linamarase enzymes on MCF-7 cancer cells (adenocarcinoma breast cancer), HT-29 (adenocarcinoma colon) and HL-60 (cell line leukemia). Meanwhile, the IC50 values of crude extracts are higher than linamarin if not given along with the linamarase enzyme. Likewise, the results of Al Fourjani's5 study showed that the IC50 values of MCF cells after treatment with raw cassava leaf extract and boiled cassava leaves were 63.1 and 79.4 μg mL–1, respectively.
Crude extracts are said to have strong potential as anticancer agents if the IC50 value20 is <30 μg mL–1. The results of this study showed IC50 value of Raji cells after linamarin administration to be greater than 30 μg mL–1. In fact, they registered as high as 71.865±0.229 μg mL–1, meaning that the potency of linamarin toxicity in active Raji cells was weaker, or only moderately active (30<IC50<100 μg mL–1). This was presumably due to differences in the characteristics of cancer cells used in the study.
Raji cells are found in the Burkitt's lymphoma cell line in humans. Burkitt's lymphoma at the molecular level is characterized by synergistic Bcl-2 and c-myc expressions. C-Myc is upregulation Bcl-2, so the increase in c-myc expression can also increase the expression of Bcl-2. As a result of this increase in expression, cells do not experience apoptosis21,22. Burkitt's lymphoma has chromosome translocation that activates c-myc. In some patients it also shows the occurrence of mutations in p53 which result in the inhibition of the apoptotic process in these cancer cells. Activating p16INK4a resulted in loss of CDK inhibitory function, diminishing loss of cell control of its growth. Changes (mutations) also occur in the expression of pRb and p53, which are gene suppressor tumors and in other genes, such as Bax, p73 and Bcl-6, which provide sufficient growth signals and inhibit apoptosis in cancer cells23-25. Mutations also occur in downstream Caspase-3 which causes Raji cells to be resistant to apoptosis26,27.
The protein expression of the Epstein-Barr Nuclear Antigen 1 (EBNA1) in Burkitt's lymphoma, infected by Epstein-Barr Virus (EBV), can also inhibit the occurrence of apoptosis in cancer cells28. Through this mechanism, it is suspected that Raji cells can avoid the apoptotic mechanism triggered by linamarin compounds from cassava leaves. This is why the suspected cause of cassava leaf extract cytotoxicity against Raji cells is considered moderate.
Linamarin is said to be antineoplastic by its release of HCN during the process of hydrolysis. When HCN is released, the cancer cell is exposed to the lethal cyanide effect released by linamarin. Linamarin is broken down and cyanide is released only in the areas around the cancer cells. This causes gradual cancer cell death. Because normal cells do not have the linamarase gene, they will not be affected5,6.
Inhibition of Raji cell growth is also due to ß-carotene content in cassava leaves. β-carotene has an anticancer mechanism by its carcinogen-modulating metabolism and antioxidant activity, thus modulating the immune system, increasing cell differentiation, stimulating communication gap cell junctions to cells and affecting retinoid-dependent signals29. β-carotene is also directly related to inhibition of cell proliferation, increased apoptosis, induces cell cycle arrest14. In his research, Enger et al.16 stated that β-carotene is protective toward colon adenoma in the early stages of tumor formation. The same thing was determined by Gloria et al.14 who proved that carotenoids were able to increase breast cancer cell apoptosis.
Inhibition of Raji cell growth by linamarin can also be influenced by vitamin C. Cassava leaves contain vitamin C of 103 mg, higher than other green vegetables16. Vitamin C is known to act as an antioxidant in preventing infection, helps the absorption of iron and calcium and is associated with the synthesis of collagen, carnitine, noradrenaline and serotonin in the body30-33. Besides its function, vitamin C also plays an important role in activating genes involved in DNA repair, as well as modulating DNA damage in ROS-affected cells. The results of the Kontek et al.17 study prove that vitamin C has a positive effect on the level of oxidative DNA damage. Vitamin C provides a protective effect for normal tissue to counteract the activity of toxic substances and their metabolites, thus affecting the extent of colon cancer cell inhibition34,35.
Antiproliferative activity of Raji cells: Analysis of cell proliferation inhibition can be done by the doubling time test. Compounds that delay the multiplication times of cells can inhibit genes or proteins that regulate the cell cycle. The doubling time test is done by counting the number of cells treated in a time unit (e.g., 24 h). Each sample is calculated by a hemocytometer and then a curve with cell number versus incubation time is made. Differences in cells’ doubling times can be determined from the slope of the curve or calculated by extrapolation36. Raji cells were previously fasted (starved) for 24 h using RPMI 1640 media containing FBS 0.5%. Reducing this growth signal is necessary because it reduces the speed of cell growth, which causes the cell to be in the same initial start, or G0 phase. Without fasting when treated, the cells remain in different phases which makes it difficult to observe the inhibition properties of linamarin on cell cycle progression37.
From Table 2, it can be seen that the doubling time value of Raji cells with linamarin treatment concentrations of 62.5 μg mL–1 is greater than the doubling time value of Raji cells with linamarin treatments of 32.5 and 15.63 μg mL–1. This is supported by the linamarin curve slope value of 62.5 μg mL–1, which is smaller than the linamarin slope curve of the treatment with 32.5 and 15.63 μg mL–1. This means that linamarin 62.5 μg mL–1 has a better chance of postponing cell doubling time of Raji cells than linamarin 32.5 and 15.63 μg mL–1. It is suspected that the linamarin in cassava leaf extract can inhibit genes or proteins that regulate cell division. It may inhibit signal transduction through inhibition of growth signals or through inhibition of cell cycle progression by inhibiting proto-oncogenes such as CycD, cdk 4/6 and c-myc. Similarly, it may activate suppressor tumors such as caspase 3/8/9, p53, pRb and Bcl2 inactivation5,6.
The data in Table 2 shows that the doubling time value of Raji cells with linamarin treatment concentrations of 62.5 μg mL–1 is twice the doubling time value of Raji cells without treatment (control). This means that linamarin concentration of 62.5 μg mL–1 can cut the doubling time of Raji cells to half that of Raji cells doubling times without treatment (control). The price of doubling time for linamarin treatment is greater than that for control. This indicates that linamarin has the ability to inhibit Raji cell proliferation and possess cytotoxic activity. The higher the linamarin concentration, the longer the doubling time of Raji cells. A linamarin construction of 31.25 μg mL–1 can inhibit cell proliferation better than linamarin 15.63 μg mL–1. This inhibition may occur in signal transduction through inhibition of growth signals or through inhibition of cell cycle progression by inhibiting proto-oncogenes such as CycD, cdk 4/6 and C-Myc. Or, it may be able to activate suppressor tumors such as caspase 3/8/9, p53, pRb and Bcl2 inactivation38,39.
Expression of p53 protein in Raji cells with linamarin treatment: Immunocytochemical analysis is intended to determine the expression of p53 protein in Raji cells. In this study antibodies can be used to detect both wild and mutant type p53 proteins in cancer cells. Positive expression of p53 protein is indicated by brown color in the cell nucleus or cytoplasm; wild or mutant types cannot be distinguished. The results showed that linamarin could increase the expression of p53 protein in Raji cell. Linamarin concentrations of 62.5 μg mL–1 can increase positive p53 protein expression (77.5±3.07%) greater than linamarin 31.25 μg mL–1 (60±1.87%) (Table 1). In Raji control cells or with linamarin treatment from cassava leaf extract, most p53 protein expressions are located in the cell nucleus, although some are located in the cytoplasmic part (Table 3). The control cells also shown have positive p53 protein expression but the amount was less than the treatment with linamarin concentrations of 31.25 and 62.5 μg mL–1 (Fig. 1). This shows that Raji cell death occurred through the mechanism of inhibition of Raji cell proliferation, by activating suppressor gene tumors such as p53. The presence of stress or DNA damage can spur the expression of p53 protein in Raji cells40.
The increase in p53 protein expression in Raji cells after the linamarin treatment proved several possibilities: first, the increase was an increase in wild type p53 expression. P53 protein is encoded by p53 tumor suppressor genes and has an important role in cell regulation and proliferation23. The wild type of p53 protein is expressed very little in normal conditions, but there will be an increase in response to normal cells if there is DNA damage41. Increased expression of wild-type p53 will be activated through the p21 protein to stop DNA replication and cell division when DNA damage occurs. This happens because an increase in p53 protein will stimulate p21 gene transcription. The p21 protein is an inhibitor of CDK and has the ability to inhibit phosphorylation of pRB, thus blocking the release of E2F transcription factors and DNA replication. However, if DNA damage is too severe and cannot be repaired, p53 will induce apoptosis by stimulating Bax transcription, which will then inhibit the activity of the Bcl2 gene42. The Bcl2 gene functions to inhibit the response of apoptosis to various cell types caused by various stimulations related to apoptosis. Thus, p53 plays an important role in preventing the accumulation of cells with DNA abnormalities that can mutate into cancer cells43.
If the p53 expression is the wild type, then DNA damage will cause a rapid rise in p53 protein expression, thus inducing a resting phase of the cell cycle during the G1 phase. Wild-type p53 will cause a cessation of growth in the G1 phase44, thus providing sufficient time for the DNA repair genes such as MLH, MSH2, PMS1, PMS2, Mdm2, BRCA1 and BRCA245. If the DNA damage can be repaired, the cell will continue to divide into the S phase; if this improvement is not possible, then p53 will induce apoptosis46.
The second possibility is that the increase in p53 expression is an accumulation of mutant type p53. P53 mutations will cause the protein to be more stable and have a longer half-life than the wild type. This causes the mutant type of p53 protein to be more easily detected immunocytochemically, although positive expression of p53 is not always associated with its gene mutation47.
P53 mutation is the most common genetic lesion in neoplasms. P53 mutations are associated with increased cellular proliferation and transformation toward malignancy48. They will cause changes in the encoded protein products, so they cannot stimulate the transcription of p21 and Bax42, thus causing the accumulation of cells with DNA damage, which can turn into cancer cells23.
The presence of positive p53 protein expression in the cytoplasm shows that inhibition of Raji cell growth occurs in the G1 phase of the cell cycle. Linamarin from cassava leaves can increase the expression of p53 protein in the cytoplasm compared to the control cells. Linamarin is thought to inhibit cell division in the G1 phase of the cell cycle by increasing the expression of p53 protein in the cytoplasm. According to Groeger et al.49, most of the p53 genes act as 'the guardian of the genome': (1) p53 levels increase rapidly in response to DNA damage, (2) cause cell cycle inhibition during the G1 phase, (3) give cells time to repair DNA damage, (4) if damage cannot be repaired, p53 will induce programmed cell death (apoptosis). Both wild type and mutant proteins migrate in the cell nucleus known as Nuclear Localization Signals (NLS) that are attached to their primary sequences50. According to Burck et al.51 and McManus and Alessi52 p53 wild-type causes growth inhibition in the G1 phase, so that it can be interpreted that in order to enter S phase of the cell, p53 must be inactive.
Overall it can be concluded that linamarin from cassava leaves is toxic to Raji cells and can inhibit Raji cell proliferation through increased expression of p53 protein. The expression of p53 protein cannot be distinguished whether p53 is wild or mutant type but seeing the expression of p53 protein in the cytoplasm shows that inhibition of Raji cell proliferation is through cell cycle progression inhibition that occurs in the G1 phase. This provides an opportunity for genes that control DNA repair to restore DNA function. The limitation of this study is that it only observes the mechanism of Raji cell proliferation via p53 protein induction, so further research is necessary to discern the pathway(s) for proliferation inhibition through apoptosis induction, p21 expression, DNA repair pathways and proliferative inhibition locations in the G1 phase of the cell cycle.
CONCLUSION
Linamarin isolated from cassava leaves (M. esculenta Cranz) has the potential to be developed as an anticancer agent. Linamarin from cassava leaves (M. esculenta Cranz) has cytotoxic activity on Raji cells with IC50 values of 71.865±0.229 μg mL–1, antiproliferation activity on Raji cells with a doubling time value of 40.723 h on linamarin concentration of 62.5 μg mL–1 and can increase the expression of p53 protein in the nuclei and cytoplasm of Raji cells.
SIGNIFICANCE STATEMENT
Findings from this study could contribute to a better understanding of the mechanism of action of linamarin, which is derived from cassava leaves as an anticancer agent. Future efforts should be directed towards determining the specific cell signaling pathways involved in cancer cell toxicity. It also needs in vivo models in experimental animals and the development of an ideal anti-cancer drug formulation.
ACKNOWLEDGMENTS
The authors would like to thank the Dean of Public Health Faculty of University of Diponegoro who has funded this study through APBN DIPA of Public Health Faculty of University of Diponegoro funding No. 106/UN7.5.9/HK/ 2018, dated May 31, 2018.
REFERENCES
- Akinpelu, A.O., L.E.F. Amamgbo, A.O. Olojede and A.S. Oyekale, 2011. Health implications of cassava production and consumption. J. Agric. Social Res., 11: 118-125.
Direct Link - Adenle, A.A., O.C. Aworh, R. Akromah and G. Parayil, 2012. Developing GM super cassava for improved health and food security: Future challenges in Africa. Agric. Food Secur., Vol. 1.
CrossRefDirect Link - Ernesto, M., A.P. Cardoso, D. Nicala, E. Mirione and F. Massaza et al., 2002. Persistent konzo and cyanogen toxicity from cassava in Northern Mozambique. Acta Trop., 82: 357-362.
CrossRefDirect Link - Sayre, R., J.R. Beeching, E.B. Cahoon, C. Egesi and C. Fauquet et al., 2011. The BioCassava plus program: Biofortification of cassava for sub-Saharan Africa. Annu. Rev. Plant Biol., 62: 251-272.
CrossRefDirect Link - Idibie, C.A., H. Davids and S.E. Iyuke, 2007. Cytotoxicity of purified cassava linamarin to a selected cancer cell lines. Bioprocess Biosyst. Eng., 30: 261-269.
CrossRefDirect Link - Yusuf, U.F., F.R. Ahmadun, R. Rosli, S.E. Iyuke, N. Billa, N. Abdullah and N. Umar-Tsafe, 2006. An in vitro inhibition of human malignant cell growth of crude water extract of cassava (Manihot esculenta Crantz) and commercial linamarin. Songklanakarin J. Sci. Technol., 28: 145-155.
Direct Link - Haque, M.R. and J.H. Bradbury, 1999. Preparation of linamarase solution from cassava latex for use in the cassava cyanide kit. Food Chem., 67: 305-309.
CrossRefDirect Link - Girald, W., A. Collin and M. Izquierdo, 2011. Toxicity and delivery methods for the linamarase/linamarin/glucose oxidase system, when used against human glioma tumors implanted in the brain of nude rats. Cancer Lett., 313: 99-107.
CrossRefDirect Link - Dorgan, J.F., A. Sowell, C.A. Swanson, N. Potischman, R. Miller, N. Schussler and H.E. Stephenson Jr., 1998. Relationships of serum carotenoids, retinol, α-tocopherol and selenium with breast cancer risk: Results from a prospective study in Columbia, Missouri (United States). Cancer Causes Control, 9: 89-97.
CrossRefDirect Link - Cortes, M.L., V. Garcia-Escudero, M. Hughes and M. Izquierdo, 2002. Cyanide bystander effect of the linamarase/linamarin killer-suicide gene therapy system. J. Gene Med., 4: 407-414.
CrossRefDirect Link - Rivadeneyra-Dominguez, E., A. Vazquez-Luna, J.F. Rodriguez-Landa and R. Diaz-Sobac, 2013. Neurotoxic effect of linamarin in rats associated with cassava (Manihot esculenta Crantz) consumption. Food Chem. Toxicol., 59: 230-235.
CrossRefDirect Link - Van Duijnhoven, F.J.B., H.B. Bueno-De-Mesquita, P. Ferrari, M. Jenab and H.C. Boshuizen et al., 2009. Fruit, vegetables and colorectal cancer risk: The European prospective investigation into cancer and nutrition. Am. J. Clin. Nutr., 89: 1441-1452.
CrossRefDirect Link - Gloria, N.F., N. Soares, C. Brand, F.L. Oliveira, R. Borojevic and A.J. Teodoro, 2014. Lycopene and beta-carotene induce cell-cycle arrest and apoptosis in human breast cancer cell lines. Anticancer Res., 34: 1377-1386.
Direct Link - Levrero, M., V. de Laurenzi, A. Costanzo, J. Gong, J.Y. Wang and G. Melino, 2000. The p53/p63/p73 family of transcription factors: Overlapping and distinct functions. J. Cell Sci., 113: 1661-1670.
Direct Link - Enger, S.M., M.P. Longnecker, M.J. Chen, J.M. Harper, E.R. Lee, H.D. Frankl and R.W. Haile, 1996. Dietary intake of specific carotenoids and vitamins A, C, and E and prevalence of colorectal adenomas. Cancer Epidemiol. Prev. Biomarkers, 5: 147-153.
Direct Link - Kontek, R., B. Kontek and K. Grzegorczyk, 2013. Vitamin C modulates DNA damage induced by hydrogen peroxide in human colorectal adenocarcinoma cell lines (HT29) estimated by comet assay in vitro. Arch. Med. Sci., 9: 1006-1012.
CrossRefPubMedDirect Link - Mosmann, T., 1983. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods, 65: 55-63.
CrossRefPubMedDirect Link - Wikanta, T., M. Rasyidin, L. Rahayu and A. Pratitis, 2012. [Cytotoxic activity and apoptosis induction of Ulva fasciata Delile ethyl acetate extract against CaSki and MCF-7 cell lines]. J. Pascapanen Bioteknol. Kelautan Perikanan, 7: 87-96, (In Indonesian).
CrossRefDirect Link - Itharat, A and B. Ooraikul, 2007. Research on Thai medicinal plants for cancer treatment. Adv. Med. Plant Res., 37: 287-317.
Direct Link - He, Y., Q. Zhu, M. Chen, Q. Huang and W. Wang et al., 2016. The changing 50% inhibitory concentration (IC50) of cisplatin: A pilot study on the artifacts of the MTT assay and the precise measurement of density-dependent chemoresistance in ovarian cancer. Oncotarget, 7: 70803-70821.
CrossRefDirect Link - Jorgensen, K., A.V. Morant, M. Morant, N.B. Jensen and C.E. Olsen et al., 2011. Biosynthesis of the cyanogenic glucosides linamarin and lotaustralin in cassava: Isolation, biochemical characterization and expression pattern of cyp71e7, the oxime-metabolizing cytochrome P450 enzyme. Plant Physiol., 155: 282-292.
CrossRefPubMedDirect Link - Lane, D.P., C.F. Cheok and S. Lain, 2010. P53-based cancer therapy. Cold Spring Harbor Perspect. Biol., Vol. 2, No. 9.
CrossRefDirect Link - Afsar, T., J.H. Trembley, C.E. Salomon, S. Razak, M.R. Khan and K. Ahmed, 2016. Growth inhibition and apoptosis in cancer cells induced by polyphenolic compounds of Acacia hydaspica: Involvement of multiple signal transduction pathways. Scient. Rep., Vol. 6.
CrossRefDirect Link - Lehmann, B.D., J.A. Bauer, X. Chen, M.E. Sanders, A.B. Chakravarthy, Y. Shyr and J.A. Pietenpol, 2011. Identification of human triple-negative breast cancer subtypes and preclinical models for selection of targeted therapies. J. Clin. Invest., 121: 2750-2767.
CrossRefDirect Link - Khan, N., F. Afaq, M. Saleem, N. Ahmad and H. Mukhtar, 2006. Targeting multiple signaling pathways by green tea polyphenol (−)-epigallocatechin-3-gallate. Cancer Res., 66: 2500-2505.
CrossRefDirect Link - Ghate, N.B., B. Hazra, R. Sarkar and N. Mandal, 2014. Heartwood extract of Acacia catechu induces apoptosis in human breast carcinoma by altering bax/bcl-2 ratio. Pharmacogn. Mag., 10: 27-30.
CrossRefDirect Link - Catz, S.D. and J.L. Johnson, 2001. Transcriptional regulation of bcl-2 by nuclear factor κB and its significance in prostate cancer. Oncogene, 20: 7342-7351.
CrossRefDirect Link - Bolhassani, A., A. Khavari, S.Z. Bathaie, 2014. Saffron and natural carotenoids: Biochemical activities and anti-tumor effects. Biochim. Biophys. Acta (BBA)-Rev. Cancer, 1845: 20-30.
CrossRefPubMedDirect Link - Duarte, T.L. and J. Lunec, 2005. Review: When is an antioxidant not an antioxidant? A review of novel actions and reactions of vitamin C. Free Radical Res., 39: 671-686.
CrossRefPubMedDirect Link - Verma, R.S., A. Mehta and N. Srivastava, 2007. In vivo chlorpyrifos induced oxidative stress: Attenuation by antioxidant vitamins. Pesticide Biochem. Phys., 88: 191-196.
CrossRefDirect Link - Szarka, A., B. Tomasskovics and G. Banhegyi, 2012. The Ascorbate-glutathione-α-tocopherol triad in abiotic stress response. Int. J. Mol. Sci., 13: 4458-4483.
CrossRefDirect Link - Bindhumol, V., K.C. Chitra and P.P. Mathur, 2003. Bisphenol A induces reactive oxygen species generation in the liver of male rats. Toxicology, 188: 117-124.
CrossRefDirect Link - Winkler, B.S., S.M. Orselli and T.S. Rex, 1994. The redox couple between glutathione and ascorbic acid: A chemical and physiological perspective. Free Radic. Biol. Med., 17: 333-349.
CrossRefDirect Link - Griffiths, H.R. and J. Lunec, 2001. Ascorbic acid in the 21st century-more than a simple antioxidant. Environ. Toxicol. Pharmacol., 10: 173-182.
CrossRefDirect Link - Finlay, C.A., P.W. Hinds and A.J. Levine, 1989. The p53 proto-oncogene can act as a suppressor of transformation. Cell, 57: 1083-1093.
CrossRefPubMedDirect Link - Oraiopoulou, M.E., E. Tzamali, G. Tzedakis, A. Vakis, J. Papamatheakis and V. Sakkalis, 2017. In vitro/in silico study on the role of doubling time heterogeneity among primary glioblastoma cell lines. BioMed Res. Int., Vol. 2017.
CrossRefDirect Link - Atuegwu, N.C., L.R. Arlinghaus, X. Li, A.B. Chakravarthy, V.G. Abramson, M.E. Sanders and T.E. Yankeelov, 2013. Parameterizing the logistic model of tumor growth by DW-MRI and DCE-MRI data to predict treatment response and changes in breast cancer cellularity during neoadjuvant chemotherapy. Transl. Oncol., 6: 256-264.
CrossRefPubMedDirect Link - Bertuzzi, A., A. Gandolfi, C. Sinisgalli, G. Starace and P. Ubezio, 1997. Cell loss and the concept of potential doubling time. Cytometry Part A: J. Quant. Cell Sci., 29: 34-40.
CrossRefDirect Link - Lowe, S.W., 1999. Activation of p53 by oncogenes. Endocr. Relat. Cancer, 6: 45-48.
CrossRefDirect Link - Rivlin, N., R. Brosh, M. Oren and V. Rotter, 2011. Mutations in the p53 tumor suppressor gene: Important milestones at the various steps of tumorigenesis. Genes Cancer, 2: 466-474.
CrossRefDirect Link - Sugerman, P.B. and N.W. Savage, 1999. Current concepts in oral cancer. Aust. Dent. J., 44: 147-156.
CrossRefDirect Link - Petitjean, A., E. Mathe, S. Kato, C. Ishioka, S.V. Tavtigian, P. Hainaut and M. Olivier, 2007. Impact of mutant p53 functional properties on TP53 mutation patterns and tumor phenotype: Lessons from recent developments in the IARC TP53 database. Hum. Mutat., 28: 622-629.
CrossRefDirect Link - Hainaut, P. and M. Hollstein, 1999. p53 and human cancer: The first ten thousand mutations. Adv. Cancer Res., 77: 81-137.
CrossRefDirect Link - Schlomm, T., L. Iwers, P. Kirstein, B. Jessen and J. Kollermann et al., 2008. Clinical significance of p53 alterations in surgically treated prostate cancers. Mod. Pathol., 21: 1371-1378.
CrossRefDirect Link - Nozaki, M., M. Tada, H. Kobayashi, C.L. Zhang and Y. Sawamura et al., 1999. Roles of the functional loss of p53 and other genes in astrocytoma tumorigenesis and progression. Neuro-oncology, 1: 124-137.
CrossRefDirect Link - Oren, M. and V. Rotter, 2010. Mutant p53 gain-of-function in cancer. Cold Spring Harbor Perspect. Biol., Vol. 2.
CrossRefDirect Link - Groeger, A.M., V. Esposito, A. de Luca, R. Cassandro and G. Tonini et al., 2004. Prognostic value of immunohistochemical expression of p53, bax, Bcl-2 and Bcl-xL in resected non-small-cell lung cancers. Histopathology, 44: 54-63.
CrossRefDirect Link - Shaulsky, G., N. Goldfinger, M.S. Tosky, A.J. Levine and V. Rotter, 1991. Nuclear localization is essential for the activity of p53 protein. Oncogene, 6: 2055-2065.
PubMed - McManus, E.J. and D.R. Alessi, 2004. Cancer, oncogenes and signal transduction. Genome Biol., Vol. 5.
CrossRefDirect Link