
R. Sunkara
Department of Food and Animal Sciences, Nutrition and Carcinogenesis Laboratory, Normal, Alabama
M. Verghese
Department of Food and Animal Sciences, Nutrition and Carcinogenesis Laboratory, Normal, Alabama
L. Shackelford
Department of Food and Animal Sciences, Nutrition and Carcinogenesis Laboratory, Normal, Alabama
L.T. Walker
Department of Food and Animal Sciences, Nutrition and Carcinogenesis Laboratory, Normal, Alabama
E. Cebert
Department of Natural Resources and Environmental Sciences, Alabama A and M University, Normal, Alabama
International Journal of Cancer Research
Year: 2009 | Volume: 5 | Issue: 4 | Page No.: 167-175







ABSTRACT
The objective of this study was to determine the effects of dietary administration of sorrel calyx and sorrel juice on the induction and development of azoxymethane (AOM) induced Aberrant Crypt Foci (ACF) in Fisher 344 rats. Thirty male F344 rats were randomly divided into 5 groups and fed AIN 93-G diet as control, AIN 93-G+sorrel meal (5 and 10%) or juice (2.5 and 5%). Rats were given subcutaneous injections of AOM (16 mg kg-1 b.wt.) once a week for 2 weeks and were killed after administering sorrel for 13 weeks. The effect of sorrel on detoxification (phase I and II) and antioxidant enzymes (catalase (CAT) and superoxide dismutase (SOD)) was also determined. Treatment with dietary sorrel meal and juice significantly (p<0.05) decreased total number of ACF and the total number of aberrant crypts compared to the control fed rats. Feeding sorrel meal at 10% level significantly (p<0.05) reduced the percentage of ACF by 70% compared to the control. Administration of sorrel to rats significantly (p<0.05) increased hepatic Glutathione-S-Transferase (GST), CAT and SOD activities by 3.3-6.5 fold compared to the control fed rats. However, no effect on CYP2E1 activity was observed among the treatment groups. Present results demonstrate the chemopreventive potential of sorrel either as meal or juice on AOM induced ACF as well as its ability to modulate detoxification and antioxidant enzymes.
PDF Abstract XML References Citation
How to cite this article
R. Sunkara, M. Verghese, L. Shackelford, L.T. Walker and E. Cebert, 2009. Preventive Potential of Sorrel (Hibiscus sabdariffa) Calyx on the Formation of Azoxymethane Induced Aberrant Crypt Foci in Colon of Rats. International Journal of Cancer Research, 5: 167-175.
DOI: 10.3923/ijcr.2009.167.175
URL: https://scialert.net/abstract/?doi=ijcr.2009.167.175
DOI: 10.3923/ijcr.2009.167.175
URL: https://scialert.net/abstract/?doi=ijcr.2009.167.175
INTRODUCTION
Colon cancer may occur through genetic mutations by exposure to carcinogens and by environmental factors. Epidemiological studies revealed the highest incidence of colon cancer in populations with diets containing high fat, low fiber, high calorie and low fruit and vegetable intakes (Bingham, 1999; Slattery et al., 1998). An effective preventive measure which includes diet modification is one of the approaches to reduce colon cancer morbidity and mortality. Current evidence suggests that bioactive food components in fruits and vegetables may affect the development of colon cancer (Ross et al., 2006). Ubiquitous in nature, bioactive compounds are known to act as antioxidants (Geber and Bowerman, 2001), activate liver detoxification enzymes, destroy harmful ROS, block carcinogens, suppress malignant cells and interfere with several processes known to cause cancer (Pennington, 2002).
Sorrel, Hibiscus sabdariffa L., belongs to Malvaceae family and the calyx of this plant is commonly used to prepare cold drinks, hot tea and jellies. It is commonly known as Sorrel, rossele, or karkade. In some parts of the world, the leaves are used as green leafy vegetables. The calyx contains bioactive compounds such as polyphenols, flavonoids, anthocyanins, volatile acids and protocatechuic acid along with several minerals and vitamins. Anthocyanins are the most abundant bioactive components in sorrel drink. The water or solvent extracts of sorrel calyx have shown hepatoprotective, cardioprotective (Jonadet et al., 1990; Chen et al., 2003), antihypertensive (Herrera-Arellano et al., 2004; Odigie et al., 2003), antidiabetic, antidiuretic (Mojiminiyi et al., 2000) and in vitro anticancer properties. Hibiscus anthocyanins (HAs) possess antioxidant bioactivity both in vivo and in vitro (Tseng et al., 1997; Wang et al., 2000).
The azoxymethane (AOM)-Fisher 344 (F344) model is extensively being used in colon cancer prevention studies and has provided significant answers on the development of the disease. Perhaps, the most important aspect of this model is the fact that it is reliable and many of the clinical, histological and molecular features resemble sporadic human colon cancer (Takahashi and Wakabayashi, 2004). Aberrant Crypt Foci (ACF) are the earliest identifiable mucosal abnormalities in carcinogen-treated rodents and can be scored at 14 wks after two AOM injections in rats (Bird, 1995).
Biotransformation of carcinogens occurs by the action of phase I and II enzymes within the cell. At the initiation stage of carcinogenesis, carcinogens are converted into reactive electrophiles by the action of Phase I CYP450 enzymes, leading to mutations and genetic lesions in DNA. Reactive electrophiles conjugate with different water soluble substrates by phase II detoxification enzymes (GST, Sulfotranferases etc.) and are eliminated from the body. In general, the bioactivation reaction often generates Reactive Oxygen Species (ROS) directly as a spin-off product, since it uses oxygen; hence, dietary antioxidants such those from fruits and vegetables can help protect tissue from damage that may occur by this reaction.
Therefore, focus has been given for the potential use of whole foods and synergy of phytochemicals to reduce cancer. In this study we used sorrel calyx in the form of meal and juice to determine its chemopreventive potential against AOM induced ACF in Fisher 344 rats.
MATERIALS AND METHODS
Preparation of Sorrel Meal and Juice
Dry Sorrel calyxes were obtained (Alabama A and M University) and ground to a fine powder (particle size of 0.5 mm) using a Wiley mill (Swedesboro, NJ) and mixed into the diet. Modifications were made to cornstarch, sucrose and fiber in order to make the diets isocaloric with AIN 93G. One thousand milliliter of distilled water was heated to a boiling temperature. One hundred grams of dried sorrel was added to the boiling water and allowed to boil for an additional 10 min. The sorrel decoction was allowed to cool to room temperature and was filtered. Desired 2.5 and 5% sorrel juice was prepared daily from the concentrated stock.
Animal Care and Experimental Design
Male Fisher 344 weanling rats (Harlan, IN) rats were housed following standard protocols. All rats were subjected to a 1-week adaptation period and randomly assigned to 5 groups (n = 6) and assigned to the following diets: AIN-93G (Reeves et al., 1993) (control), AIN-93G diet containing 5 and 10% ground sorrel calyx with modifications and AIN-93G with 2.5 and 5% sorrel juice in place of water. Rats were given ad libitum access to the diets, weekly body weight and daily feed and daily fluid intakes were recorded. The diets were prepared weekly and stored at 4°C. Ingredients for preparing AIN93G diets were obtained from M.P. Biomedicals (Costa Mesa, CA). The Institutional Animal Care and Use Committee of Alabama A and M University approved all protocols involving rats.
Carcinogen Injection and Tissue Sample Collection
To induce ACF, two subcutaneous injections of Azoxymethane (AOM), (NCI Repository and Kansas City, MO) were administered to the rats’ at 16 mg kg-1 b.wt. during the 7th and 8th week of age. After 13 week, feed was withheld overnight and the rats were killed by CO2 asphyxiation. The cecum from each rat was excised, weighed and split open and the pH of the cecal contents was recorded. Colon was removed for ACF enumeration and liver was collected, rinsed in potassium phosphate buffer (pH 7.0, 0.1 M), flash frozen in liquid nitrogen and immediately frozen at -80°C until enzyme assays were conducted.
Quantification of ACF
Colons of rats from each group were removed and flushed with PBS (0.1 M, pH 7.2). These were cut open then fixed in buffered formalin. Each colon was cut into 2 equal sections, distal and proximal and stained with methylene blue. ACF and crypts per focus were enumerated as described by Bird (1987). ACF were classified into small (≤3 crypts) and large (≥4 crypts) based on number of ACF/focus.
Preparation of Homogenates and Microsomal Fractions
Hepatic tissue was homogenized in ice-cold homogenizing buffer (20 mM Hepes, pH 7.4 containing 100 mM KCl, 1 mM MgCl2, 1 mM EGTA, 210 mM mannitol and 70 mM sucrose) to prepare 10% (w/v) homogenate. An aliquot was saved from the homogenate after centrifuging at 10,000 g for 30 min to quantify the antioxidative enzymes: SOD and catalase activities. The remaining supernatant was further centrifuged at 100,000 g for 60 min at 4°C. Microsomal pellets were mixed with equal volumes of homogenization buffer and used to determine CYP2E1 and GST activities.
Hepatic Enzyme Activities
Hydroxylation of 4-nitrophenol to 4-nitrocatechol was determined by the method described by Koop et al. (1997). Briefly, 4-Nitrophenol (0.2 mM) was dissolved in 50 mM Tris-HCl, 25 mM MgCl2, pH 7.4. An aliquot of 930 μL of this solution and 50 μL of liver microsomal sample were incubated at 37°C. Reactions were started by adding 50 mM NADPH and stopped by the addition of 0.6 N perchloric acid. The formation of 4-Nitrocatechol formation was spectrophotometrically determined with the addition of 10 N NaOH at 600 nm. Hepatic GST activity was assayed according to the spectrophotometric methods using (Habig et al., 1974) 1, chloro 2, 4-dinitrobenzene (CDNB) (Sigma chemicals, St. Louis, MO) as substrate. Change in absorbance was recorded every minute for 5 min in a Cary 1/3 UV/VIS dual beam spectrophotometer (Palo Alto, CA). The CAT and SOD activities were analyzed using kits from Cayman Chemical (Ann Arbor, MI) according to the manufacturer’s instructions.
Statistical Analysis
Data are expressed as Means±SEM and were analyzed using the SAS statistical program (SAS 9.1) by Analysis of Variance (ANOVA) and means were separated using Tukey’s studentized range test. Differences were determined for statistical significance using two-way ANOVA. Differences were considered significant at p<0.05.
RESULTS
General Observations
Rats fed the treatment diets had similar weight gain, feed intake, cecal weight and pH (Table 1). Weight gain (13 weeks) in rats fed sorrel was comparable to the control fed rats, indicating that the dose or treatments used did not cause any toxicity or side effects. The highest feed consumption was seen in rats fed SM 10% and lowest intake was seen in control fed rats. Cecal pH was near neutrality in all the groups including the control; although, the lowest cecal pH and highest cecal weight were observed in rats fed SM (Table 1).
Incidence of Aberrant Crypt Foci (ACF) in Colon of Fisher 344 Male Rats
Administration of sorrel significantly (p<0.05) reduced the number of ACF compared to the control fed rats (Fig. 1). In all the groups, the incidence of ACF in the distal colon was higher than in the proximal section, which has also been seen in humans. While the number of ACF in the proximal colon was comparable in all the treatment groups, it was however significantly (p<0.05) lower (50%) compared to the control fed rats. Rats fed sorrel showed a significantly (p<0.05) lower number of distal ACF compared with rats fed the control diet, with the highest reduction seen in the groups fed SM (SM 10% (62.35%) and SM 5% (51.23%)). A similar trend was also observed in the total number of ACF.
| Table 1: | Effect of sorrel calyx on weight gain, feed intake, cecal weight and cecal pH on AOM-induced Fisher 344 male rats |
 | |
| SJ: Sorrel juice; SM: Sorrel meal. Values are as Means±SEM, n = 6 | |
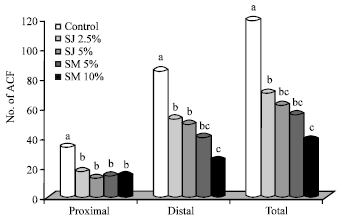 | |
| Fig. 1: | Effect of Sorrel calyx on AOM-induced ACF in the colon of male F344 rats. SJ: Sorrel juice, SM: Sorrel meal. Values are Means±SD n = 6. Bars not sharing a common superscript are significantly different (p<0.05) with Tukey’s studentized range test |
We observed that compared to the control fed rats, supplementation of sorrel meal in the diet significantly (p<0.05) reduced the incidence of ACF by 70.23% (SM 10%) and 50.23% (SM 5%) and as juice by 49.32% (SJ 5%) and 32.45% (SJ 2.5%).
Crypt Multiplicity and Total Crypts
Total crypts in rats fed the control diet (330.50±18.87) was significantly (p<0.05) higher compared to rats fed the treatment diets (Table 2). Present results showed that among the treatment groups, total crypts were similar in the rats fed SJ (2.5 and 5%) and SM 5% but significantly lower in the SM10% fed rats. Compared to the control, the highest reduction in total crypts was observed in rats fed SM 10% (72.54%) and the lowest reduction was observed in the rats fed SJ 2.5% (49%). Increased dosage of SJ and SM resulted in decreased incidence of total crypt. Dietary administration of sorrel juice and meal significantly (p<0.05) reduced the number of small (ACF with crypts ≤3) and large crypts (ACF with crypts ≥4) compared to the control fed rats. Among the treatment groups, ACF with small crypts were significantly (p<0.05) lower in rats fed 10% SM (28.75±3.88), however, we did not observe any significant differences among the treatment groups with regard to ACF with large crypts.
Activity of Phase I and Phase II Enzymes
Feeding sorrel meal and juice did not have any effect in modulating hepatic phase 1 CYP2E1 activity. CYP2E1 activity was not significantly different in all the groups including the control. However, the administration of treatment diets resulted in a significant increase in GST (Table 3) activity compared to the control fed rats. Administration of sorrel meal at (10%) and SJ (5%) increased GST activity (mmol/min/mg) by 6.5 and 5.28 fold, respectively, compared to the control (10.35±1.87). Rats fed SJ (2.5%) had the lowest induction of GST activity among all the treatment groups, however had significantly (p<0.05) higher induction (3.32 fold) compared to the control.
| Table 2: | Crypt multiplicity and number of crypts in colon of rats |
 | |
| SJ: Sorrel juice, SM: Sorrel meal. Values are Means±SEM; n = 6. Values in a column not sharing a common superscript are significantly different (p<0.05) with Tukey’s studentized range test | |
| Table 3: | CYP2E1 and Glutathione S-transferase activity in liver of rats |
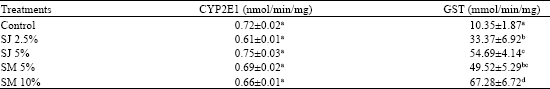 | |
| SJ: Sorrel Juice; SM: Sorrel meal. Values are Mean±SEM; n = 6. Values in a column not sharing a common superscript are significantly different (p<0.05) with Tukey’s studentized range | |
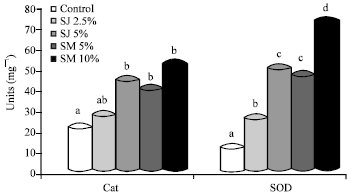 | |
| Fig. 2: | Activity of hepatic antioxidative enzyme. SJ: Sorrel juice, SM: Sorrel meal, CAT catalase, SOD: Superoxide dismutase values are Means±SEM, n = 6. Bars not sharing a common superscript are significantly different (p<0.05) with Tukey’s studentized range test |
Activity of Antioxidative Enzymes (SOD and CAT)
CAT and SOD were significantly (p<0.05) higher in rats fed treatment diets compared with the control rats (Fig. 2). Except in the group fed SJ (2.5%), CAT activity (U mg-1) in rats fed SM (10%), SM (5%) and SJ (5%) were significantly (p<0.05) higher compared to the group fed control diet (20.65). With SOD activity, the highest induction, compared to the control fed rats, was observed in rats fed SM 10% (6.77 fold), followed by SJ 5% (4.61 fold), SM (5%) (4.33 fold) and SJ (2.5%) (2.35 fold). Among the treatment groups, administration of sorrel at 5% as juice and meal did not have any significant effect in induction of SOD activity. However, both these groups had significantly (p<0.05) higher activity compared to the control.
DISCUSSION
The present study was conducted to evaluate the potential of sorrel in reducing the incidence of ACF in Fisher 344 male rats. The dose levels in this study were selected to determine the upper limits of efficacy of sorrel calyx as a chemopreventive agent as these doses correspond to 1.5-2 servings of fruits and vegetables for human consumption. Present results indicate that administration of sorrel significantly (p<0.05) reduced the incidence of ACF by 32-70% compared to the control fed rats.
Sorrel calyx is a composite of several compounds including anthocyanins, cyanidin-3-rutinoside, protocatechuic acid and quercetin. The additive and synergistic effect of these bioactive compounds present in sorrel might have contributed to the reduction in colonic ACF. One of the mechanisms associated with these phenolic compounds are their in-vivo antioxidant properties as well as their ability to influence signal transduction. The generation of Reactive Oxygen Species (ROS) is by far the most potent threat faced by any living organism (Chandra et al., 2000; MacCarthy and Shah, 2003). As evidenced, accumulation of ROS such as superoxide anion, hydrogen peroxide, singlet oxygen, hydroxyl radical and peroxyl radical, may lead to damage to key biological macromolecules, including DNA and lipids (Paravicini and Touyz, 2008). Oxidative damage to DNA is one of the contributory factors in tumor initiation. Phenolic compounds that possess antioxidant or reactive oxygen scavenging activity may prove beneficial as chemoprevention agents.
Chewonarin et al. (1999) remarked that the reduced incidence of ACF in rats administered a gavage of sorrel may be due to the inhibition of DNA methylation and cytochrome P450 1A2, however, the exact mechanisms are yet to be determined. In our study we noted that rats fed SM had significant (p<0.05) reduction in ACF compared to the rats fed SJ. Aside from its phenolic content, sorrel calyx is a reliable source of dietary fiber. Fiber present in the meal might enhance the release of bioactive components into the colon of rats by the action of colonic microflora. Fiber can also act as blocking agent and decrease the interaction of the carcinogen with the colonic mucosa.
Crypt multiplicity is an important indicator of tumor incidence. Large ACF (crypts≥4) are likely to develop into tumors over time by proliferation (Zhang et al., 1992) and small ACF (crypts≤3) might get dissolved and disappear. Rats fed SM and SJ had significantly (p<0.05) lower number of large and small ACF compared to the rats fed the control diet, thus implying that polyphenolic compounds present in sorrel might have led to the induction of apoptosis thereby decreasing cell proliferation.
AOM is metabolized by CYP2E1 enzyme in the liver to methylazoxymethanol which is responsible for the formation of DNA adducts. Inhibition of CYP2E1 by phytochemicals can halt the process of cancer by preventing the occurrence of genetic lesions. In our study we showed that administration of sorrel had no effect in modulating CYP2E1 enzyme.
Phase II detoxification enzymes such as GST and QR make the carcinogen more water soluble and readily excreted in the urine or feces and bile. Our results are comparable to enhancement of GST and QR in liver and colonic mucosa of male F344 rats given a dose 100 mg kg-1 of ferulic acid (Kawabata et al., 2000). Silymarin, a flavonoid did not influence liver P 450 2E1 in rats (Miguez et al., 1994). Diallyl sulphide, a volatile compound in garlic inhibited the activity of CYP 2E1 in rats (Brady et al., 1991). The GST and QR activities in the liver and colon of rats gavaged with 40, 200 and 400 mg kg-1 b.wt. of silymarin were significantly higher than the control rats induced with AOM (Kohno et al., 2002). No changes in CYP 2E1 activity and enhanced activity of GST suggest an important mechanism in reducing the formation of ACF in sorrel fed rats.
Polyphenols can affect the process of carcinogenesis by counteracting oxidative stress through the induction of antioxidative enzymes that are capable of neutralizing free radicals. Catalase and SOD activities were elevated when rats were fed SJ and SM.
The exact mechanisms of action by sorrel calyx for reducing the number of ACF and crypt multiplicity by AOM are not fully known. However, there are many possible mechanisms of action by the bioactive components present in sorrel. Colon cancer occurs in a step wise manner by the transformation of normal epithelial cells into preneoplastic, neoplastic and metastatic states (Fearon and Vogelstein, 1990). Phytochemicals may act at every stage to halt the process of carcinogenesis. Some of the components such as fiber and polyphenols may act as blocking agents and hence, affect the initiation stage of carcinogenesis. The induction of a crucial phase II detoxification enzyme, GST and antioxidative enzymes suggests their role in reducing the oxidative stress caused by AOM and other xenobiotics.
CONCLUSION
The results of this study showed that administration of sorrel as meal (5 and 10%) and juice (2.5 and 5%) significantly reduced ACF formation induced by AOM. Different mechanisms of action in reducing the incidence of ACF suggest the potential combinational effect of phytochemicals and importance of a food based approach. Further mechanistic and long term tumor model studies should be conducted to confirm the chemopreventive potential observed in the short tern feeding of sorrel calyx.
ACKNOWLEDGMENT
The funding for this research was provided by The Alabama Agricultural Research Experimental Station, Alabama A and M University, Normal, AL 35762.
REFERENCES
- Bingham, S.A., 1999. High-meat diets and cancer risk. Proc. Nutr. Soc., 58: 243-248.
CrossRefPubMedDirect Link - Bird, R.P., 1995. Role of aberrant crypt foci in understanding the pathogenesis of colon cancer. Cancer Lett., 93: 55-71.
CrossRefPubMedDirect Link - Bird, R.P., 1987. Observation and quantification of aberrant crypts in the murine colon treated with a colon carcinogen: Preliminary findings. Cancer Lett., 37: 147-151.
PubMedDirect Link - Brady, J.F., M.H. Wang, J.Y. Hong, F. Xiao and Y. Li et al., 1991. Modulation of rat hepatic microsomal monooxygenase enzymes and cytotoxicity by diallyl sulfide. Toxicol. Applied Pharmacol., 108: 342-354.
CrossRefDirect Link - Chandra, J., A. Samali and S. Orrenius, 2000. Triggering and modulation of apoptosis by oxidative stress. Free Radical Biol. Med., 29: 323-333.
CrossRefPubMedDirect Link - Chen, C.C., J.D. Hsu, S.F. Wang, H.C. Chiang and M.Y. Yang et al., 2003. Hibiscus sabdariffa extract inhibits the development of atherosclerosis in cholesterol-fed rabbits. J. Agric. Food Chem., 51: 5472-5477.
CrossRefDirect Link - Chewonarin, T., T. Kinouchi, K. Kataoka, H. Arimochi, T. Kuwahara, U. Vinitketkumnuen and Y. Ohnishi, 1999. Effects of roselle (Hibiscus sabdariffa Linn), a Thai medicinal plant, on the mutagenicity of various known mutagens in Salmonella typhimurium and on formation of aberrant crypt foci induced by the colon carcinogens azoxymethane and 2-amino-1-methyl-6-phenylimidazo[4, 5-b]pyridine in F 344 rats. Food Chem. Toxicol., 37: 571-601.
- Fearon, E.R. and B. Vogelstein, 1990. A genetic model for colorectal tumorigenesis. Cell, 61: 759-767.
PubMedDirect Link - Habig, W.H., M.J. Pabst and W.B. Jakoby, 1974. Glutathione S-transferases: The first enzymatic step in mercapturic acid formation. J. Biol. Chem., 249: 7130-7139.
CrossRefPubMedDirect Link - Geber, D. and S. Bowerman, 2001. Applying science to changing dietary patterns. J. Nutr., 131: 3078S-3081S.
PubMed - Herrera-Arellano, A., S. Flores-Romero, M.A. Chavez-Soto and J. Tortorie-Ilo, 2004. Effectiveness and tolerability of a standardized extract from Hibiscus sabdariffa in patients with mild to moderate hypertension: A controlled and randomized clinical trial. Phytomedicine, 5: 375-382.
CrossRefPubMedDirect Link - Jonadet, M., J. Bastide, P. Bastide, B. Boyer, A.P. Carnat and J.L. Lamaison, 1990. In vitro enzyme inhibitory and in vivo cardioprotective activities of hibiscus (Hibiscus sabdariffa L.). J. Pharm. Belg., 45: 120-124.
PubMed - Kawabata, K., T. Yamamoto, A. Hara, M. Shimizu and Y. Yamada et al., 2000. Modifying effects of ferulic acid on azoxymethane-induced colon carcinogenesis in F344 rats. Cancer Lett., 157: 15-21.
PubMed - Kohno, H., T. Tanaka, K. Kawabata, Y. Hirose, S. Sugie, H. Tsuda and H. Mori, 2002. Silymarin, a naturally occurring polyphenolic antioxidant flavonoid, inhibits azoxymethane-induced colon carcinogenesis in male F344 rats. Int. J. Cancer, 101: 461-468.
PubMed - Koop, D.R., B. Klopfenstein, Y. Iimuro and R.G. Thurman, 1997. Gadolinium chloride blocks alcohol-dependent liver toxicity in rats treated chronically with intragastric alcohol despite the induction of CYP2E1. Mol. Pharmacol., 51: 944-950.
PubMed - MacCarthy, P.A. and A.M. Shah, 2003. Oxidative stress and heart failure. Coron. Artery Dis., 14: 109-113.
PubMed - Miguez, M.P., I. Anundi, L.A. Sainz-Pardo and K.O. Lindros, 1994. Hepatoprotective mechanism of silymarin: No evidence for involvement of cytochrome P450 2E1. Chem. Biol. Interact., 91: 51-63.
PubMed - Odigie, I.P., R.R. Ettarh and S.A. Adigun, 2003. Chronic administration of aqueous extract of Hibiscus sabdariffa attenuates hypertension and reverses cardiac hypertrophy in 2K-1 C hypertensive rats. J. Ethnopharmacol., 86: 181-185.
PubMed - Pennington, A.T., 2002. Food composition databases for bioactive food components. J. Food Compost Anal., 15: 419-434.
CrossRef - Reeves, P.G., F.H. Nielsen and G.C. Fahey Jr., 1993. AIN-93 purified diets for laboratory rodents: Final report of the American Institute of Nutrition ad hoc writing committee on the reformulation of the AIN-76A rodent diet. J. Nutr., 123: 1939-1951.
CrossRefPubMedDirect Link - Ross, S.A., J.W. Finley and J.A. Milner, 2006. Allyl sulfur compounds from garlic modulate aberrant crypt formation. J. Nutr., 136: 852S-854S.
PubMed - Slattery, M.L., K.M. Boucher and B.J. Caan, 1998. Eating patterns and risk of colon cancer. Am. J. Epidemiol., 148: 4-16.
PubMed - Takahashi, M. and K. Wakabayashi, 2004. Gene mutations and altered gene expression in azoxymethane-induced colon carcinogenesis in rodents. Cancer Sci., 95: 475-480.
PubMed - Zhang, X.M., D. Stamp, S. Minkin, A. Medline, D.E. Corpet, W.R. Bruce and M.C. Archer, 1992. Promotion of aberrant crypt foci and cancer in rat colon by thermolyzed protein. J. Natl. Cancer Inst., 84: 1026-1030.
PubMed - Tseng, T.H., E.S. Kao, C.Y. Chu, F.P. Chou, H.M.L. Wu and C.J. Wang, 1997. Protective effects of dried flower extracts of Hibiscus sabdariffa L. against oxidative stress in rat primary hepatocytes. Food Chem. Toxicol., 35: 1159-1164.
CrossRef